
Vector Competence of Florida Culicoides insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2

 , , ,
, , ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
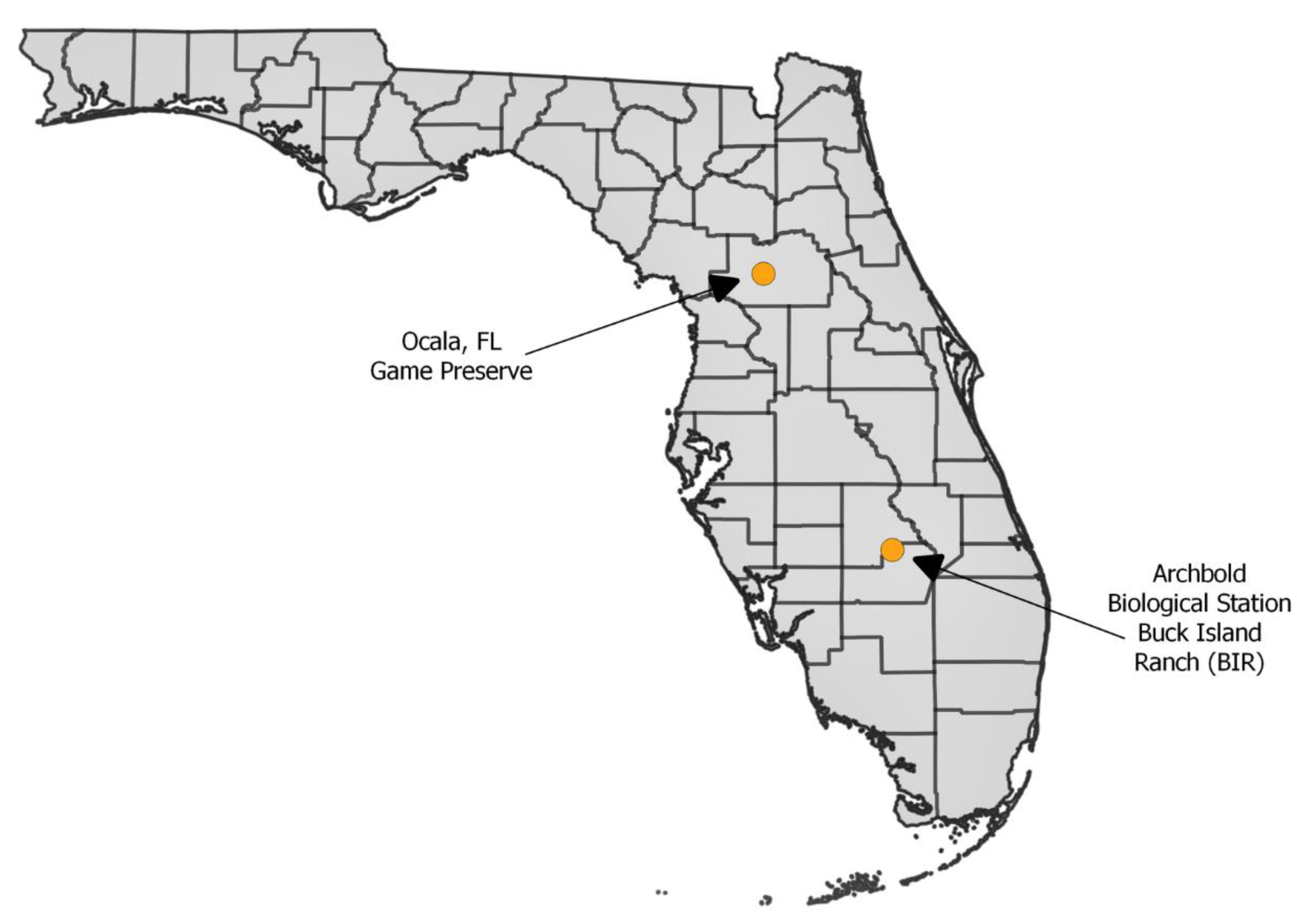
2.1. Field Collections of Culicoides insignis
2.2. Viral Screening of Field-Collected Midges
2.3. Per Os Infections of Culicoides
2.4. Midge Tissue Collection and Processing
2.5. Statistical Analysis
2.6. Intrathoracic Inoculation Assays
3. Results
3.1. Viral Screening of Pooled Field-Collected Midges
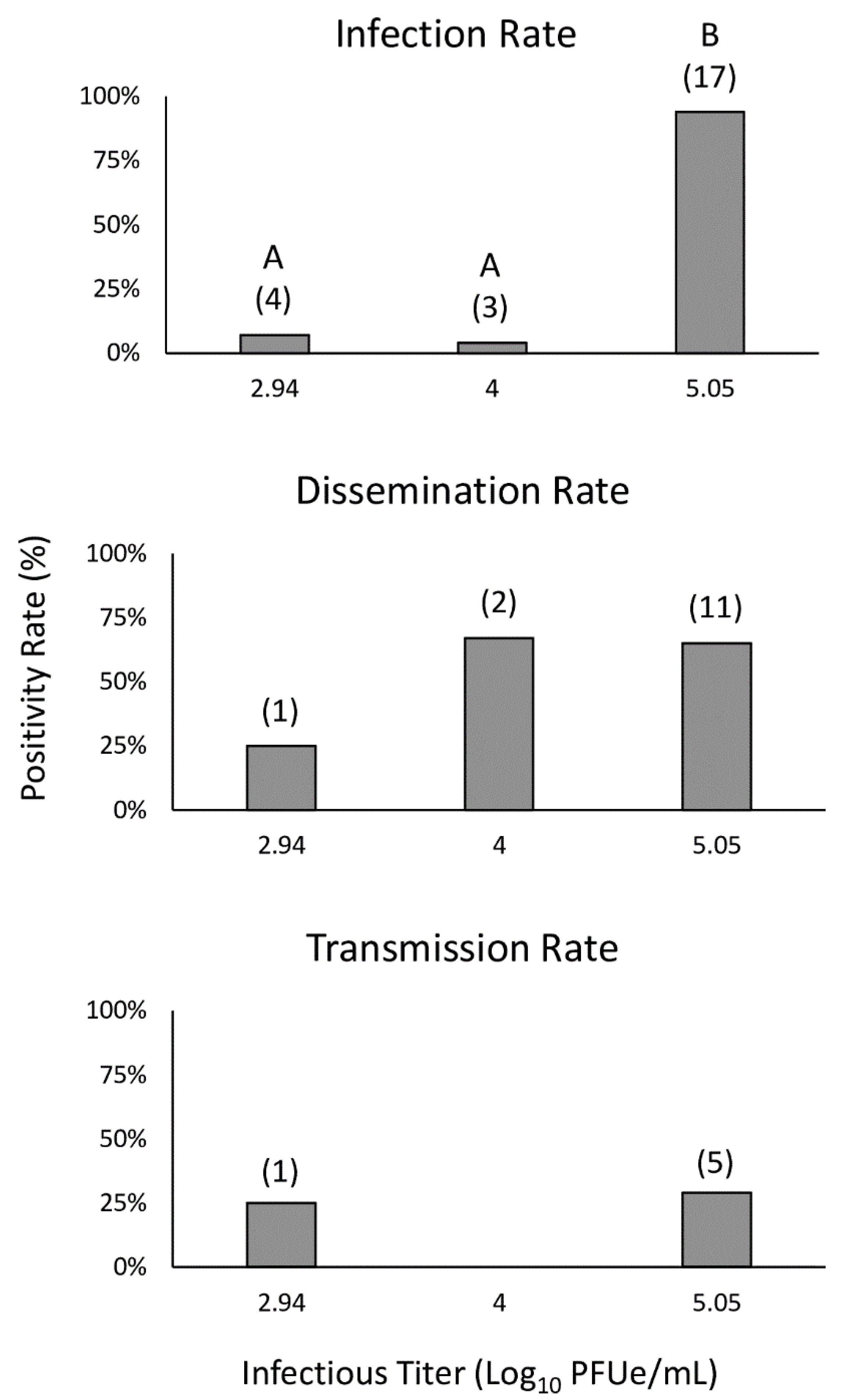
3.2. Infection, Dissemination, and Transmission Potential of C. insignis
3.3. Infectious Titer Comparisons
3.4. Intrathoracic Inoculation Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savini, G.; Afonso, A.; Mellor, P.; Aradaib, I.; Yadin, H.; Sanaa, M.; Wilson, W.; Monaco, F.; Domingo, M. Epizootic hemorrhagic disease. Res. Vet. Sci. 2011, 91, 1–17. [Google Scholar] [CrossRef]
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current perspectives, research gaps, and future directions. Vector-Borne Zoonotic Dis. 2015, 15, 348–363. [Google Scholar] [CrossRef]
- Haigh, J.; Mackintosh, C.; Griffin, F. Viral, parasitic and prion diseases of farmed deer and bison. Rev. Sci. Tech. L’OIE 2002, 21, 219–248. [Google Scholar] [CrossRef] [Green Version]
- Kedmi, M.; Van Straten, M.; Ezra, E.; Galon, N.; Klement, E. Assessment of the productivity effects associated with epizootic hemorrhagic disease in dairy herds. J. Dairy Sci. 2010, 93, 2486–2495. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.M.; Breckon, R.D.; Luedke, A.J.; Jones, R.H.; Metcalf, H.E. Transmission of two strains of Epizootic Hemorrhagic Disease Virus in deer by Culicoides variipennis. J. Wildl. Dis. 1977, 13, 9–16. [Google Scholar] [CrossRef]
- Jones, R.H.; Roughton, R.D.; Foster, N.M.; Bando, B.M. Culicoides, the vector of epizootic hemorrhagic disease in white-tailed deer in kentucky in 1971. J. Wildl. Dis. 1977, 13, 2–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruder, M.G.; Stallknecht, D.E.; Howerth, E.W.; Carter, D.L.; Pfannenstiel, R.S.; Allison, A.B.; Mead, D.G. Effect of temperature on replication of epizootic hemorrahgic disease viruses in Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2015, 52, 1050–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruder, M.G.; Stallknecht, D.E.; Allison, A.B.; Mead, D.G.; Carter, D.L.; Howerth, E.W. Host and potential vector susceptibility to an emerging orbivirus in the United States: Epizootic hemorrhagic disease virus serotype 6. Vet. Pathol. 2016, 53, 574–584. [Google Scholar] [CrossRef] [Green Version]
- Ruder, M.G.; Allison, A.B.; Stallknecht, D.E.; Mead, D.G.; McGraw, S.M.; Carter, D.L.; Kubiski, S.V.; Batten, C.A.; Klement, E.; Howerth, E.W. Susceptibility of white-tailed deer (Odocoileus virginianus) to experimental infection with epizootic hemorrhagic disease virus serotype 7. J. Wildl. Dis. 2012, 48, 676–685. [Google Scholar] [CrossRef] [Green Version]
- Borkent, A.; Grogan, W.L. Catalog of the new world biting midges north of Mexico (Diptera: Ceratopogonidae). Zootaxa 2009, 2273, 1–48. [Google Scholar] [CrossRef]
- Vigil, S.L.; Wlodkowski, J.C.; Parris, J. New records of biting midges of the genus Culicoides Latreille from the southeastern United States (Diptera: Ceratopogonidae). Insecta Mundi 2014, 394, 1–14. [Google Scholar]
- Jewiss-Gaines, A.; Barelli, L.; Hunter, F.F. First records of Culicoides sonorensis (Diptera: Ceratopogonidae), a known vector of Bluetongue Virus, in Southern Ontario. J. Med. Èntomol. 2016, 54, 757–762. [Google Scholar] [CrossRef]
- Smith, K.E.; Stallknecht, D.E. Culicoides (Diptera: Ceratopogonidae) collected during epizootics of hemorrhagic disease among captive white-tailed deer. J. Med. Èntomol. 1996, 33, 507–510. [Google Scholar] [CrossRef]
- Smith, K.E.; Stallknecht, D.E.; Sewell, C.T.; Rollor, E.A.; Mullen, G.R.; Anderson, R.R. Monitoring of Culicoides spp. at a site enzootic for hemorrhagic disease in white-tailed deer in Georgia, USA. J. Wildl. Dis. 1996, 32, 627–642. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Runkel IV, A.E.; Wisely, S.M.; Burkett-Cadena, N.D. Vertical stratification of Culicoides biting midges at a Florida big game preserve. Parasites Vectors 2018, 11, 505. [Google Scholar] [CrossRef] [Green Version]
- Sloyer, K.E.; Wisely, S.M.; Burkett-Cadena, N.D. Effects of ultraviolet LED versus incandescent bulb and carbon dioxide for sampling abundance and diversity of Culicoides in Florida. J. Med. Èntomol. 2019, 56, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.H.; Schmidtmann, E.T.; Foster, N.M. Vector-competence studies for bluetongue and epizootic hemorrhagic disease viruses with Culiocides venustus (Ceratopogonidae). Mosq. News 1983, 43, 184–186. [Google Scholar]
- Mullen, G.R.; Hayes, M.E.; Nusbaum, K.E. Potential vectors of bluetongue and epizootic hemorrhagic disease viruses of cattle and white-tailed deer in Alabama. Prog. Clin. Boil. Res. 1985, 178, 201–206. [Google Scholar]
- Pfannenstiel, R.S.; Mullens, B.A.; Ruder, M.G.; Zurek, L.; Cohnstaedt, L.W.; Nayduch, D. Management of North American Culicoides biting midges: Current knowledge and research needs. Vector-Borne Zoonotic Dis. 2015, 15, 374–384. [Google Scholar] [CrossRef]
- McGregor, B.L.; Stenn, T.; Sayler, K.A.; Blosser, E.M.; Blackburn, J.K.; Wisely, S.M.; Burkett-Cadena, N.D. Host use patterns of Culicoides spp. biting midges at a big game preserve in Florida, U.S.A. and implications for the transmission of orbiviruses. Med. Vet. Èntomol. 2018, 33, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Kramer, W.; Greiner, E.; Gibbs, E. A survey of Culicoides midges (Diptera: Ceratopogonidae) associated with cattle operations in Florida, USA. J. Med. Èntomol. 1985, 22, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Greiner, E.C.; Knausenberger, W.I.; Messersmith, M.; Kramer, W.L.; Gibbs, E.P.J. Culicoides spp. (Diptera: Ceratopogonidae) associated with cattle in St. Croix, Virgin Islands, and their relevance to Bluetongue virus. J. Med. Èntomol. 1990, 27, 1071–1077. [Google Scholar] [CrossRef]
- Vigil, S.L.; Ruder, M.G.; Shaw, D.; Wlodkowski, J.; Garrett, K.; Walter, M.; Corn, J.L. Apparent range expansion of Culicoides (Hoffmania) insignis (Diptera: Ceratopogonidae) in the Southeastern United States. J. Med. Èntomol. 2018, 55, 1043–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestwood, A.K.; Kistner, T.P.; Kellogg, F.E.; Hayes, F.A. The 1971 outbreak of hemorrhagic disease among white-tailed deer of the southeastern United States. J. Wildl. Dis. 1974, 10, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Tanya, V.; Greiner, E.; Gibbs, E. Evaluation of Culicoides insignis (Diptera: Ceratopogonidae) as a vector of bluetongue virus. Vet. Microbiol. 1992, 32, 1–14. [Google Scholar] [CrossRef]
- Wilson, W. Molecular comparison of VP3 from bluetongue and epizootic hemorrhagic disease viruses. Virus Res. 1991, 21, 225–236. [Google Scholar] [CrossRef]
- Moore, D.L.; Lee, V.H. Antigenic relationship between the virus of epizootic haemorrhagic disease of deer and bluetongue virus. Arch. Virol. 1972, 37, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.M.; Jones, R.H.; Luedke, A.J. Transmission of attenuated and virulent bluetongue virus with Culicoides variipennis infected orally via sheep. J. Vet. Res. 1968, 19, 275–279. [Google Scholar]
- Parsonson, I.; Snowdon, W. Bluetongue, epizootic hemorrhagic disease of deer and related viruses: Current situation in Australia. Prog. Clin. Biol. Res. 1985, 178, 27–35. [Google Scholar] [PubMed]
- Muller, M. Transmission and in vitro cxcretion of Bluetongue virus serotype 1 by inoculated Culicoides brevitarsis (Diptera: Ceratopogonidae). J. Med. Èntomol. 1987, 24, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Paweska, J.T.; Venter, G.J.; Guillet, P.; Hamblin, C. A comparison of the susceptibility of Culicoides imicola and C. bolitinos to oral infection with either serotypes of epizootic haemorrhagic disease virus. Med. Vet. Entomol. 2005, 19, 200–207. [Google Scholar] [CrossRef]
- Venter, G.J.; Mellor, P.S.; Paweska, J.T. Oral susceptibility of South African stock-associated Culicoides species to bluetongue virus. Med. Vet. Èntomol. 2006, 20, 329–334. [Google Scholar] [CrossRef]
- Erram, D.; Burkett-Cadena, N. Laboratory studies on the oviposition stimuli of Culicoides stellifer (Diptera: Ceratopogonidae), a suspected vector of Orbiviruses in the United States. Parasites Vectors 2018, 11, 300. [Google Scholar] [CrossRef] [Green Version]
- McGregor, B.L.; Erram, D.; Acevedo, C.; Alto, B.W.; Burkett-Cadena, N.D. Vector competence of Culicoides sonorensis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease virus serotype 2 Strains from Canada and Florida. Viruses 2019, 11, 367. [Google Scholar] [CrossRef] [Green Version]
- Wernike, K.; Hoffmann, B.; Beer, M. Simultaneous detection of five notifiable viral diseases of cattle by single-tube multiplex real-time RT-PCR. J. Virol. Methods 2015, 217, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.C.; O’Hearn, E.S.; Tellgren-Roth, C.; Stallknecht, D.E.; Mead, D.G.; Mecham, J.O. Detection of all eight serotypes of Epizootic hemorrhagic disease virus by real-time reverse transcription polymerase chain reaction. J. Vet. Diagn. Investig. 2009, 21, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, G.J.; Hill, E.; Pajor, I.T.; Nevill, E.M. The use of a membrane feeding technique to determine the infection rate of Culicoides imicola (Diptera, Ceratopogonidae) for 2 bluetongue virus serotypes in South Africa. Onderstepoort J. Vet. Res. 1991, 58, 5–9. [Google Scholar]
- Blanton, F.S.; Wirth, W.W. The Sand Flies (Culicoides) of Florida (Diptera: Ceratopogonidae); Florida Department of Agriculture and Consumer Services: Gainesville, FL, USA, 1979. [Google Scholar]
- Foster, N.M.; Jones, R.H.; McCrory, B.R. Preliminary investigations on insect transmission of Bluetongue virus in sheep. Am. J. Vet. Res. 1963, 24, 1195–1200. [Google Scholar] [PubMed]
- Richards, S.L.; Anderson, S.L.; Alto, B.W. Vector competence of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) for dengue virus in the Florida Keys. J. Med. Entomol. 2012, 49, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.L.; Huang, Y.-J.S.; Lyons, A.C.; Alto, B.W.; Unlu, I.; Higgs, S.; VanLandingham, D.L. North American Culex pipiens and Culex quinquefasciatus are competent vectors for Usutu virus. PLOS Negl. Trop. Dis. 2018, 12, e0006732. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Moncayo, A.C.; Edman, J.D.; Turell, M.J. Effect of Eastern Equine Encephalomyelitis Virus on the survival of Aedes albopictus, Anopheles quadrimaculatus, and Coquillettidia perturbans (Diptera: Culicidae). J. Med. Èntomol. 2000, 37, 701–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Leake, C.J.; Mertens, P.P.C.; Mellor, P.S. The barriers to bluetongue virus infection, dissemination and transmission in the vector, Culicoides variipennis (Diptera: Ceratopogonidae). Arch. Virol. 1999, 144, 747–761. [Google Scholar] [CrossRef]
- Mendiola, S.Y.; Mills, M.K.; Maki, E.; Drolet, B.S.; Wilson, W.C.; Berghaus, R.; Stallknecht, D.E.; Breitenbach, J.; McVey, D.S.; Ruder, M.G. EHDV-2 Infection prevalence varies in Culicoides sonorensis after feeding on infected shite-tailed deer over the course of viremia. Viruses 2019, 11, 371. [Google Scholar] [CrossRef] [Green Version]
- Ruder, M.G.; Howerth, E.W.; Stallknecht, D.E.; Allison, A.B.; Carter, D.L.; Drolet, B.S.; Klement, E.; Mead, D.G. Vector competence of Culicoides sonorensis (Diptera: Ceratopogonidae) to epizootic hemorrhagic disease virus serotype 7. Parasites Vectors 2012, 5, 236. [Google Scholar] [CrossRef] [Green Version]
- Mills, M.K.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of epizootic hemorrhagic disease virus infection within the vector, Culicoides sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.E.; Stallknecht, D.E.; Nettles, V.F. Experimental infection of Culicoides lahillei (Diptera: Ceratopogonidae) with epizootic hemorrhagic disease virus serotype 2 (Orbivirus: Reoviridae). J. Med. Entomol. 1996, 33, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.M.; Jones, R.H. Bluetongue Virus transmission with Culicoides variipennis via embryonating chicken eggs. J. Med. Èntomol. 1973, 10, 529–532. [Google Scholar] [CrossRef]
- Venter, G.J.; Graham, S.D.; Hamblin, C. African horse sickness epidemiology: Vector competence of South African Culicoides species for virus serotypes 3, 5 and 8. Med. Vet. Èntomol. 2000, 14, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Venter, G.J.; Wright, I.M.; Van Der Linde, T.C.; Paweska, J.T. The oral susceptibility of South African field populations of Culicoides to African horse sickness virus. Med. Vet. Entomol. 2009, 23, 367–378. [Google Scholar] [CrossRef]
- Garvin, M.C.; Greiner, E.C. Ecology of Culicoides (diptera: Ceratopogonidae) in southcentral Florida and experimental Culicoides vectors of the avian hematozoan Haemoproteus danilewskyi kruse. J. Wildl. Dis. 2003, 39, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.H.; Foster, N.M. Heterogeneity of Culicoides Variipennis field populations to oral infection with Bluetongue Virus. Am. J. Trop. Med. Hyg. 1978, 27, 178–183. [Google Scholar] [CrossRef] [PubMed]
- McGregor, B.L.; Sloyer, K.E.; Sayler, K.A.; Goodfriend, O.; Krauer, J.M.C.; Acevedo, C.; Zhang, X.; Mathias, D.; Wisely, S.M.; Burkett-Cadena, N.D. Field data implicating Culicoides stellifer and Culicoides venustus (Diptera: Ceratopogonidae) as vectors of epizootic hemorrhagic disease virus. Parasites Vectors 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Overall Rates | % Adjusted Rates | |||||||
|---|---|---|---|---|---|---|---|---|
| Trial | Population | Titer | N | IR a | DR a | TR a | DR b | TR b |
| 1 | Buck Island Ranch | 5.05 | 18 | 94.4 (17) | 61.1 (11) | 27.8 (5) | 64.7 (11) | 29.4 (5) |
| 2 | Ocala | 4.00 | 70 | 4.3 (3) | 2.9 (2) | 0.0 (0) | 66.7 (2) | 0.0 (0) |
| 3 | Ocala | 2.94 | 54 | 7.4 (4) | 1.9 (1) | 1.9 (1) | 25.0 (1) | 25.0 (1) |
| MI | Buck Island Ranch | 5.00 | 12 | - | - | 41.7 (5) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGregor, B.L.; Erram, D.; Alto, B.W.; Lednicky, J.A.; Wisely, S.M.; Burkett-Cadena, N.D. Vector Competence of Florida Culicoides insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2. Viruses 2021, 13, 410. https://doi.org/10.3390/v13030410
McGregor BL, Erram D, Alto BW, Lednicky JA, Wisely SM, Burkett-Cadena ND. Vector Competence of Florida Culicoides insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2. Viruses. 2021; 13(3):410. https://doi.org/10.3390/v13030410
Chicago/Turabian StyleMcGregor, Bethany L., Dinesh Erram, Barry W. Alto, John A. Lednicky, Samantha M. Wisely, and Nathan D. Burkett-Cadena. 2021. "Vector Competence of Florida Culicoides insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2" Viruses 13, no. 3: 410. https://doi.org/10.3390/v13030410
APA StyleMcGregor, B. L., Erram, D., Alto, B. W., Lednicky, J. A., Wisely, S. M., & Burkett-Cadena, N. D. (2021). Vector Competence of Florida Culicoides insignis (Diptera: Ceratopogonidae) for Epizootic Hemorrhagic Disease Virus Serotype-2. Viruses, 13(3), 410. https://doi.org/10.3390/v13030410