
Bacteriophage-Resistant Salmonella rissen: An In Vitro Mitigated Inflammatory Response

 , , , and
, , , and 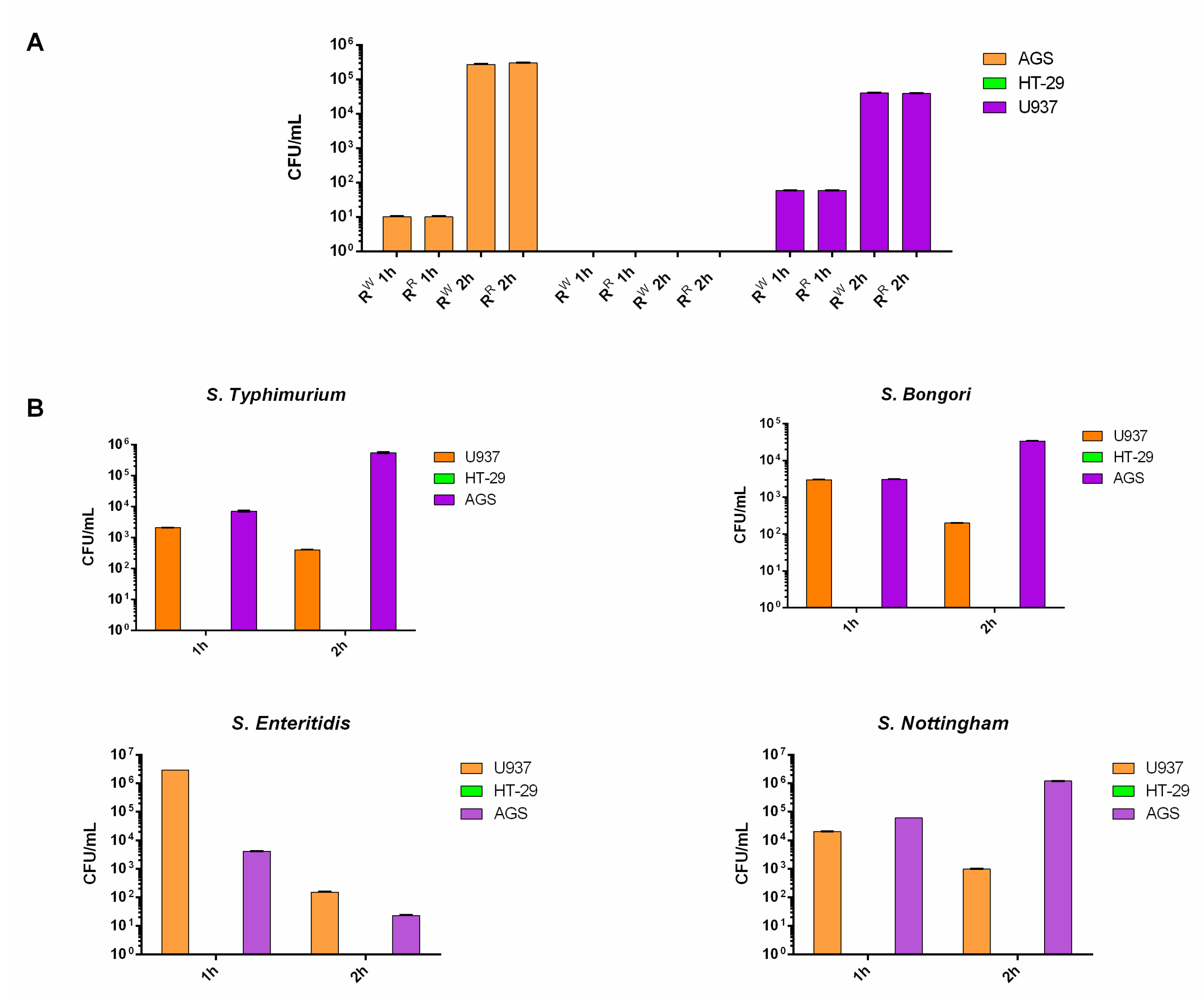{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Cell Lines and Culture Conditions
2.3. Salmonella Invasion Assay
2.4. Expression Levels of Virulence Genes
2.5. Infection on AGS Cell Line
2.6. RNA Extraction and RT-qPCR
2.7. Cytokine Determination by Bio-Plex Assay
2.8. Western Blotting Analysis
2.9. Statistical Analysis
3. Results
3.1. RW and RR Strains Display the Same Antigenic and Antibiotic Resistance Profiles
3.2. The RW and RR Strains Both Exhibit the Same Capacity to Colonize Host Cells
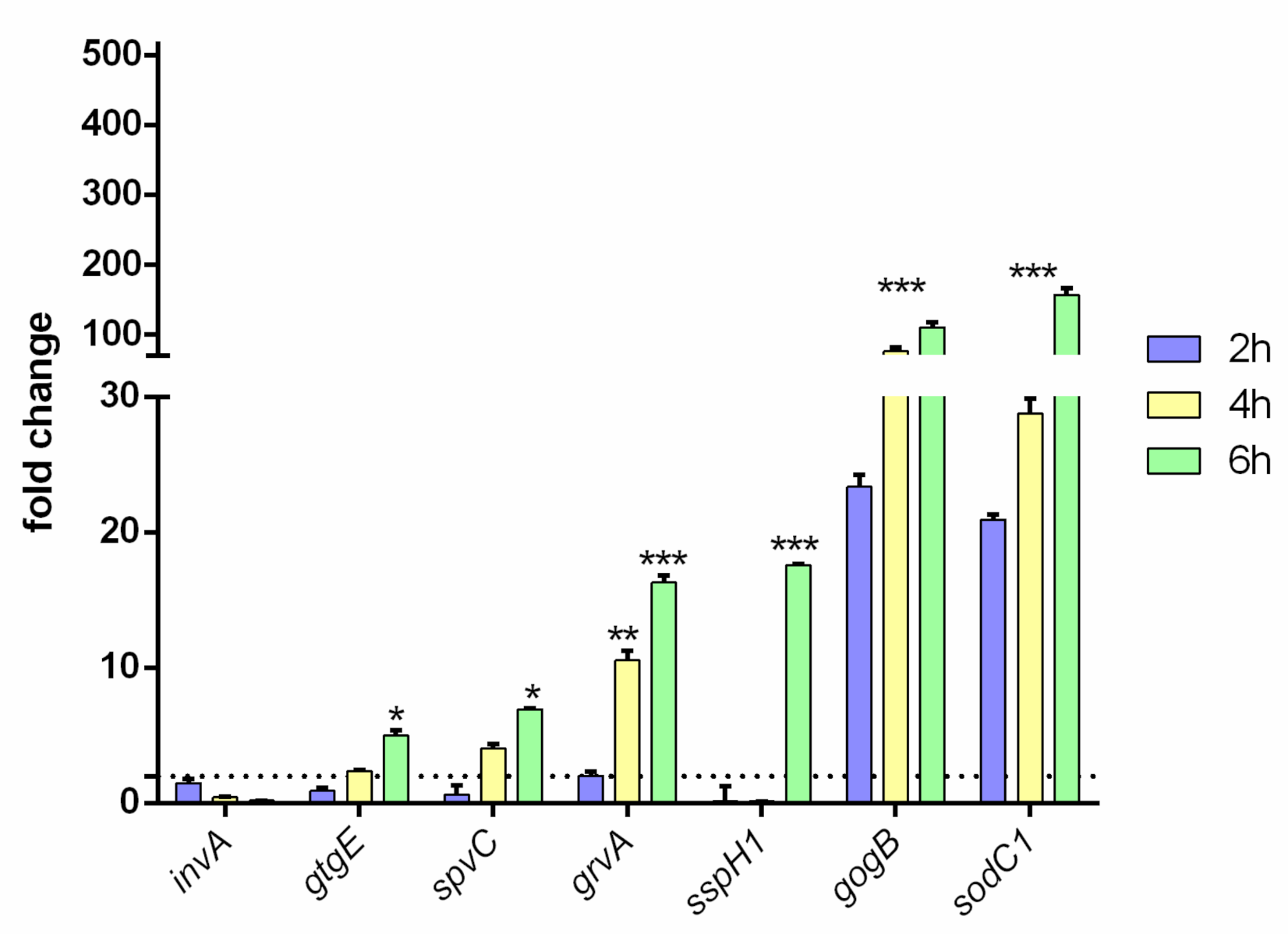
3.3. RW and RR Strains Exhibit Different Virulence Profiles
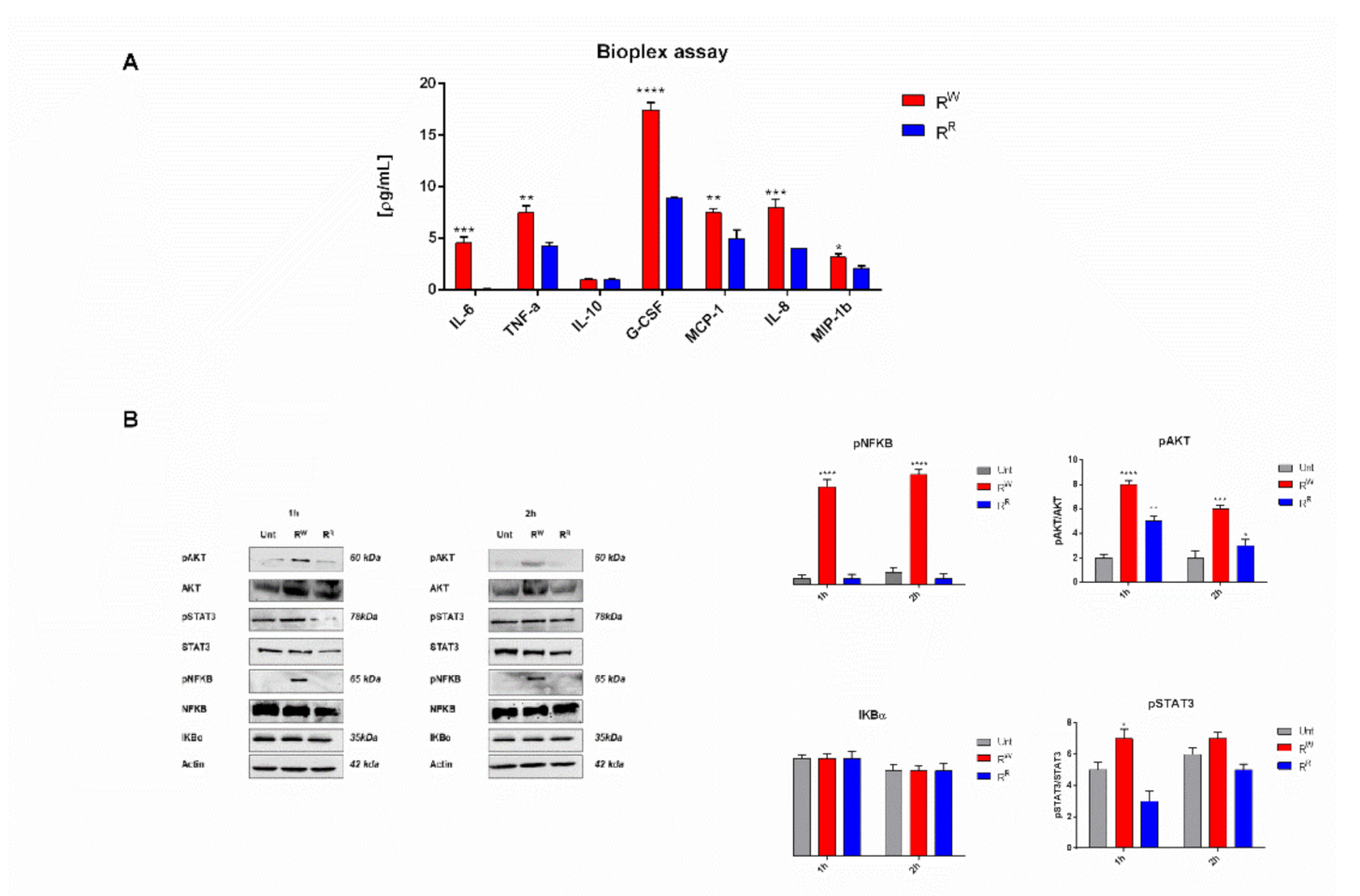
3.4. RW and RR Strains Induce a Different Inflammatory Host Response
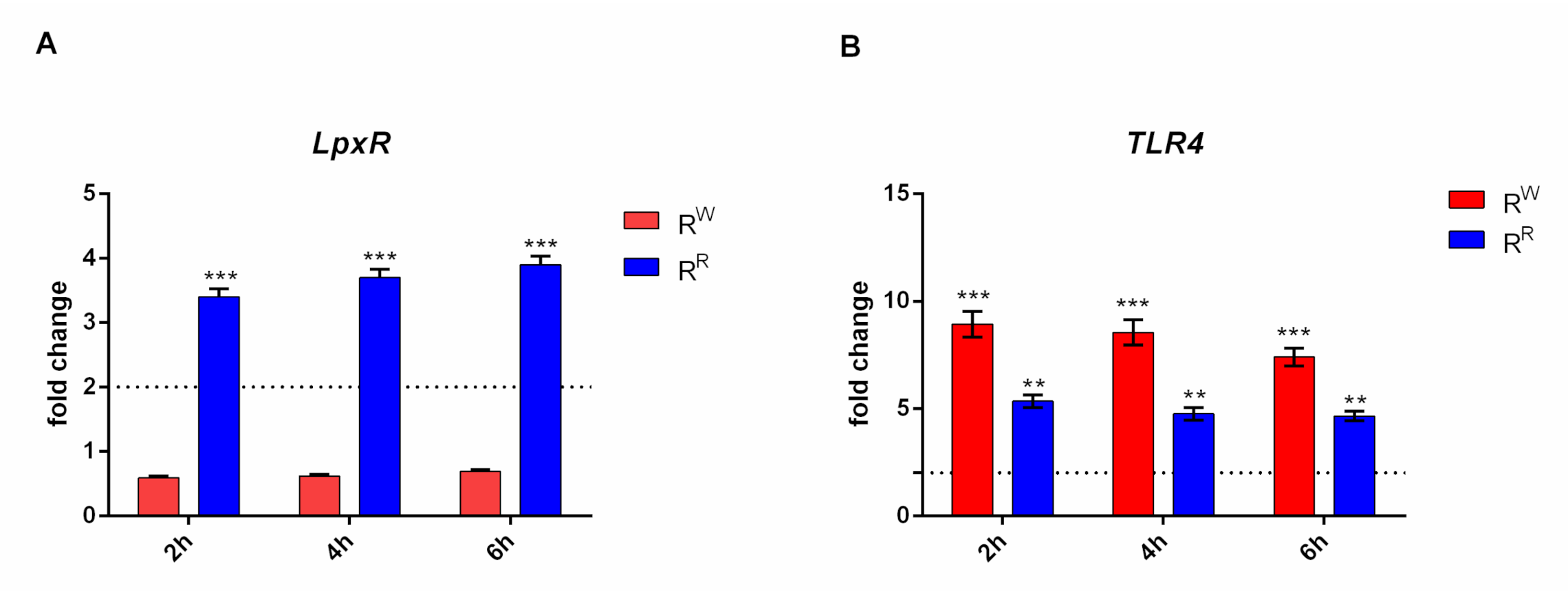
3.5. RW and RR Strains Display a Different LpxR and TLR4 Gene Expression Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chattaway, M.A.; Langridge, G.C.; Wain, J. Salmonella nomenclature in the genomic era: A time for change. Sci. Rep. 2021, 11, 1–8. [Google Scholar] [CrossRef]
- Gal-Mor, O.; Boyle, E.C.; Grassl, G.A. Same species, different diseases: How and why typhoidal and non-typhoidal Salmonella enterica serovars differ. Front. Microbiol. 2014, 5, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemyss, M.; Pearson, J.S. Host Cell Death Responses to Non-typhoidal Salmonella Infection. Front. Immunol. 2019, 10, 1758. [Google Scholar] [CrossRef]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, J.; Mourão, J.; Peixe, L.; Antunes, P. Non-typhoidal salmonella in the pig production chain: A comprehensive analysis of its impact on human health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The role of vaccines in combatting antimicrobial resistance. Nat. Rev. Genet. 2021, 19, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef]
- Worsena, C.R.; Miller, A.S.; King, M.A. Salmonella Infections. Pediatr. Rev. 2019, 40, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Hatfull, G.F. Dark Matter of the Biosphere: The amazing world of bacteriophage diversity. J. Virol. 2015, 89, 8107–8110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koskella, B.; Brockhurst, M.A. Bacteria–phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiol. Rev. 2014, 38, 916–931. [Google Scholar] [CrossRef] [Green Version]
- Rostøl, J.T.; Marraffini, L. (Ph)ighting Phages: How bacteria resist their parasites. Cell Host Microbe 2019, 25, 184–194. [Google Scholar] [CrossRef] [Green Version]
- Oechslin, F. Resistance development to bacteriophages occurring during bacteriophage therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.-Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.M.; Nikolich, M.P. Bacteriophage-Resistant Mutants in Yersinia pestis: Identification of Phage Receptors and Attenuation for Mice. PLoS ONE 2011, 6, e25486. [Google Scholar] [CrossRef] [Green Version]
- Capparelli, R.; Nocerino, N.; Lanzetta, R.; Silipo, A.; Amoresano, A.; Giangrande, C.; Becker, K.; Blaiotta, G.; Evidente, A.; Cimmino, A.; et al. Bacteriophage-resistant staphylococcus aureus mutant confers broad immunity against staphylococcal infection in mice. PLoS ONE 2010, 5, e11720. [Google Scholar] [CrossRef] [Green Version]
- Capparelli, R.; Nocerino, N.; Iannaccone, M.; Ercolini, D.; Parlato, M.; Chiara, M.; Iannelli, D. Bacteriophage Therapy of Salmonella enterica:A Fresh Appraisal of Bacteriophage Therapy. J. Infect. Dis. 2010, 201, 52–61. [Google Scholar] [CrossRef] [Green Version]
- Rakhuba, D.V.; Kolomiets, E.I.; Dey, E.S.; Novik, G.I. Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol. J. Microbiol. 2010, 59, 145–155. [Google Scholar] [CrossRef]
- Kulikov, E.E.; Golomidova, A.K.; Prokhorov, N.; Ivanov, P.A.; Letarov, A.V. High-throughput LPS profiling as a tool for revealing of bacteriophage infection strategies. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Papaianni, M.; Contaldi, F.; Fulgione, A.; Woo, S.L.; Casillo, A.; Corsaro, M.M.; Parrilli, E.; Marcolungo, L.; Rossato, M.; Delledonne, M.; et al. Role of phage ϕ1 in two strains of Salmonella rissen, sensitive and resistant to phage ϕ1. BMC Microbiol. 2018, 18, 208. [Google Scholar] [CrossRef]
- Ematsuura, M. Structural modifications of bacterial lipopolysaccharide that facilitate gram-negative bacteria evasion of host innate immunity. Front. Immunol. 2013, 4, 109. [Google Scholar] [CrossRef] [Green Version]
- Zariri, A.; Van Der Ley, P. Biosynthetically engineered lipopolysaccharide as vaccine adjuvant. Expert Rev. Vaccines 2015, 14, 861–876. [Google Scholar] [CrossRef]
- Roche, S.M.; Gracieux, P.; Milohanic, E.; Albert, I.; Virlogeux-Payant, I.; Témoin, S.; Grépinet, O.; Kerouanton, A.; Jacquet, C.; Cossart, P.; et al. Investigation of specific substitutions in virulence genes characterizing phenotypic groups of low-virulence field strains of listeria monocytogenes. Appl. Environ. Microbiol. 2005, 71, 6039–6048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borriello, G.; Lucibelli, M.G.; Pesciaroli, M.; Carullo, M.R.; Graziani, C.; Ammendola, S.; Battistoni, A.; Ercolini, D.; Pasquali, P.; Galiero, G. Diversity of Salmonella spp. serovars isolated from the intestines of water buffalo calves with gastroenteritis. BMC Vet. Res. 2012, 8, 201. [Google Scholar] [CrossRef] [Green Version]
- Spatuzza, C.; Schiavone, M.; Di Salle, E.; Janda, E.; Sardiello, M.; Fiume, G.; Fierro, O.; Simonetta, M.; Argiriou, N.; Faraonio, R.; et al. Physical and functional characterization of the genetic locus of IBtk, an inhibitor of Bruton’s tyrosine kinase: Evidence for three protein isoforms of IBtk. Nucleic Acids Res. 2008, 36, 4402–4416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Front. Plant Sci. 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hu, N.; Qian, L.; Hu, Y.; Shou, J.-Z.; Wang, C.; Giffen, C.; Wang, Q.-H.; Wang, Y.; Goldstein, A.M.; Emmert-Buck, M.; et al. Quantitative real-time RT-PCR validation of differential mRNA expression of SPARC, FADD, Fascin, COL7A1, CK4, TGM3, ECM1, PPL and EVPLin esophageal squamous cell carcinoma. BMC Cancer 2006, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Iannelli, D.; D’Apice, L.; Cottone, C.; Viscardi, M.; Scala, F.; Zoina, A.; Del Sorbo, G.; Spigno, P.; Capparelli, R. Simultaneous detection of cucumber mosaic virus, tomato mosaic virus and potato virus Y by flow cytometry. J. Virol. Methods 1997, 69, 137–145. [Google Scholar] [CrossRef]
- Lotze, M. Measuring Immunity: Basic Biology and Clinical Assessment; Elsevier Academic Press: San Diego, CA, USA; London, UK, 2005; ISBN 978-1-42-375541-8. [Google Scholar]
- Burmeister, A.R.; Fortier, A.; Roush, C.; Lessing, A.J.; Bender, R.G.; Barahman, R.; Grant, R.; Chan, B.K.; Turner, P.E. Pleiotropy complicates a trade-off between phage resistance and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2020, 117, 11207–11216. [Google Scholar] [CrossRef]
- Zhou, D.; Galán, J. Salmonella entry into host cells: The work in concert of type III secreted effector proteins. Microbes Infect. 2001, 3, 1293–1298. [Google Scholar] [CrossRef]
- Galán, J.E.; Ginocchio, C.; Costeas, P. Molecular and functional characterization of the Salmonella invasion gene invA: Homology of InvA to members of a new protein family. J. Bacteriol. 1992, 174, 4338–4349. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.D.; Slauch, J.M. Characterization of grvA, an Antivirulence Gene on the Gifsy-2 Phage in Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2001, 183, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Qazi, I.H.; Wang, L.; Zhou, G.; Han, H. Salmonella Virulence and Immune Escape. Microorganisms 2020, 8, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.D.; Figueroa-Bossi, N.; Wang, M.; Uzzau, S.; Bossi, L.; Slauch, J.M. Identification of GtgE, a Novel Virulence Factor Encoded on the Gifsy-2 Bacteriophage of Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2002, 184, 5234–5239. [Google Scholar] [CrossRef] [Green Version]
- Ranade, K.; Poteete, A.R. Superinfection exclusion (sieB) genes of bacteriophages P22 and lambda. J. Bacteriol. 1993, 175, 4712–4718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, R.; Zhang, Y.-G.; Sun, J. STAT3 activation in infection and infection-associated cancer. Mol. Cell. Endocrinol. 2017, 451, 80–87. [Google Scholar] [CrossRef]
- Reynolds, C.M.; Ribeiro, A.A.; McGrath, S.C.; Cotter, R.J.; Raetz, C.R.H.; Trent, M.S. An Outer Membrane Enzyme Encoded by Salmonella typhimurium lpxR That Removes the 3′-Acyloxyacyl Moiety of Lipid A. J. Biol. Chem. 2006, 281, 21974–21987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.; Lara-Tejero, M.; Waxham, M.N.; Li, W.; Hu, B.; Galán, J.E.; Liu, J. Visualization of the type III secretion mediated Salmonella–host cell interface using cryo-electron tomography. eLife 2018, 7. [Google Scholar] [CrossRef]
- Que, F.; Wu, S.; Huang, R. Salmonella Pathogenicity Island 1(SPI-1) at Work. Curr. Microbiol. 2013, 66, 582–587. [Google Scholar] [CrossRef]
- Rushing, M.D.; Slauch, J.M. Either periplasmic tethering or protease resistance is sufficient to allow a SodC to protect Salmonella enterica serovar Typhimurium from phagocytic superoxide. Mol. Microbiol. 2011, 82, 952–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M.; Rossi, A.; Amici, C. New embo member’s review: NF-kappaB and virus infection: Who controls whom. EMBO J. 2003, 22, 2552–2560. [Google Scholar] [CrossRef]
- Mukaida, N.; Harada, A.; Matsushima, K. Interleukin-8 (IL-8) and monocyte chemotactic and activating factor (MCAF/MCP-1), chemokines essentially involved in inflammatory and immune reactions. Cytokine Growth Factor Rev. 1998, 9, 9–23. [Google Scholar] [CrossRef]
- Maurer, M.; von Stebut, E. Macrophage inflammatory protein-1. Int. J. Biochem. Cell Biol. 2004, 36, 1882–1886. [Google Scholar] [CrossRef]
- Lissoni, P.; Messina, G.; Pelizzoni, F.; Rovelli, F.; Brivio, F.; Monzon, A.; Crivelli, N.; Lissoni, A.; Tassoni, S.; Sassola, A.; et al. The Fascination of Cytokine Immuno-logical Science. J. Infect. Rev. Artic. 2020, 3, 18–28. [Google Scholar]
- Arango Duque, G.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckmann, L.; Kagnoff, M.F. Cytokines in host defense against Salmonella. Microbes Infect. 2001, 3, 1191–1200. [Google Scholar] [CrossRef]
- Steward-Tharp, S.M.; Laurence, A.; Kanno, Y.; Kotlyar, A.; Villarino, A.V.; Sciume, G.; Kuchen, S.; Resch, W.; Wohlfert, E.A.; Jiang, K.; et al. A mouse model of HIES reveals pro- and anti-inflammatory functions of STAT3. Blood 2014, 123, 2978–2987. [Google Scholar] [CrossRef] [Green Version]
- León, M.; Bastías, R. Virulence reduction in bacteriophage resistant bacteria. Front. Microbiol. 2015, 6, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrell, K.F.; Kropinski, A. Isolation and characterization of a bacteriophage specific for the lipopolysaccharide of rough derivatives of Pseudomonas aeruginosa strain PAO. J. Virol. 1981, 38, 529–538. [Google Scholar] [CrossRef] [Green Version]
- Nijland, R.; Hofland, T.; Van Strijp, J.A.G. Recognition of LPS by TLR4: Potential for Anti-Inflammatory Therapies. Mar. Drugs 2014, 12, 4260–4273. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capparelli, R.; Cuomo, P.; Papaianni, M.; Pagano, C.; Montone, A.M.I.; Ricciardelli, A.; Iannelli, D. Bacteriophage-Resistant Salmonella rissen: An In Vitro Mitigated Inflammatory Response. Viruses 2021, 13, 2468. https://doi.org/10.3390/v13122468
Capparelli R, Cuomo P, Papaianni M, Pagano C, Montone AMI, Ricciardelli A, Iannelli D. Bacteriophage-Resistant Salmonella rissen: An In Vitro Mitigated Inflammatory Response. Viruses. 2021; 13(12):2468. https://doi.org/10.3390/v13122468
Chicago/Turabian StyleCapparelli, Rosanna, Paola Cuomo, Marina Papaianni, Cristina Pagano, Angela Michela Immacolata Montone, Annarita Ricciardelli, and Domenico Iannelli. 2021. "Bacteriophage-Resistant Salmonella rissen: An In Vitro Mitigated Inflammatory Response" Viruses 13, no. 12: 2468. https://doi.org/10.3390/v13122468
APA StyleCapparelli, R., Cuomo, P., Papaianni, M., Pagano, C., Montone, A. M. I., Ricciardelli, A., & Iannelli, D. (2021). Bacteriophage-Resistant Salmonella rissen: An In Vitro Mitigated Inflammatory Response. Viruses, 13(12), 2468. https://doi.org/10.3390/v13122468