
Synthetic Messenger RNA-Based Vaccines: From Scorn to Hype


{kind=link}
{kind=link}
Abstract
:1. Introduction
- -
- RNA contains genetic information and, thereby, is being used to allow for the organism to encode a specific protein of interest (Figure 1); this approach has been optimized for nearly 30 years to be used as a vaccine platform [2,3,4,5] and just reached approval in December 2020 (with the first vaccine against COVID-19);
- -
- RNA forming 3D structures, as in ribosomes or any ribo-nucleoprotein complexes, is used to generate aptamers. This led to the design of Macugen® (a structured 2′ Fluoro oligonucleotide, which binds and blocks VEGF) for the treatment of Age-Related Macular Degeneration;
- -
- RNA can perform enzymatic activities as evidenced for ribozymes (this technology has not yet been turned into an approved drug);
- -
- RNAs control gene expression, as seen with micro RNAs (miRNAs) that has been turned into a drug (siRNA) that degrades messenger RNA encoding transthyretin and it is used for the treatment of hereditary transthyretin amyloidosis.
2. History of Synthetic Non-Replicating mRNA Vaccines
3. Advantages of Synthetic mRNA Vaccines
- -
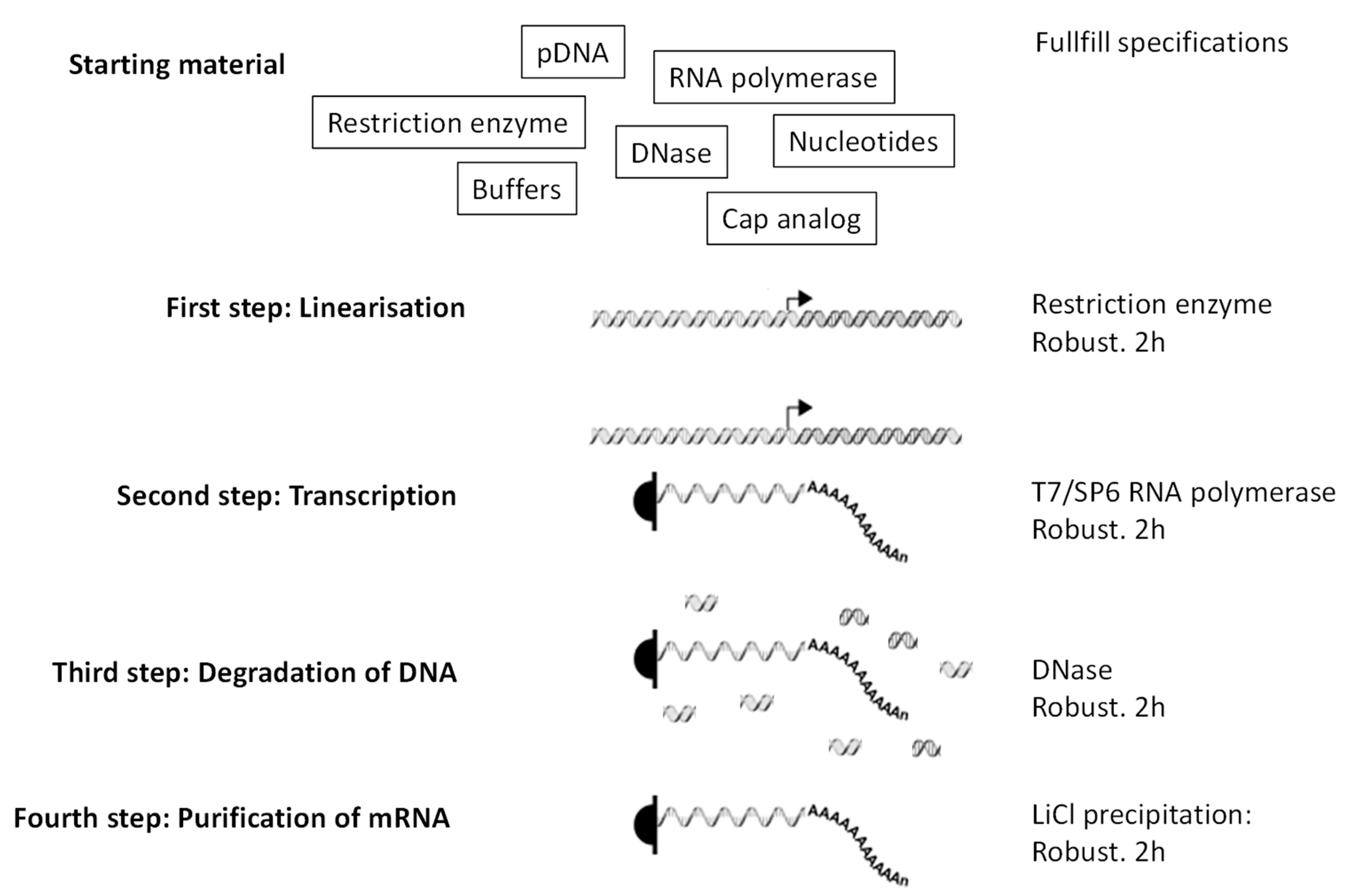
- Fast and easy production in pharmaceutical conditions. The steps leading to production of mRNA (Figure 2) are simple, fast, and most importantly robust, which means that they do not depend on the mRNA sequence. Thus, once the pharmaceutical process for mRNA production is established, it can be used for any RNA sequence coding any protein. That is not the case for vaccines that are based on peptides, proteins, nanoparticles, or viruses. In those cases, preexisting knowledge and infrastructures do not guarantee, in the context of a new pathogen/antigen, that production will be fast and successful. Peptides, depending on their sequences, can be very difficult to synthesize, to purify, or to store. Proteins, depending on their structure and glycosylation status, must be expressed by adequate organisms (to be individually defined) and their purification and preservation is a specific project for each new protein. Similarly, production, purification, and preservation of attenuated or inactivated viruses require extensive testing and optimization to define the methods that are adapted to the precise pathogen of interest. The versatility of mRNA production is key to producing individualized anti-cancer vaccines: tumor biopsies are sequenced, cancer-specific mutations identified, and ivt mRNA are produced in order to encode short proteins that contain identified mutations (http://merit-consortium.eu/about/ (accessed on 06 February 2021)). Each patient receives a personalized vaccine. The whole process, from the biopsy to the tailor-made mRNA vaccine, takes less than three months. Thus, mRNA companies are accustomed to producing ivt mRNA vaccines from whatever nucleotide sequence within less than three months.
- -
- Highly stable in vitro. Contrary to the general belief, RNA is very stable in vitro. It is degraded in the presence of RNases or at elevated pH. However, should the RNA be free of RNases (which is of course the case for a pharmaceutical ivt mRNA) and at neutral/slightly acidic pH, it is then extremely stable. It is the only biological molecule that can be precipitated, resuspended in water, heated up to 90 degrees Celsius, frozen, and lyophilized, all without damage. No other biological product is as robust (particularly concerning the elevated temperatures). In addition, lyophilized RNA can be stored at room temperature for a very long time. Being resuspended in water, it will again be immediately biologically active (independently of its sequence, i.e., independently of the encoded protein). On this basis, in 2014 CureVac received a special prize of two million Euros from the EU (https://ec.europa.eu/commission/presscorner/detail/en/IP_14_229 (accessed on 06 February 2021)). The prize was for innovative solutions for vaccine transportation and storage where the cold chain cannot be guaranteed.
- -
- Safe and defined therapeutic window. As opposed to DNA-based vaccines (i.e., plasmid DNA or DNA viruses), the ivt mRNA is, per nature, a transient molecule in vivo. It is totally degraded by abundant RNases outside and inside cells. It can also not affect the DNA genome of the host. For those reasons, ivt mRNA is particularly safe for therapies. There is another advantage of the very transient nature of ivt mRNA in vivo: after a time that can be evaluated, to a maximum of a few days to less than a week (depending on the mRNA sequence and delivery vehicle), it can be assumed that the ivt mRNA is no longer active. Thus, the therapeutic window is very well defined. Re-injections can be performed and they will not cumulate with previous injections. Therefore, there can not appear any surprising side effects that are associated with a cumulative aspect and individuals can be repeatedly injected, as required, for the efficacy of the treatment.
- -
- Mono-antigenic. Vaccines in the form of recombinant viruses express many different proteins, including the one of interest (SARS-CoV-2 Spike protein, for example). Thus, boost vaccines are additionally (i) triggering immunity against non-targeted antigens and can (ii) be neutralized by antibodies triggered by the prime against the vehicle. A similar problem can be seen with complex recombinant vaccines (virus-like particles) or even protein vaccines that, by nature, contain contaminants (e.g., degradation products, misfolded proteins, or impurities from production) that may trigger and boost immune responses that are not the ones of interest.
4. Design and Optimization of Non-Replicating Ivt mRNAs
- -
- Optimizing the mRNA. Four functional parts in the molecule can be defined and each can be independently optimized in order to lead to superlative mRNA molecules that are (i) more stable in the cytosol of cells and (ii) better translated, as shown in Figure 1. It should be noted that these two characteristics are correlated and that usually the most stable mRNAs are also best translated and vice-versa.
- -
- Optimizing the vehicle. Cationic compounds can bind and stabilize the anionic RNA. At the same time, they neutralize the molecule and generate particles, facilitating uptake by cells and the passage through cell membranes. Cationic liposomes, as well as cationic polymers, have been used from the beginning of ivt mRNA vaccine optimization (liposomes and Protamines in the study by Hoerr [9]). Optimized formulations need to incorporate two opposing features: a very stable and resistant structure that protects RNA outside of the cells and a very efficacious dissociation of the RNA from its carrier inside the cytosol. Should the interaction between RNA and carrier be too strong, the RNA may not be released in the cytosol, and should it be too weak, the particles may be instable in vitro as well as after the injection and release RNA before it gets into the cytosol. In addition, the vehicle must consist of safe compounds that are catabolized naturally. Cationic liposomes (especially those containing ionizable cationic lipids) have shown, so far, to be the best. Their development has been accelerated, since the beginning of the century, for the delivery of siRNA drugs. These siRNAs must very efficiently reach the cytosol of cells to be efficacious. Companies, such as Acuitas Therapeutics, have made enormous progress in liposomal formulations aimed at delivering RNA, which facilitated the design of Onpattro® (Alnylam Pharmaceuticals, Inc., Cambridge, MA, USA ), the first drug that is based on siRNA. As Onpattro® (Alnylam Pharmaceuticals, Inc., Cambridge, MA, USA ) treats a genetic defect, it must be administered frequently and, as it controls gene expression in a large organ, it also needs to be given at a high dose. Thus, the formulation had to be efficacious and non-toxic. It consists of a delivery of up to 30 miligrams of RNA every three weeks by intravenous injection. Based on this expertise, safe and efficacious formulations of ivt mRNA could be developed. The liposome that is used to deliver the first approved ivt mRNA drug, the vaccine from BioNTech/Pfizer, is based on the technologies that were developed by Acuitas Therapeutics. CureVac is also using Acuitas Therapeutics formulations for its ivt mRNA vaccine against COVID-19 in clinical studies. Details on the formulations that were used by BioNTech, CureVac and Moderna for their anti-COVID-19 vaccines are given in a recent review by Buschmann et al. [31].
5. Anti-SARS-CoV-2 Non-Replicating mRNA Vaccines
- -
- Moderna: injecting the first volunteer on 16 March 2020, Moderna was the first drug developer to start clinical testing of an anti-COVID-19 vaccine. The single vaccine consists of a PseudoUridine-modified mRNA coding for the full-length pre-fusion conformation spike in a liposomal shell, and it was produced in record speed. The company has published the results of its phase I dose escalation study in November 2020 [33]. The three tested doses (in three cohorts of 15 participants) of 25, 100, and 250 µg ivt mRNA formulated in liposomes and given intramuscular as a prime-boost regimen (with four weeks interval between injections), induced high neutralizing antibody titers (similar or higher than the titers found in the sera from patients having recovered from COVID-19). In general, there have been side effects of the vaccine in half of the participants, such as fatigue, chills, headache, myalgia, and pain at the injection site. These side effects were more pronounced after the boost injection and at the highest dose of 250 µg. Clinical laboratory values revealed no severe issues. The company selected the middle dose of 100 µg for further studies and completed the enrollment of 30,000 participants for its phase III study on 22 October 2020. A preliminary analysis by the company indicated a 95% efficacy of the vaccine in protecting against COVID-19: 196 COVID cases (185 in placebo), 30 severe (30 placebo). Meanwhile, studies in vaccinated and virus-challenged non-human primates demonstrated that the vaccine could prevent virus replication in the upper and lower airways [34]. Based on the data that were provided by the company, the US FDA approved the vaccine for emergency use. This is the second approval (after the BioNTech vaccine, see below) of an mRNA drug, and the second approved vaccine against SARS-CoV-2.
- -
- BioNTech: having a broad range of ivt mRNA technologies and wanting to provide the safest and most efficient vaccine against COVID-19, in January 2020 the company designed four different ivt mRNA vaccines: two mRNAs encoding the full-length pre-fusion conformation spike, one having pseudoUridines, and the other having unmodified Uridines (as there is currently no available evidence that, for this type of vaccine, whether one or the other version is advantageous), one mRNA coding for only the Receptor Binding Domain of the spike (as antibodies directed solely against this domain may be more potent and might avoid facilitated infection) and one corresponding to a self-amplifying mRNA (as this format requires much lower doses than non-replicating mRNA). Having started the first injection on 23 April 2020, BioNTech decided to move the pseudoU ivt mRNA coding for the full length pre-fusion conformation spike (BNT162b2) to its phase III study. The phase I data for this vaccine tested at doses of 10, 30, and 100 µg demonstrated seroconversion in all volunteers and high neutralizing antibody titers (again similar or higher than titers in the sera of patients having recovered from COVID-19), even in participants that were older than 65 years of age and, additionally, the vaccine showed a good safety profile [6]. Having announced their collaboration on 17 March 2020, Pfizer and BioNTech evaluated the vaccine together, strongly accelerating the recruitment of participants. The phase III study was completed as early as 18 November 2020. It involved over 40,000 participants and demonstrated 95% protection against COVID-19: 9 cases of COVID-19 at least seven days after the second dose were observed among vaccine recipients and 169 among placebo recipients. Related adverse events were observed for 21% of the vaccine recipients and 5% in the placebo group. Four vaccine-related serious adverse events were reported among vaccine recipients (shoulder injury that is related to vaccine administration, right axillary lymphadenopathy, paroxysmal ventricular arrhythmia, and right leg paresthesia). Severe fatigue was observed in approximately 4% of BNT162b2 recipients. Elevated temperatures (38.9 to 40 °C) were reported in 0.8% (vaccine) and 0.1% (placebo) recipients after the second dose. Based on these favorable safety and efficacy data, BNT162b2 became the first ever approved mRNA-based drug and first authorized vaccine against COVID-19 when the UK approved its implementation on 2 December 2020. Subsequently, the US have approved the vaccine under emergency use on 11 December 2020. On 19 December 2020, Switzerland granted a conditional marketing authorization to the BioNTech/Pfizer anti-SARS-CoV-2 mRNA vaccine that will be distributed under the name COMIRNATY. It is the first authorization worldwide that is not under emergency use.
- -
- CureVac: having started its phase I study in June 2020, the company uses a liposome (cholesterol, 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC), PEG-ylated lipid, and a cationic lipid) intramuscular delivery of an unmodified mRNA coding for the full-length Spike in its pre-fusion conformation. The dose escalation went from 2 µg up to 12 µg. It is a prime boost regimen with 28 days between injections. The study population was 248 adults that were aged 18 to 60 years old. The company communicated (although the corresponding article is not yet published in a peer reviewed journal https://www.medrxiv.org/content/10.1101/2020.11.09.20228551v1.full-text (accessed on 06 February 2021)) that the vaccine was well tolerated (although the frequency and severity of adverse events increased with the dose level) and induced neutralizing antibodies in all subjects at the dose of 12 µg. The vaccine could also boost pre-existing immune responses in people who have been previously infected by SARS-CoV-2. At the end of September 2020, CureVac’s phase II study started and it included older adult participants. On 14 December 2020, the company announced the start of its pivotal clinical study.
6. Production and Storage
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gilbert, W. Origin of life: The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Pascolo, S. Messenger RNA-based vaccines. Expert Opin. Biol. Ther. 2004, 4, 1285–1294. [Google Scholar] [CrossRef]
- Pascolo, S. Vaccination with messenger RNA. Methods Mol. Med. 2006, 127, 23–40. [Google Scholar] [PubMed]
- Pascolo, S. Vaccination with messenger RNA (mRNA). Handb. Exp. Pharmacol. 2008, 221–235. [Google Scholar] [CrossRef]
- Sahin, U.; Kariko, K.; Tureci, O. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Krishnan, S.; Lenzen, G.; Magné, R.; Gomard, E.; Guillet, J.G.; Lévy, J.P.; Meulien, P. Induction of virus-specific cytotoxic T lymphocytes in vivo by liposome-entrapped mRNA. Eur. J. Immunol. 1993, 23, 1719–1722. [Google Scholar] [CrossRef] [PubMed]
- Conry, R.M.; LoBuglio, A.F.; Wright, M.; Sumerel, L.; Pike, M.J.; Johanning, F.; Benjamin, R.; Lu, D.; Curiel, D.T. Characterization of a messenger RNA polynucleotide vaccine vector. Cancer Res. 1995, 55, 1397–1400. [Google Scholar]
- Hoerr, I.; Obst, R.; Rammensee, H.G.; Jung, G. In vivo application of RNA leads to induction of specific cytotoxic T lymphocytes and antibodies. Eur. J. Immunol. 2000, 30, 1–7. [Google Scholar] [CrossRef]
- Probst, J.; Weide, B.; Scheel, B.; Pichler, B.J.; Hoerr, I.; Rammensee, H.G.; Pascolo, S. Spontaneous cellular uptake of exogenous messenger RNA in vivo is nucleic acid-specific, saturable and ion dependent. Gene Ther. 2007, 14, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Rittig, S.M.; Haentschel, M.; Weimer, K.J.; Heine, A.; Muller, M.R.; Brugger, W.; Horger, M.S.; Maksimovic, O.; Stenzl, A.; Hoerr, I.; et al. Intradermal vaccinations with RNA coding for TAA generate CD8+ and CD4+ immune responses and induce clinical benefit in vaccinated patients. Mol. Ther. 2011, 19, 990–999. [Google Scholar] [CrossRef]
- Weide, B.; Carralot, J.P.; Reese, A.; Scheel, B.; Eigentler, T.K.; Hoerr, I.; Rammensee, H.G.; Garbe, C.; Pascolo, S. Results of the first phase I/II clinical vaccination trial with direct injection of mRNA. J. Immunother. 2008, 31, 180–188. [Google Scholar] [CrossRef]
- Weide, B.; Pascolo, S.; Scheel, B.; Derhovanessian, E.; Pflugfelder, A.; Eigentler, T.K.; Pawelec, G.; Hoerr, I.; Rammensee, H.G.; Garbe, C. Direct injection of protamine-protected mRNA: Results of a phase 1/2 vaccination trial in metastatic melanoma patients. J. Immunother. 2009, 32, 498–507. [Google Scholar] [CrossRef]
- Ceppi, M.; de Bruin, M.G.; Seuberlich, T.; Balmelli, C.; Pascolo, S.; Ruggli, N.; Wienhold, D.; Tratschin, J.D.; McCullough, K.C.; Summerfield, A. Identification of classical swine fever virus protein E2 as a target for cytotoxic T cells by using mRNA-transfected antigen-presenting cells. J. Gen. Virol. 2005, 86 Pt 9, 2525–2534. [Google Scholar] [CrossRef]
- Carralot, J.P.; Probst, J.; Hoerr, I.; Scheel, B.; Teufel, R.; Jung, G.; Rammensee, H.G.; Pascolo, S. Polarization of immunity induced by direct injection of naked sequence-stabilized mRNA vaccines. Cell. Mol. Life Sci. 2004, 61, 2418–2424. [Google Scholar] [CrossRef]
- Scheel, B.; Braedel, S.; Probst, J.; Carralot, J.P.; Wagner, H.; Schild, H.; Jung, G.; Rammensee, H.G.; Pascolo, S. Immunostimulating capacities of stabilized RNA molecules. Eur. J. Immunol. 2004, 34, 537–547. [Google Scholar] [CrossRef]
- Scheel, B.; Teufel, R.; Probst, J.; Carralot, J.P.; Geginat, J.; Radsak, M.; Jarrossay, D.; Wagner, H.; Jung, G.; Rammensee, H.G.; et al. Toll-like receptor-dependent activation of several human blood cell types by protamine-condensed mRNA. Eur. J. Immunol 2005, 35, 1557–1566. [Google Scholar] [CrossRef]
- Carralot, J.P.; Weide, B.; Schoor, O.; Probst, J.; Scheel, B.; Teufel, R.; Hoerr, I.; Garbe, C.; Rammensee, H.G.; Pascolo, S. Production and characterization of amplified tumor-derived cRNA libraries to be used as vaccines against metastatic melanomas. Genet. Vaccines Ther. 2005, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittig, S.M.; Haentschel, M.; Weimer, K.J.; Heine, A.; Müller, M.R.; Brugger, W.; Horger, M.S.; Maksimovic, O.; Stenzl, A.; Hoerr, I.; et al. Long-term survival correlates with immunological responses in renal cell carcinoma patients treated with mRNA-based immunotherapy. Oncoimmunology 2016, 5, e1108511. [Google Scholar] [CrossRef] [Green Version]
- Kübler, H.; Scheel, B.; Gnad-Vogt, U.; Miller, K.; Schultze-Seemann, W.; Vom Dorp, F.; Parmiani, G.; Hampel, C.; Wedel, S.; Trojan, L.; et al. Self-adjuvanted mRNA vaccination in advanced prostate cancer patients: A first-in-man phase I/IIa study. J. Immunothe.r Cancer 2015, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Kreiter, S.; Vormehr, M.; Van de Roemer, N.; Diken, M.; Löwer, M.; Diekmann, J.; Boegel, S.; Schrörs, B.; Vascotto, F.; Castle, J.C.; et al. Mutant MHC class II epitopes drive therapeutic immune responses to cancer. Nature 2015, 520, 692–696. [Google Scholar] [CrossRef] [Green Version]
- Kranz, L.M.; Diken, M.; Haas, H.; Kreiter, S.; Loquai, C.; Reuter, K.C.; Meng, M.; Fritz, D.; Vascotto, F.; Hefesha, H.; et al. Systemic RNA delivery to dendritic cells exploits antiviral defence for cancer immunotherapy. Nature 2016, 534, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.A.; Fuhr, R.; Smolenov, I.; Ribeiro, A.M.; Panther, L.; Watson, M.; Senn, J.J.; Smith, M.; Almarsson, Ö.; Pujar, H.S.; et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine 2019, 37, 3326–3334. [Google Scholar] [CrossRef]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: An open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef]
- Elfakess, R.; Dikstein, R. A translation initiation element specific to mRNAs with very short 5’UTR that also regulates transcription. PLoS ONE 2008, 3, e3094. [Google Scholar] [CrossRef] [Green Version]
- Castelo-Szekely, V.; De Matos, M.; Tusup, M.; Pascolo, S.; Ule, J.; Gatfield, D. Charting DENR-dependent translation reinitiation uncovers predictive uORF features and links to circadian timekeeping via Clock. Nucleic Acids Res. 2019, 47, 5193–5209. [Google Scholar] [CrossRef] [Green Version]
- Tusup, M.; Kundig, T.; Pascolo, S. An eIF4G-recruiting aptamer increases the functionality of in vitro transcribed mRNA. Int. J. Med. Health Sci. 2018, 4. [Google Scholar]
- Sample, P.J.; Wang, B.; Reid, D.W.; Presnyak, V.; McFadyen, I.J.; Morris, D.R.; Seelig, G. Human 5′ UTR design and variant effect prediction from a massively parallel translation assay. Nat. Biotechnol. 2019, 37, 803–809. [Google Scholar] [CrossRef]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Türeci, O.; Sahin, U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef] [PubMed]
- von Niessen, A.G.; Poleganov, M.A.; Rechner, C.; Plaschke, A.; Kranz, L.M.; Fesser, S.; Diken, M.; Löwer, M.; Vallazza, B.; Beissert, T.; et al. Improving mRNA-Based Therapeutic Gene Delivery by Expression-Augmenting 3′ UTRs Identified by Cellular Library Screening. Mol. Ther. 2018, 27, 824–836. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, M.D.; Carrasco, M.J.; Alishetty, S.; Paige, M.; Alameh, M.G.; Weissman, D. Nanomaterial Delivery Systems for mRNA Vaccines. Vaccines 2021, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Pallesen, J.; Wang, N.; Corbett, K.S.; Wrapp, D.; Kirchdoerfer, R.N.; Turner, H.L.; Cottrell, C.A.; Becker, M.M.; Wang, L.; Shi, W.; et al. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA 2017, 114, E7348–E7357. [Google Scholar] [CrossRef] [Green Version]
- Jackson, L.A.; Anderson, E.J.; Rouphael, N.G.; Roberts, P.C.; Makhene, M.; Coler, R.N.; McCullough, M.P.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; et al. An mRNA Vaccine against SARS-CoV-2—Preliminary Report. N. Engl. J. Med. 2020, 383, 1920–1931. [Google Scholar] [CrossRef]
- Corbett, K.S.; Flynn, B.; Foulds, K.E.; Francica, J.R.; Boyoglu-Barnum, S.; Werner, A.P.; Flach, B.; O’Connell, S.; Bock, K.W.; Minai, M.; et al. Evaluation of the mRNA-1273 Vaccine against SARS-CoV-2 in Nonhuman Primates. N. Engl. J. Med. 2020, 383, 1544–1555. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pascolo, S. Synthetic Messenger RNA-Based Vaccines: From Scorn to Hype. Viruses 2021, 13, 270. https://doi.org/10.3390/v13020270
Pascolo S. Synthetic Messenger RNA-Based Vaccines: From Scorn to Hype. Viruses. 2021; 13(2):270. https://doi.org/10.3390/v13020270
Chicago/Turabian StylePascolo, Steve. 2021. "Synthetic Messenger RNA-Based Vaccines: From Scorn to Hype" Viruses 13, no. 2: 270. https://doi.org/10.3390/v13020270
APA StylePascolo, S. (2021). Synthetic Messenger RNA-Based Vaccines: From Scorn to Hype. Viruses, 13(2), 270. https://doi.org/10.3390/v13020270