
Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review

 , ,
, ,  ,
,
Abstract
:1. Introduction
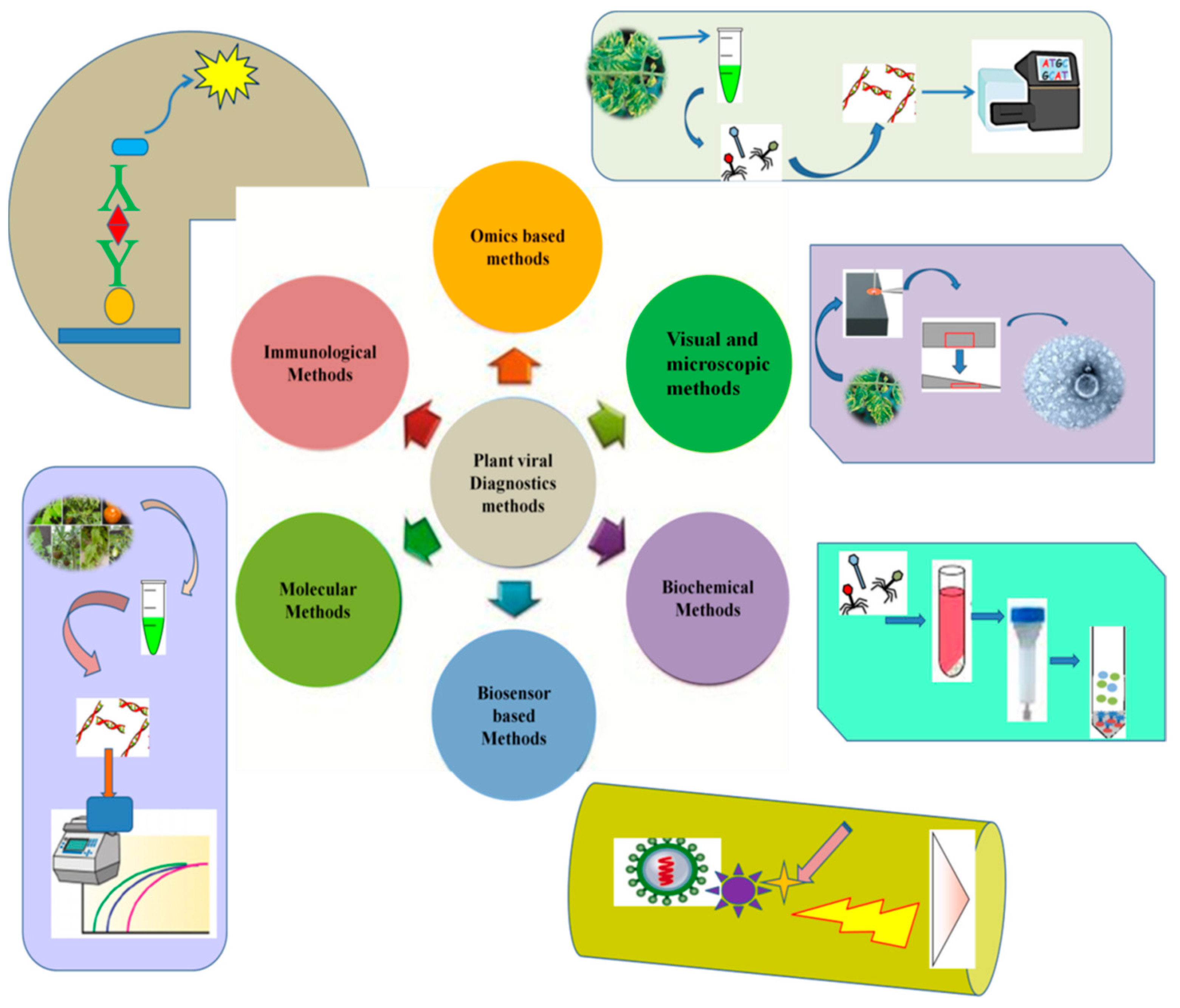
2. Methods for Plant Viral Diagnostic
2.1. Visual Inspections and Indicator Plants
2.2. Microscopic Methods
2.3. Serological Methods
2.4. Nucleic Acid-Based Methods
2.4.1. PCR-Based Methods
2.4.2. Next-Generation Sequencing (omics)-Based Methods
2.4.3. Analysis Using Second-Generation Sequencing Technologies
2.4.4. Analysis Using Third-Generation Sequencing Technologies
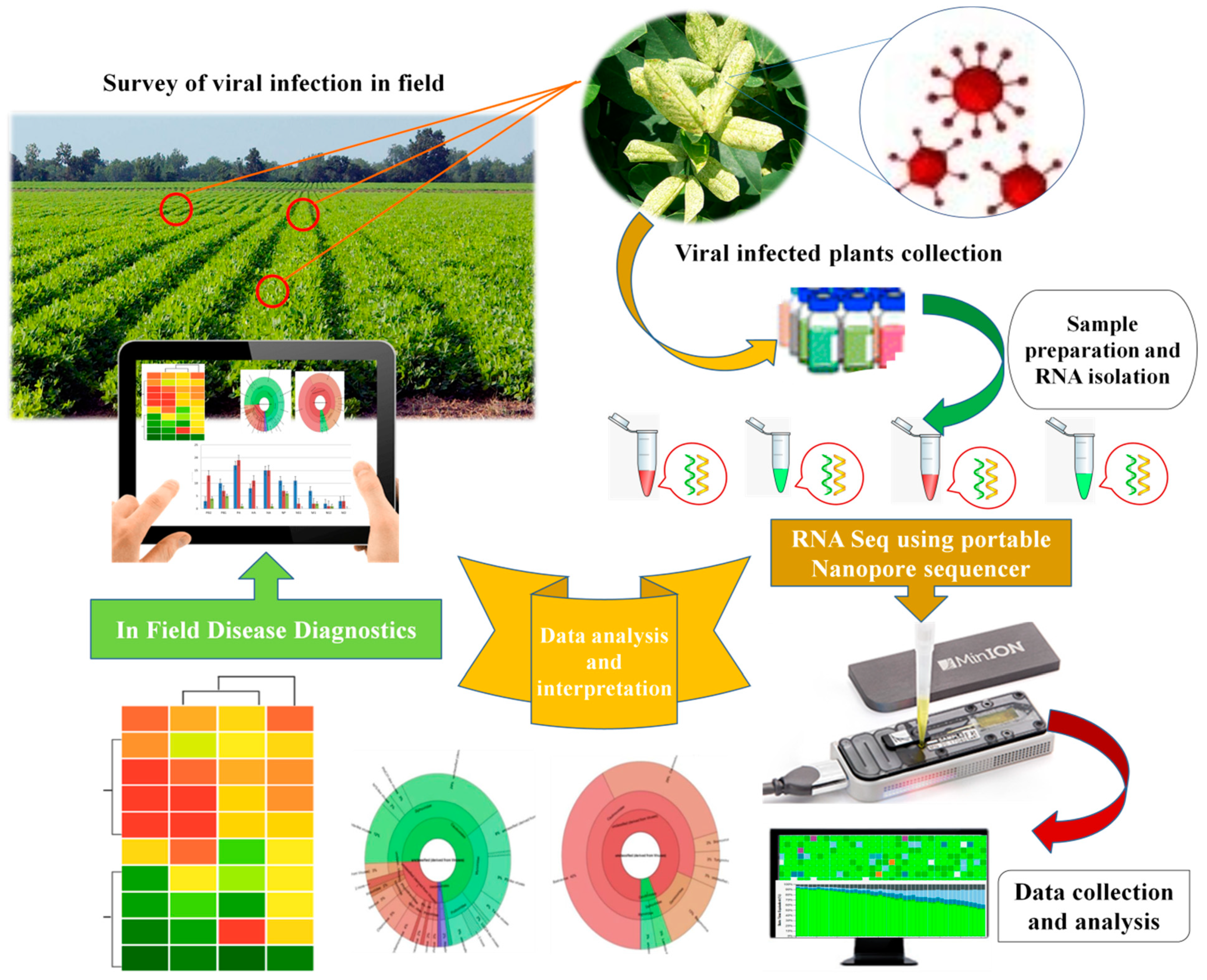
3. Current Challenges and Future Prospects (Nanopore Technology)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rojas, M.R.; Gilbertson, R.L. Emerging plant viruses: A diversity of mechanisms and opportunities. In Plant Virus Evolution; Roossinck, M.J., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 27–51. [Google Scholar]
- Maksimov, I.V.; Sorokan, A.V.; Burkhanova, G.F.; Veselova, S.V.; Alekseev, V.Y.; Shein, M.Y.; Avalbaev, A.M.; Dhaware, P.D.; Mehetre, G.T.; Singh, B.P.; et al. Mechanisms of plant tolerance to RNA viruses induced by plant-growth-promoting microorganisms. Plants 2019, 8, 575. [Google Scholar]
- Hančinský, R.; Mihálik, D.; Mrkvová, M.; Candresse, T.; Glasa, M. Plant viruses infecting Solanaceae family members in the cultivated and wild environments: A review. Plants 2020, 9, 667. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Krainer, K.M. Pandemics of people and plants: Which is the greater threat to food security? Mol. Plant 2020, 13, 933–934. [Google Scholar] [CrossRef]
- Gallet, R.; Michalakis, Y.; Blanc, S. Vector-transmission of plant viruses and constraints imposed by virus-vector interactions. Curr. Opin. Virol. 2018, 33, 144–150. [Google Scholar] [CrossRef]
- Seo, J.-K.; Kim, M.-K.; Kwak, H.-R.; Choi, H.-S.; Nam, M.; Choe, J.; Choi, B.; Han, S.-J.; Kang, J.-H.; Jung, C. Molecular dissection of distinct symptoms induced by tomato chlorosis virus and tomato yellow leaf curl virus based on comparative transcriptome analysis. Virology 2018, 516, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Adams, I.P.; Glover, R.H.; Monger, W.A.; Mumford, R.; Jackeviciene, E.; Navalinskiene, M.; Samuitiene, M.; Boonham, N. Next-generation sequencing and metagenomic analysis: A universal diagnostic tool in plant virology. Mol. Plant Pathol. 2009, 10, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Rubio, L.; Galipienso, L.; Ferriol, I. Detection of plant viruses and disease management: Relevance of genetic diversity and evolution. Front. Plant. Sci. 2020, 11, 1092. [Google Scholar] [CrossRef]
- Juárez, M.; Legua, P.; Mengual, C.M.; Kassem, M.A.; Sempere, R.N.; Gomez, P.; Truniger, V.; Aranda, M.A. Relative incidence, spatial distribution and genetic diversity of cucurbit viruses in eastern Spain. Ann. Appl. Biol. 2013, 162, 362–370. [Google Scholar] [CrossRef]
- Sanjuán, R.; Domingo-Calap, P. Mechanisms of viral mutation. Cell. Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massart, S.; Olmos, A.; Jijakli, H.; Candresse, T. Current impact and future directions of high throughput sequencing in plant virus diagnostics. Virus Res. 2014, 188, 90–96. [Google Scholar] [CrossRef]
- Mandal, B.; Jain, R.K.; Krishnareddy, M.; Krishna Kumar, N.K.; Ravi, K.S.; Pappu, H.R. Emerging problems of tospoviruses (Bunyaviridae) and their management in the Indian subcontinent. Plant Dis. 2012, 96, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Maree, H.J.; Fox, A.; Al Rwahnih, M.; Boonham, N.; Candresse, T. Application of HTS for routine plant virus diagnostics: State of the art and challenges. Front. Plant. Sci. 2018, 9, 1082. [Google Scholar] [CrossRef] [Green Version]
- Villamor, D.E.; Mekuria, T.A.; Pillai, S.S.; Eastwell, K.C. High-throughput sequencing identifies novel viruses in nectarine: Insights to the etiology of stem-pitting disease. Phytopathology 2016, 106, 519–527. [Google Scholar] [CrossRef] [Green Version]
- Stobbe, A.H.; Roossinck, M.J. Plant virus metagenomics: What we know and why we need to know more. Front. Plant Sci. 2014, 5, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malmstrom, C.M.; Melcher, U.; Bosque-Pérez, N.A. The expanding field of plant virus ecology: Historical foundations, knowledge gaps, and research directions. Virus Res. 2011, 159, 84–94. [Google Scholar] [CrossRef]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Theuns, S.; Vanmechelen, B.; Bernaert, Q.; Deboutte, W.; Vandenhole, M.; Beller, L.; Matthijnssens, J.; Maes, P.; Nauwynck, H.J. Nanopore sequencing as a revolutionary diagnostic tool for porcine viral enteric disease complexes identifies porcine kobuvirus as an important enteric virus. Sci. Rep. 2018, 8, 9830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, F.; Scalenghe, R.; Davino, S.; Panno, S.; Scuderi, G.; Ruisi, P.; Villa, P.; Stroppiana, D.; Boschetti, M.; Goulart, L.R.; et al. Advanced methods of plant disease detection. A review. Agron. Sustain. Dev. 2015, 35, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Barba, M.; Czosnek, H.; Hadidi, A. Historical perspective, development and applications of next-generation sequencing in plant virology. Viruses 2014, 6, 106–136. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahlquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Naidu, R.A.; Hughes, J.d.A. Methods for the detection of plant virus diseases. In Proceedings of the Conference on Plant Virology in Sub Saharan Africa organized by IITA, Ibadan, Nigeria, 4–8 June 2001; pp. 233–253. [Google Scholar]
- Bos, L. Symptoms of Virus Diseases in Plants, 2nd ed.; Pudoc: Wageningen, The Netherlands, 1970; p. 132. [Google Scholar]
- Webster, C.G.; Wylie, S.J.; Jones, M.G. Diagnosis of plant viral pathogens. Curr. Sci. 2004, 86, 1604–1607. [Google Scholar]
- Wu, Q.; Habili, N.; Constable, F.; Al Rwahnih, M.; Goszczynski, D.E.; Wang, Y.; Pagay, V. Virus pathogens in Australian vineyards with an emphasis on shiraz disease. Viruses 2020, 12, 818. [Google Scholar] [CrossRef] [PubMed]
- Tolkach, V.F.; Kakareka, N.N.; Volkov, Y.G.; Kozlovskaya, Z.N.; Sapotskiy, M.V.; Pleshakova, T.I.; D’yakonov, K.P.; Shchelkanov, M.Y. Virus diseases of vegetable and melon crops in the South of the Russian far East. S. Russia Ecol. Develop. 2019, 14, 121–133. [Google Scholar] [CrossRef]
- Yamaguchi, A. Increased respiration of leaves bearing necrotic local virus lesions. Virology 1960, 10, 287–293. [Google Scholar] [CrossRef]
- Senshu, H.; Yamaji, Y.; Minato, N.; Shiraishi, T.; Maejima, K.; Hashimoto, M.; Miura, C.; Neriya, Y.; Namba, S. A dual strategy for the suppression of host antiviral silencing: Two distinct suppressors for viral replication and viral movement encoded by potato virus M. J. Virol. 2011, 85, 10269–10278. [Google Scholar] [CrossRef] [Green Version]
- Balodi, R.; Bisht, S.; Ghatak, A.; Rao, K.H. Plant disease diagnosis: Technological advancements and challenges. Indian Phytopathol. 2017, 70, 275–281. [Google Scholar] [CrossRef]
- Mutka, A.M.; Bart, R.S. Image-based phenotyping of plant disease symptoms. Front. Plant Sci. 2015, 5, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, K.K.; Ramsdell, D.C.; Gillett, J. Electron microscopy: Current applications to plant virology. Plant Dis. 1985, 69, 85–91. [Google Scholar] [CrossRef]
- Horne, R.W. The development and application of electron microscopy to the structure of isolated plant viruses. In Molecular Plant Virology; eBook; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9781351074797. [Google Scholar]
- Steere, R.L.; Williams, R.C. Identification of crystalline inclusion bodies extracted intact from plant cells infected with tobacco mosaic virus. Am. J. Bot. 1953, 40, 81–84. [Google Scholar] [CrossRef]
- Kitajima, E.W. Electron microscopy in plant virology: Past, present and future. Microsc. Microanal. 2004, 10, 212–213. [Google Scholar] [CrossRef]
- Richert-Pöggeler, K.R.; Franzke, K.; Hipp, K.; Kleespies, R.G. Electron microscopy methods for virus diagnosis and high resolution analysis of viruses. Front. Microbiol. 2019, 9, 3255. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Wu, J.Y.; Xie, Y.; Liu, Y.; Qian, Y.J.; Zhou, X.P.; Wu, J.X. Monoclonal antibody-based serological assays for detection of Potato virus S in potato plants. J. Zhejiang Univ. Sci. B 2017, 18, 1075–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Wu, J.; Chen, R.; Hong, J.; Zhou, X.; Wu, J. Monoclonal antibody-based serological detection of Rice stripe mosaic virus infection in rice plants or Leafhoppers. Virol. Sin. 2020, 35, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Klap, C.; Luria, N.; Smith, E.; Bakelman, E.; Belausov, E.; Laskar, O.; Lachman, O.; Gal-On, A.; Dombrovsky, A. The potential risk of plant-virus disease initiation by infected tomatoes. Plants 2020, 9, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ye, J.; He, J.S.; Zhang, F.; Ping, J.; Qian, C.; Wu, J. Visual detection for nucleic acid-based techniques as potential on-site detection methods. A review. Anal. Chim. Acta 2020, 1099, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zechmann, B.; Zellnig, G. Rapid diagnosis of plant virus diseases by transmission electron microscopy. J. Virol. Methods 2009, 162, 163–169. [Google Scholar] [CrossRef]
- Deng, J.; Li, S.; Hong, J.; Ji, Y.; Zhou, Y. Investigation on subcellular localization of Rice stripe virus in its vector small brown planthopper by electron microscopy. Virol. J. 2013, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- El-Deeb, S.H.; IsHak, J. Histological and cytological changes due to the infection with Sweetpotato feathery mottle virus (SPFMV). J. Plant. Dis. Protect. 2004, 111, 247–256. [Google Scholar]
- El-Abhar, M.A.M.; Elkady, M.A.S.; Ghanem, K.M.; Bosila, H.A.E. Identification, characterization and ultrastructure aspects of Alfalfa mosaic virus infecting potato (Solanum tuberosum L.) in Egypt. J. Virol. Sci. 2018, 3, 68–77. [Google Scholar]
- Kozieł, E.; Otulak-Kozieł, K.; Bujarski, J.J. Analysis of Prune dwarf virus intercellular transport and pathogenesis. Int. J. Mol. Sci. 2018, 19, 2570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rong, W.; Wang, X.; Wang, X.; Massart, S.; Zhang, Z. Molecular and ultrastructural mechanisms underlying yellow dwarf symptom formation in wheat after infection of barley yellow dwarf virus. Int. J. Mol. Sci. 2018, 19, 1187. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, A.; Kulshrestha, S. Advance methods for the isolation and characterization of plant viruses infecting crops. In Applied Plant Virology; Academic Press: Cambridge, MA, USA, 2020; pp. 39–53. [Google Scholar]
- Xie, Y.; Jiao, X.; Zhou, X.; Liu, H.; Ni, Y.; Wu, J. Highly sensitive serological methods for detecting tomato yellow leaf curl virus in tomato plants and whiteflies. Virol. J. 2013, 10, 142. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Hartung, J.S. Reverse Transcription-Polymerase Chain Reaction-Based Detection of Plant Viruses. Curr. Protoc. Microbiol. 2007, 6, 16C-1. [Google Scholar] [CrossRef] [PubMed]
- Kothandaraman, S.V.; Devadason, A.; Ganesan, M.V. Seed-borne nature of a begomovirus, Mung bean yellow mosaic virus in black gram. Appl. Microbiol. Biotechnol. 2016, 100, 1925–1933. [Google Scholar] [CrossRef]
- Gorovits, R.; Czosnek, H. The involvement of heat shock proteins in the establishment of Tomato yellow leaf curl virus infection. Front. Plant Sci. 2017, 8, 355. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, P.; Budziszewska, M.; Frąckowiak, P.; Obrępalska-Stęplowska, A. Development of a new tomato Torrado virus-based vector tagged with GFP for monitoring virus movement in plants. Viruses 2020, 12, 1195. [Google Scholar] [CrossRef] [PubMed]
- Hull, R. Plant Virology, 4th ed.; Academic Press, Elsevier: Cambridge, MA, USA, 2014; ISBN 978-0-12-384871-0. [Google Scholar]
- Kang, S.H.; Sun, Y.D.; Atallah, O.O.; Huguet-Tapia, J.C.; Noble, J.D.; Folimonova, S.Y. A long non-coding RNA of Citrus tristeza virus: Role in the virus interplay with the host immunity. Viruses 2019, 11, 436. [Google Scholar] [CrossRef] [Green Version]
- Zamora, M.; Méndez-López, E.; Agirrezabala, X.; Cuesta, R.; Lavin, J.L.; Sánchez-Pina, M.A.; Aranda, M.A.; Valle, M. Potyvirus virion structure shows conserved protein fold and RNA binding site in ssRNA viruses. Sci. Adv. 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertel, K.J.; Benefield, D.; Castano-Diez, D.; Pennington, J.G.; Horswill, M.; den Boon, J.A.; Otegui, M.S.; Ahlquist, P. Cryo-electron tomography reveals novel features of a viral RNA replication compartment. eLife 2017, 6, 25940. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Atkinson, P.H.; Matthews, R.E. On the origin of dark green tissue in tobacco leaves infected with tobacco mosaic virus. Virology 1970, 40, 344–356. [Google Scholar] [CrossRef]
- Rani, A.; Donovan, N.; Mantri, N. The future of plant pathogen diagnostics in a nursery production system. Biosens. Bioelectron. 2019, 145, 111631. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, M.H. Three modern serological methods to detect plant viruses. J. Plant. Sci. Phytopathol. 2019, 3, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Ni, Y.; Liu, H.; Ding, M.; Zhou, X. Monoclonal antibody-based serological assays and immunocapture-RT-PCR for detecting Rice dwarf virus in field rice plants and leafhopper vectors. J. Virol. Methods 2014, 195, 134–140. [Google Scholar] [CrossRef]
- Nascimento, A.K.Q.; Lima, J.A.A.; Barbosa, G.D.S. A simple kit of plate-trapped antigen enzyme-linked immunosorbent assay for identification of plant viruses. Rev. Ciênc. Agron. 2017, 48, 216–220. [Google Scholar] [CrossRef]
- Seepiban, C.; Charoenvilaisiri, S.; Warin, N.; Bhunchoth, A.; Phironrit, N.; Phuangrat, B.; Chatchawankanphanich, O.; Attathom, S.; Gajanandana, O. Development and application of triple antibody sandwich enzyme-linked immunosorbent assays for begomovirus detection using monoclonal antibodies against Tomato yellow leaf curl Thailand virus. Virol. J. 2017, 14, 99. [Google Scholar] [CrossRef]
- Seal, S.; Turaki, A.; Muller, E.; Kumar, P.L.; Kenyon, L.; Filloux, D.; Galzi, S.; Lopez-Montes, A.; Iskra-Caruana, M. The prevalence of badnaviruses in West African yams (Dioscorea cayenensis-rotundata) and evidence of endogenous pararetrovirus sequences in their genomes. Virus Res. 2014, 186, 144–154. [Google Scholar] [CrossRef] [Green Version]
- Galvino-Costa, S.B.; dos Reis Figueira, A.; de Assis Câmara Rabelo-Filho, F.; Moraes, F.H.R.; Nikolaeva, O.V.; Karasev, A.V. Molecular and serological typing of Potato virus Y isolates from Brazil reveals a diverse set of recombinant strains. Plant Dis. 2012, 96, 1451–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutluk Yilmaz, N.D.; Uzunbacak, H.; Arli-Sokmen, M.; Kaya, R. Distribution of resistance-breaking isolates of beet necrotic yellow vein virus differing in virulence in sugar beet fields in Turkey. Acta Agricult. Scand. Sect. B Soil Plant Sci. 2018, 68, 546–554. [Google Scholar] [CrossRef]
- Wisler, G.C.; Lewellen, R.T.; Sears, J.L.; Liu, H.Y.; Duffus, J.E. Specificity of TAS-ELISA for beet necrotic yellow vein virus and its application for determining rhizomania resistance in field-grown sugar beets. Plant Dis. 1999, 83, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Uehara-Ichiki, T.; Shiba, T.; Matsukura, K.; Ueno, T.; Hirae, M.; Sasaya, T. Detection and diagnosis of rice-infecting viruses. Front. Microbiol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, J.A.A.; Nascimento, A.K.Q.; Silva, F.R.; Silva, A.K.F.; Aragão, M.L. An immune precipitation enzyme-linked immunosorbent (IP-ELISA) technique for identification of plant viruses. In National Meeting of Virology, Virology Meeting of Mercosur; Virus Reviews and Research: São Paulo, MO, USA, 2011; Volume 22, p. 56. [Google Scholar]
- Lima, J.A.A.; Nascimento, A.K.Q.; Radaelli, P.; Purcifull, D.E. Serology applied to plant virology. In Moslih Al-Moslih. (Org.) Serological Diagnosis of Certain Human, Animal and Plant Diseases; InTech: Rijeka, Croatia, 2012; Volume 1, pp. 71–94. [Google Scholar]
- Boonham, N.; Kreuze, J.; Winter, S.; van der Vlugt, R.; Bergervoet, J.; Tomlinson, J.; Mumford, R. Methods in virus diagnostics: From ELISA to next generation sequencing. Virus Res. 2014, 186, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Byzova, N.A.; Safenkova, I.V.; Chirkov, S.N.; Zherdev, A.V.; Blintsov, A.N.; Dzantiev, B.B.; Atabekov, I.G. Development of immunochromatographic test systems for express detection of plant viruses. Appl. Biochem. Microbiol. 2009, 45, 204–209. [Google Scholar] [CrossRef]
- Niu, Y.; Wang, D.; Cui, L.; Wang, B.; Pang, X.; Yu, P. Monoclonal antibody-based colloid gold immunochromatographic strip for the rapid detection of Tomato zonate spot tospovirus. Virol. J. 2018, 15, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bin, Y.; Li, Z.; Wu, J.; Wang, X.; Zhou, Y.; Li, T.; Song, Z. Development of an immunochromatographic strip test for rapid detection of citrus yellow vein clearing virus. Arch. Virol. 2017, 163, 349–357. [Google Scholar] [CrossRef]
- Li, R.; Mock, R.; Huang, Q.; Abad, J.; Hartung, J.; Kinard, G. A reliable and inexpensive method of nucleic acid extraction for the PCR-based detection of diverse plant pathogens. J. Virol. Methods 2008, 154, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-J.; Ju, H.J.; Noh, J. A review of detection methods for the plant viruses. Res. Plant Dis. 2014, 20, 173–181. [Google Scholar] [CrossRef]
- Mirmajlessi, S.M.; Loit, E.; Maend, M.; Mansouripour, S.M. Real-time PCR applied to study on plant pathogens: Potential applications in diagnosis-a review. Plant. Prot. Sci. 2015, 51, 177–190. [Google Scholar] [CrossRef]
- Nateqi, M.; Koohi Habibi, M.; Dizadji, A.; Parizad, S. Detection and molecular characterization of the Iris severe mosaic virus-Ir isolate from Iran. J. Plant. Protect. Res. 2015, 55, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Kimathi, R.H.; Wilisiani, F.; Mashiko, T.; Neriya, Y.; Miinda, A.E.; Nishigawa, H.; Natsuaki, T. First report of Tomato chlorosis virus infecting tomato in Kenya. Sci. Afr. 2020, 7, e00286. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Ryu, K.H.; Choi, S.H. Reverse transcription polymerase chain reaction-based system for simultaneous detection of multiple lily-infecting viruses. Plant Pathol. J. 2013, 29, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Jeevalatha, A.; Baswaraj, R.; Sharma, S.; Nagesh, M. A multiplex RT-PCR assay for simultaneous detection of five viruses in potato. J. Plant Pathol. 2017, 99, 37–45. [Google Scholar]
- Ge, B.; Li, Q.; Liu, G.; Lu, M.; Li, S.; Wang, H. Simultaneous detection and identification of four viruses infecting pepino by multiplex RT-PCR. Arch. Virol. 2013, 158, 1181–1187. [Google Scholar] [CrossRef] [PubMed]
- Mehle, N.; Gregur, L.; Bogožalec Košir, A.; Dobnik, D. One-Step Reverse-Transcription Digital PCR for Reliable Quantification of Different Pepino Mosaic Virus Genotypes. Plants 2020, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Alfaro-Fernández, A.; Sánchez-Navarro, J.Á.; del Carmen Cebrián, M.; del Carmen Córdoba-Sellés, M.; Pallás, V.; Jordá, C. Simultaneous detection and identification of Pepino mosaic virus (PepMV) isolates by multiplex one-step RT-PCR. Europ. J. Plant. Pathol. 2009, 125, 143–158. [Google Scholar] [CrossRef]
- Ling, K.S. Molecular characterization of two Pepino mosaic virus variants from imported tomato seed reveals high levels of sequence identity between Chilean and US isolates. Virus Genes 2007, 34, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Menzel, W.; Knierim, D.; Winter, S.; Hamacher, J.; Heupel, M. First report of tomato brown rugose fruit virus infecting tomato in Germany. New Dis. Rep. 2019, 39. [Google Scholar] [CrossRef] [Green Version]
- Ling, K.S.; Tian, T.; Gurung, S.; Salati, R.; Gilliard, A. First report of tomato brown rugose fruit virus infecting greenhouse tomato in the United States. Plant Dis. 2019, 103, 1439. [Google Scholar] [CrossRef]
- Kumar, S.; Udaya Shankar, A.C.; Nayaka, S.C.; Lund, O.S.; Prakash, H.S. Detection of Tobacco mosaic virus and Tomato mosaic virus in pepper and tomato by multiplex RT-PCR. Lett. Appl. Microbiol. 2011, 53, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Cherian, S.; Joseph, J.; Muniyappa, V.; Savithri, H.S. Characterization of tobacco mosaic virus isolated from tomato in India. Curr. Sci. 1999, 14, 65–69. [Google Scholar]
- Nagendran, K.; Mohankumar, S.; Aravintharaj, R.; Balaji, C.G.; Manoranjitham, S.K.; Singh, A.K.; Rai, A.B.; Singh, B.; Karthikeyan, G. The occurrence and distribution of major viruses infecting cucurbits in Tamil Nadu state, India. Crop. Prot. 2017, 99, 10–16. [Google Scholar] [CrossRef]
- Vinayarani, G.; Madhusudhan, K.N.; Deepak, S.A.; Niranjana, S.R.; Prakash, H.S. Detection of mixed infection of tobamoviruses in tomato and bell pepper by using RT-PCR and duplex RT-PCR. Int. J. Plant. Pathol. 2011, 2, 89–95. [Google Scholar] [CrossRef]
- Deng, D.; McGrath, P.F.; Robinson, D.J.; Harrison, B.D. Detection and differentiation of whitefly-transmitted geminiviruses in plants and vector insects by the polymerase chain reaction with degenerate primers. Ann. Appl. Biol. 1994, 125, 327–336. [Google Scholar] [CrossRef]
- Nagendran, K.; Kumar, S.M.; Manoranjitham, S.K.; Karthikeyan, G. Molecular detection and characterization of Tomato leaf curl New Delhi virus causing mosaic disease on bitter gourd in Tamil Nadu, India. Trends Biosci. 2014, 7, 3925–3931. [Google Scholar]
- Hyun, J.W.; Hwang, R.Y.; Jung, K.E. Development of multiplex PCR for simultaneous detection of citrus viruses and the incidence of citrus viral diseases in late-maturity citrus trees in Jeju Island. Plant Pathol. J. 2017, 33, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagendran, K.; Pandey, K.K.; Rai, A.B.; Singh, B. Viruses of Vegetable Crops: Symptomatology, Diagnostics and Management; IIVR Technical Bulletin No. 75; IIVR—Indian Institute of Vegetable Research: Varanasi, India, 2017; p. 48. [Google Scholar]
- Sinha, S.; Samad, A. First report of Cucumber mosaic virus associated with yellowing mosaic disease of African basil (Ocimum gratissimum) in India. Plant Dis. 2019, 103, 167. [Google Scholar] [CrossRef]
- Lu, Y.; Yao, B.; Wang, G.; Hong, N. The detection of ACLSV and ASPV in pear plants by RT-LAMP assays. J. Virol. Methods 2018, 252, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wu, K.; Wu, W.; Mi, W.; Hao, X.; Wu, Y. A multiplex reverse transcription PCR assay for simultaneous detection of six main RNA viruses in tomato plants. J. Virol. Methods 2019, 265, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tan, G.; Lan, P.; Zhang, A.; Liu, Y.; Li, R.; Li, F. Detection of tobamoviruses by RT-PCR using a novel pair of degenerate primers. J. Virol. Methods 2018, 259, 122–128. [Google Scholar] [CrossRef]
- Majumder, S.; and Baranwal, V.K. Simultaneous detection of four garlic viruses by multiplex reverse transcription PCR and their distribution in Indian garlic accessions. J. Virol. Methods 2014, 202, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Van Brunschot, S.L.; Bergervoet, J.H.W.; Pagendam, D.E.; de Weerdt, M.; Geering, A.D.W.; Drenth, A.; van der Vlugt, R.A. Development of a multiplexed bead-based suspension array for the detection and discrimination of Pospiviroid plant pathogens. PLoS ONE 2014, 9, e84743. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Rodoni, B.C.; Gibbs, M.J.; Gibbs, A.J. A novel pair of universal primers for the detection of potyviruses. Plant. Pathol. 2009, 59, 211–220. [Google Scholar] [CrossRef]
- Pallás, V.; Sánchez-Navarro, J.A.; James, D. Recent advances on the multiplex molecular detection of plant viruses and viroids. Front. Microbiol. 2018, 9, 2087. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, V. Immunocapture-PCR for plant virus detection. In Plant Pathology; Humana Press: Totowa, NJ, USA, 2009; pp. 183–192. [Google Scholar]
- Koolivand, D.; Sokhandan-Bashir, N.; Behjatnia, S.A.A.; Jafari Joozani, R.A. Detection of Grapevine fanleaf virus by immunocapture reverse transcription-polymerase chain reaction (IC-RT-PCR) with recombinant antibody. Arch. Phytopathol. Plant. Prot. 2014, 47, 2070–2077. [Google Scholar] [CrossRef]
- Vigne, E.; Garcia, S.; Komar, V.; Lemaire, O.; Hily, J.M. Comparison of serological and molecular methods with high-throughput sequencing for the detection and quantification of Grapevine Fanleaf Virus in vineyard samples. Front. Microbiol. 2018, 9, 2726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, E63. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.M.; Li, G.; Gao, Y.; Zhu, Y.R.; Liu, J.; Zhu, X.P. Reverse transcription loop-mediated isothermal amplification assay for detecting tomato chlorosis virus. J. Virol. Methods 2015, 213, 93–97. [Google Scholar] [CrossRef]
- Bhat, A.I.; Siljo, A.; Deeshma, K.P. Rapid detection of Piper yellow mottle virus and Cucumber mosaic virus infecting black pepper (Piper nigrum) by loop-mediated isothermal amplification (LAMP). J. Virol. Methods 2013, 193, 190–196. [Google Scholar] [CrossRef]
- Ahmadi, S.; Almasi, M.A.; Fatehi, F.; Struik, P.C.; Moradi, A. Visual detection of Potato Leafroll Virus by one-step reverse transcription loop-mediated isothermal amplification of DNA with hydroxynaphthol blue dye. J. Phytopathol. 2013, 161, 120–124. [Google Scholar] [CrossRef]
- Elvira-González, L.; Puchades, A.V.; Carpino, C.; Alfaro-Fernandez, A.; Font-San-Ambrosio, M.I.; Rubio, L.; Galipienso, L. Fast detection of Southern tomato virus by one-step transcription loop-mediated isothermal amplification (RT-LAMP). J. Virol. Methods 2017, 241, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Sarkes, A.; Fu, H.; Feindel, D.; Harding, M.; Feng, J. Development and evaluation of a loop-mediated isothermal amplification (LAMP) assay for the detection of Tomato brown rugose fruit virus (ToBRFV). PLoS ONE 2020, 15, e0230403. [Google Scholar] [CrossRef]
- Peng, Q.; Ning, J.; Xu, Q.; Yang, T.; Wang, Y.; Zheng, T.; Zhuang, Q.; Xi, D. Development and application of a reverse transcription loop-mediated isothermal amplification combined with lateral flow dipstick for rapid and visual detection of Citrus leaf blotch virus in kiwifruit. Crop. Protect. 2021, 143, 105555. [Google Scholar] [CrossRef]
- Kokane, A.; Kokane, S.; Warghane, A.; Gubyad, M.G.; Sharma, A.K.; Reddy, K.M.; Ghosh, D.K. A Rapid and Sensitive Reverse Transcription-Loop-Mediated Isothermal Amplification (RT-LAMP) Assay for the Detection of Indian Citrus Ringspot Virus. Plant Dis. 2020. [Google Scholar] [CrossRef]
- Warghane, A.; Misra, P.; Bhose, S.; Biswas, K.K.; Sharma, A.K.; Reddy, M.K.; Ghosh, D.K. Development of a simple and rapid reverse transcription-loop mediated isothermal amplification (RT-LAMP) assay for sensitive detection of Citrus tristeza virus. J. Virol. Methods 2017, 250, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Qiao, N.; Dai, H.; Liu, J.; Zhu, X.; Li, J.; Zhang, D.; Liu, Y. Detection of melon necrotic spot virus by one-step reverse transcription loop-mediated isothermal amplification assay. PLoS ONE 2020, 15, e0230023. [Google Scholar]
- Wilisiani, F.; Tomiyama, A.; Katoh, H.; Hartono, S.; Neriya, Y.; Nishigawa, H.; Natsuaki, T. Development of a LAMP assay with a portable device for real-time detection of begomoviruses under field conditions. J. Virol. Methods 2019, 265, 71–76. [Google Scholar] [CrossRef]
- Selvaraj, V.; Maheshwari, Y.; Hajeri, S.; Yokomi, R. A rapid detection tool for VT isolates of Citrus tristeza virus by immunocapture-reverse transcriptase loop-mediated isothermal amplification assay. PLoS ONE 2019, 14, e0222170. [Google Scholar] [CrossRef] [Green Version]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, S. Loop Mediated Isothermal Amplification: Principles and Applications in Plant Virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Dumschott, K.; Schmidt, M.H.; Chawla, H.S.; Snowdon, R.; Usadel, B. Oxford Nanopore Sequencing: New opportunities for plant genomics? J. Exp. Bot. 2020, 71, 5313–5322. [Google Scholar] [CrossRef] [PubMed]
- Mehetre, G.T.; Paranjpe, A.S.; Dastager, S.G.; Dharne, M.S. Complete metagenome sequencing based bacterial diversity and functional insights from basaltic hot spring of Unkeshwar, Maharashtra, India. Genomics Data 2016, 7, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, A. Next-generation sequencing and CRISPR/Cas13 editing in viroid research and molecular diagnostics. Viruses 2019, 11, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riesenfeld, C.S.; Schloss, P.D.; Handelsman, J. Metagenomics: Genomic analysis of microbial communities. Annu. Rev. Genet. 2004, 38, 525–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blawid, R.; Silva, J.M.; Nagata, T. Discovering and sequencing new plant viral genomes by next-generation sequencing: Description of a practical pipeline. Ann. Appl. Biol. 2017, 170, 301–314. [Google Scholar] [CrossRef]
- Xiao, H.; Li, C.; Al Rwahnih, M.; Dolja, V.; Meng, B. Metagenomic analysis of Riesling grapevine reveals a complex virome including two new and divergent variants of grapevine leafroll-associated virus 3. Plant Dis. 2019, 103, 1275–1285. [Google Scholar] [CrossRef]
- Nie, X.; Singh, R.P. Detection of multiple potato viruses using an oligo (dT) as a common cDNA primer in multiplex RT-PCR. J. Virol. Methods 2000, 86, 179–185. [Google Scholar] [CrossRef]
- Pooggin, M.M. Small RNA-omics for plant virus identification, virome reconstruction, and antiviral defense characterization. Front. Microbiol. 2018, 9, 2779. [Google Scholar] [CrossRef]
- Ambardar, S.; Gupta, R.; Trakroo, D.; Lal, R.; Vakhlu, J. High throughput sequencing: An overview of sequencing chemistry. Ind. J. Microbiol. 2016, 56, 394–404. [Google Scholar] [CrossRef] [Green Version]
- Lipkin, W.I.; Firth, C. Viral surveillance and discovery. Curr. Opin. Virol. 2013, 3, 199–204. [Google Scholar] [CrossRef] [Green Version]
- Barzon, L.; Lavezzo, E.; Costanzi, G.; Franchin, E.; Toppo, S.; Palù, G. Next-generation sequencing technologies in diagnostic virology. J. Clin. Virol. 2013, 58, 346–350. [Google Scholar] [CrossRef]
- Kreuze, J.F.; Perez, A.; Untiveros, M.; Quispe, D.; Fuentes, S.; Barker, I.; Simon, R. Complete viral genome sequence and discovery of novel viruses by deep sequencing of small RNAs: A generic method for diagnosis, discovery and sequencing of viruses. Virology 2009, 388, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Xing, F.; Gao, D.; Liu, H.; Wang, H.; Habili, N.; Li, S. Molecular characterization and pathogenicity analysis of prunus necrotic ringspot virus isolates from China rose (Rosa chinensis Jacq.). Arch. Virol. 2020, 165, 2479–2486. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. RNA silencing as a plant immune system against viruses. Trends Genet. 2001, 17, 449–459. [Google Scholar] [CrossRef]
- Maliogka, V.I.; Minafra, A.; Saldarelli, P.; Ruiz-García, A.B.; Glasa, M.; Katis, N.; Olmos, A. Recent advances on detection and characterization of fruit tree viruses using high-throughput sequencing technologies. Viruses 2018, 10, 436. [Google Scholar] [CrossRef] [Green Version]
- Loconsole, G.; Saldarelli, P.; Doddapaneni, H.; Savino, V.; Martelli, G.P.; Saponari, M. Identification of a single-stranded DNA virus associated with citrus chlorotic dwarf disease, a new member in the family Geminiviridae. Virology 2012, 432, 162–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.J.; Wang, J.; Todd, A.K.; Bissielo, A.B.; Yen, S.; Strydom, H.; Moore, N.E.; Ren, X.; Huang, Q.S.; Carter, P.E.; et al. Evaluation of rapid and simple techniques for the enrichment of viruses prior to metagenomic virus discovery. J. Virol. Methods 2014, 195, 194–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quiñones-Mateu, M.E.; Avila, S.; Reyes-Teran, G.; Martinez, M.A. Deep sequencing: Becoming a critical tool in clinical virology. J. Clin. Virol. 2014, 61, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Mumo, N.N.; Mamati, G.E.; Ateka, E.M.; Rimberia, F.K.; Asudi, G.O.; Boykin, L.M.; Machuka, E.M.; Njuguna, J.N.; Pelle, R.; Stomeo, F. Metagenomic analysis of plant viruses associated with Papaya Ringspot disease in Carica papaya L. in Kenya. Front. Microbiol. 2020, 11, 205. [Google Scholar] [CrossRef]
- Nhlapo, T.F.; Rees, D.J.; Odeny, D.A.; Mulabisana, J.M.; Rey, M.E. Viral metagenomics reveals sweet potato virus diversity in the Eastern and Western Cape provinces of South Africa. S. Afr. J. Bot. 2018, 117, 256–267. [Google Scholar] [CrossRef]
- Palanga, E.; Filloux, D.; Martin, D.P.; Fernandez, E.; Gargani, D.; Ferdinand, R.; Zabré, J.; Bouda, Z.; Neya, J.B.; Sawadogo, M.; et al. Metagenomic-based screening and molecular characterization of cowpea-infecting viruses in Burkina Faso. PLoS ONE 2016, 11, e0165188. [Google Scholar] [CrossRef] [Green Version]
- De Nazaré Almeida dos Reis, L.; Fonseca, M.E.; Ribeiro, S.G.; Naito, F.Y.; Boiteux, L.S.; Pereira-Carvalho, R.D. Metagenomics of Neotropical single-stranded DNA viruses in tomato cultivars with and without the Ty-1 gene. Viruses 2020, 12, 819. [Google Scholar] [CrossRef]
- Wu, Q.; Kehoe, M.A.; Kinoti, W.M.; Wang, C.P.; Rinaldo, A.; Tyerman, S.; Habili, N.; Constable, F.E. First report of grapevine rupestris vein feathering virus in grapevine in Australia. Plant Dis. 2020. [Google Scholar] [CrossRef]
- Ma, Y.; Marais, A.; Lefebvre, M.; Theil, S.; Svanella-Dumas, L.; Faure, C.; Candresse, T. Phytovirome analysis of wild plant populations: Comparison of double-stranded RNA and virion-associated nucleic acid metagenomic approaches. J. Virol. 2019, 94, e01462-19. [Google Scholar] [CrossRef] [PubMed]
- Wainaina, J.M.; Ateka, E.; Makori, T.; Kehoe, M.A.; Boykin, L.M. A metagenomic study of DNA viruses from samples of local varieties of common bean in Kenya. Peer J. 2019, 7, e6465. [Google Scholar] [CrossRef] [Green Version]
- Rott, M.; Xiang, Y.; Boyes, I.; Belton, M.; Saeed, H.; Kesanakurti, P.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B.; et al. Application of next generation sequencing for diagnostic testing of tree fruit viruses and viroids. Plant Dis. 2017, 101, 1489–1499. [Google Scholar] [CrossRef] [Green Version]
- Maina, S.; Edwards, O.R.; de Almeida, L.; Ximenes, A.; Jones, R.A. Metagenomic analysis of cucumber RNA from East Timor reveals an Aphid lethal paralysis virus genome. Genome Announce 2017, 5, e01445-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecman, A.; Kutnjak, D.; Gutiérrez-Aguirre, I.; Adams, I.; Fox, A.; Boonham, N.; Ravnikar, M. Next generation sequencing for detection and discovery of plant viruses and viroids: Comparison of two approaches. Front. Microbiol. 2017, 8, 1998. [Google Scholar] [CrossRef] [Green Version]
- Mutuku, J.M.; Wamonje, F.O.; Mukeshimana, G.; Njuguna, J.; Wamalwa, M.; Choi, S.K.; Tungadim, T.; Djikeng, A.; Kelly, K.; Domelevo Entfellner, J.B.; et al. Metagenomic analysis of plant virus occurrence in common bean (Phaseolus vulgaris) in Central Kenya. Front. Microbiol. 2018, 9, 2939. [Google Scholar] [CrossRef] [PubMed]
- Baráth, D.; Jaksa-Czotter, N.; Molnár, J.; Varga, T.; Balássy, J.; Szabó, L.K.; Kirilla, Z.; Tusnády, G.E.; Preininger, É.; Várallyay, É. Small RNA NGS revealed the presence of Cherry virus A and Little cherry virus 1 on apricots in Hungary. Viruses 2018, 10, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinoti, W.M.; Nancarrow, N.; Dann, A.; Rodoni, B.C.; Constable, F.E. Updating the quarantine status of prunus infecting viruses in Australia. Viruses 2020, 12, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsiani, A.; Maliogka, V.I.; Katis, N.; Svanella-Dumas, L.; Olmos, A.; Ruiz-García, A.B.; Marais, A.; Faure, C.; Theil, S.; Lotos, L.; et al. High-throughput sequencing reveals further diversity of Little cherry virus 1 with implications for diagnostics. Viruses 2018, 10, 385. [Google Scholar] [CrossRef] [Green Version]
- Wamaitha, M.J.; Nigam, D.; Maina, S.; Stomeo, F.; Wangai, A.; Njuguna, J.N.; Holton, T.A.; Wanjala, B.W.; Wamalwa, M.; Lucas, T.; et al. Metagenomic analysis of viruses associated with maize lethal necrosis in Kenya. Virol. J. 2018, 15, 90. [Google Scholar] [CrossRef]
- Rott, M.E.; Kesanakurti, P.; Berwarth, C.; Rast, H.; Boyes, I.; Phelan, J.; Jelkmann, W. Discovery of negative-sense RNA viruses in trees infected with apple rubbery wood disease by next-generation sequencing. Plant Dis. 2018, 102, 1254–1263. [Google Scholar] [CrossRef] [Green Version]
- Bartsch, C.; Höper, D.; Mäde, D.; Johne, R. Analysis of frozen strawberries involved in a large norovirus gastroenteritis outbreak using next generation sequencing and digital PCR. Food Microbiol. 2018, 76, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, S.; Na, C.; Yang, L.; Lu, M. Analyses of virus/viroid communities in nectarine trees by next-generation sequencing and insight into viral synergisms implication in host disease symptoms. Sci. Rep. 2019, 9, 12261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akinyemi, I.A.; Wang, F.; Zhou, B.; Qi, S.; Wu, Q. Ecogenomic survey of plant viruses infecting tobacco by next generation sequencing. Virol. J. 2016, 13, 181. [Google Scholar] [CrossRef] [Green Version]
- Kinoti, W.M.; Constable, F.E.; Nancarrow, N.; Plummer, K.M.; Rodoni, B. Analysis of intra-host genetic diversity of Prunus necrotic ringspot virus (PNRSV) using amplicon next generation sequencing. PLoS ONE 2017, 12, e0179284. [Google Scholar] [CrossRef] [Green Version]
- Kinoti, W.M.; Constable, F.E.; Nancarrow, N.; Plummer, K.M.; Rodoni, B. Generic amplicon deep sequencing to determine Ilarvirus species diversity in Australian Prunus. Front. Microbiol. 2017, 8, 1219. [Google Scholar] [CrossRef]
- Díaz-Cruz, G.A.; Smith, C.M.; Wiebe, K.F.; Villanueva, S.M.; Klonowski, A.R.; Cassone, B.J. Applications of Next-generation sequencing for large-scale pathogen diagnoses in Soybean. Plant Dis. 2019, 103, 1075–1083. [Google Scholar] [CrossRef] [PubMed]
- Fajardo, T.V.; Silva, F.N.; Eiras, M.; Nickel, O. High-throughput sequencing applied for the identification of viruses infecting grapevines in Brazil and genetic variability analysis. Trop. Plant Pathol. 2017, 42, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Eichmeier, A.; Komínková, M.; Komínek, P.; Baránek, M. Comprehensive virus detection using next generation sequencing in grapevine vascular tissues of plants obtained from the wine regions of Bohemia and Moravia (Czech Republic). PLoS ONE 2016, 11, e0167966. [Google Scholar] [CrossRef] [PubMed]
- Grisoni, M.; Marais, A.; Filloux, D.; Saison, A.; Faure, C.; Julian, C.; Theil, S.; Contreras, S.; Teycheney, P.Y.; Roumagnac, P.; et al. Two novel Alphaflexiviridae members revealed by deep sequencing of the Vanilla (Orchidaceae) virome. Arch. Virol. 2017, 162, 3855–3861. [Google Scholar] [CrossRef] [PubMed]
- Greninger, A.L. A decade of RNA virus metagenomics is (not) enough. Virus Res. 2018, 244, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Luria, N.; Smith, E.; Reingold, V.; Bekelman, I.; Lapidot, M.; Levin, I.; Elad, N.; Tam, Y.; Sela, N.; Abu-Ras, A.; et al. A new Israeli Tobamovirus isolate infects tomato plants harboring Tm-22 resistance genes. PLoS ONE 2017, 12, e0170429. [Google Scholar] [CrossRef] [Green Version]
- Edwards, D.; Batley, J. Plant genome sequencing: Applications for crop improvement. Plant Biotech. J. 2010, 8, 2–9. [Google Scholar] [CrossRef]
- Lavezzo, E.; Barzon, L.; Toppo, S.; Palu, G. Third generation sequencing technologies applied to diagnostic microbiology: Benefits and challenges in applications and data analysis. Expert Rev. Mol. Diag. 2016, 16, 1011–1023. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Martin, I.; Moschetti, W.E.; Kershaw, C.M.; Tsongalis, G.J. Third-generation sequencing in the clinical laboratory: Exploring the advantages and challenges of nanopore sequencing. J. Clin. Microbiol. 2019, 58, e01315-19. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, A.; Au, K.F. PacBio sequencing and its applications. Genom. Proteom. Bioinf. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Giordano, F.; Ning, Z. Oxford Nanopore MinION sequencing and genome assembly. Genom. Proteom. Bioinf. 2016, 14, 265–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leigh, D.M.; Schefer, C.; Cornejo, C. Determining the suitability of MinION’s direct RNA and DNA amplicon sequencing for viral subtype identification. Viruses 2020, 12, 801. [Google Scholar] [CrossRef]
- Tyler, A.D.; Mataseje, L.; Urfano, C.J.; Schmidt, L.; Antonation, K.S.; Mulvey, M.R.; Corbett, C.R. Evaluation of Oxford Nanopore’s MinION sequencing device for microbial whole genome sequencing applications. Sci. Rep. 2018, 8, 10931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garalde, D.R.; Snell, E.A.; Jachimowicz, D.; Sipos, B.; Lloyd, J.H.; Bruce, M.; Pantic, N.; Admassu, T.; James, P.; Warland, A.; et al. Highly parallel direct RNA sequencing on an array of nanopores. Nat. Methods. 2018, 15, 201. [Google Scholar] [CrossRef]
- Krehenwinkel, H.; Pomerantz, A.; Prost, S. Genetic biomonitoring and biodiversity assessment using portable sequencing technologies: Current uses and future directions. Genes 2019, 10, 858. [Google Scholar] [CrossRef] [Green Version]
- Mushtaq, Z.; Prasad, K.P.; Qayoom, U. Nanopore Sequencing for diagnosis and resistance profiling of pathogens. Biotica Res. Today 2020, 2, 908–911. [Google Scholar]
- Latorre-Pérez, A.; Pascual, J.; Porcar, M.; Vilanova, C. A lab in the field: Applications of real-time, in situ metagenomic sequencing. Biol. Methods Protoc. 2020, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, J.; Runtuwene, L.R.; Hayashida, K.; Mongan, A.E.; Thi, L.A.N.; Thuy, L.N.; Nhat, C.N.; Limkittikul, K.; Sirivichayakul, C.; Sathirapongsasuti, N. Serotyping dengue virus with isothermal amplification and a portable sequencer. Sci. Rep. 2017, 7, 3510. [Google Scholar] [CrossRef]
- Batovska, J.; Lynch, S.E.; Rodoni, B.C.; Sawbridge, T.I.; Cogan, N.O.I. Metagenomic arbovirus detection using MinION nanopore sequencing. J. Virol. Methods 2017, 249, 79–84. [Google Scholar] [CrossRef]
- Quick, J.; Grubaugh, N.D.; Pullan, S.T.; Claro, I.M.; Smith, A.D.; Gangavarapu, K.; Oliveira, G.; Robles-Sikisaka, R.; Rogers, T.F.; Beutler, N.A.; et al. Multiplex PCR method for MinION and Illumina sequencing of Zika and other virus genomes directly from clinical samples. Nat. Protoc. 2017, 12, 1261–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Della Bartola, M.; Byrne, S.; Mullins, E. Characterization of potato virus Y isolates and assessment of nanopore sequencing to detect and genotype potato viruses. Viruses 2020, 12, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monger, W.A.; Goodfellow, H.A.; Back, E.J. First report of Arabis mosaic virus in potato (Solanum tuberosum), identified by nanopore sequencing. New Dis. Rep. 2020, 41, 29. [Google Scholar] [CrossRef]
- Leiva, A.M.; Siriwan, W.; Lopez-Alvarez, D.; Barrantes, I.; Hemniam, N.; Saokham, K.; Cuellar, W.J. Nanopore-based complete genome sequence of a Sri Lankan cassava mosaic virus (Geminivirus) strain from Thailand. Microbiol. Resour. Announce 2020, 9, e01274-19. [Google Scholar] [CrossRef] [Green Version]
- Stenger, D.C.; Burbank, L.P.; Wang, R.; Stewart, A.A.; Mathias, C.; Goodin, M.M. Lost and found: Rediscovery and genomic characterization of sowthistle yellow vein virus after a 30+ year hiatus. Virus Res. 2020, 284, 197987. [Google Scholar] [CrossRef]
- Duarte, M.F.; Fonseca, M.E.; Boiteux, L.S.; Costa, H.; Ribeiro, B.M.; Melo, F.L.; Pereira-Carvalho, R.C. Identification of Physalis angulata (Solanaceae) as a natural alternative weed host of tomato severe rugose virus in Brazil. Plant Dis. 2020, 104, 600. [Google Scholar] [CrossRef]
- Naito, F.Y.; Melo, F.L.; Fonseca, M.E.; Santos, C.A.; Chanes, C.R.; Ribeiro, B.M.; Gilbertson, R.L.; Boiteux, L.S.; de Cássia Pereira-Carvalho, R. Nanopore sequencing of a novel bipartite new world begomovirus infecting cowpea. Arch. Virol. 2019, 164, 1907–1910. [Google Scholar] [CrossRef]
- Fellers, J.P.; Webb, C.; Fellers, M.C.; Shoup Rupp, J.; De Wolf, E. Wheat virus identification within infected tissue using nanopore sequencing technology. Plant Dis. 2019, 103, 2199–2203. [Google Scholar] [CrossRef]
- Boykin, L.; Ghalab, A.; De Marchi, B.R.; Savill, A.; Wainaina, J.M.; Kinene, T.; Lamb, S.; Rodrigues, M.; Kehoe, M.; Ndunguru, J.; et al. Real time portable genome sequencing for global food security. F1000Res 2018, 7, 1101. [Google Scholar] [CrossRef] [Green Version]
- Chalupowicz, L.; Dombrovsky, A.; Gaba, V.; Luria, N.; Reuven, M.; Beerman, A.; Lachman, O.; Dror, O.; Nissan, G.; Manulis-Sasson, S. Diagnosis of plant diseases using the nanopore sequencing platform. Plant. Pathol. 2019, 68, 229–238. [Google Scholar] [CrossRef]
- Filloux, D.; Fernandez, E.; Loire, E.; Claude, L.; Galzi, S.; Candresse, T.; Winter, S.; Jeeva, M.L.; Makeshkumar, T.; Martin, D.P.; et al. Nanopore-based detection and characterization of yam viruses. Sci. Rep. 2018, 8, 17879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronzato Badial, A.; Sherman, D.; Stone, A.; Gopakumar, A.; Wilson, V.; Schneider, W.; King, J. Nanopore sequencing as a surveillance tool for plant pathogens in plant and insect tissues. Plant Dis. 2018, 102, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Barreto, S.S.; Hallwass, M.; Aquino, O.M.; Inoue-Nagata, A.K. A study of weeds as potential inoculum sources for a tomato-infecting begomovirus in central Brazil. Phytopathology 2013, 103, 436–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Suarez, C.J.; Banaei, N.; Pinsky, B.A. Next-generation sequencing for infectious disease diagnosis and management: A report of the Association for Molecular Pathology. J. Mol. Diagn. 2015, 17, 623–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; Ramasamy, R.P. Current and prospective methods for plant disease detection. Biosensors 2015, 5, 537–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leggett, R.M.; Clark, M.D. A world of opportunities with nanopore sequencing. J. Exp. Bot. 2017, 68, 5419–5429. [Google Scholar] [CrossRef]
- Midha, M.K.; Wu, M.; Chiu, K.P. Long-read sequencing in deciphering human genetics to a greater depth. Hum. Genet. 2019, 138, 1201–1215. [Google Scholar] [CrossRef] [PubMed]
- Shabardina, V.; Kischka, T.; Manske, F.; Grundmann, N.; Frith, M.C.; Suzuki, Y.; Makałowski, W. NanoPipe-a web server for nanopore MinION sequencing data analysis. Gigascience 2019, 8, giy169. [Google Scholar] [CrossRef] [Green Version]
- Nezhad, A.S. Future of portable devices for plant pathogen diagnosis. Lab Chip 2014, 14, 2887–2904. [Google Scholar] [CrossRef] [PubMed]
- Mokili, J.L.; Rohwer, F.; Dutilh, B.E. Metagenomics and future perspectives in virus discovery. Curr. Opin. Virol. 2012, 2, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Li, J.; Chu, X.; Liu, H.; Liu, W.; Liu, D. Nanopore sequencing: A rapid solution for infectious disease epidemics. Sci. China Life Sci. 2019, 62, 1101–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depledge, D.P.; Srinivas, K.P.; Sadaoka, T.; Bready, D.; Mori, Y.; Placantonakis, D.G.; Mohr, I.; Wilson, A.C. Direct RNA sequencing on nanopore arrays redefines the transcriptional complexity of a viral pathogen. Nat. Commun. 2019, 10, 754. [Google Scholar] [CrossRef] [Green Version]
- Rang, F.J.; Kloosterman, W.P.; de Ridder, J. From squiggle to basepair: Computational approaches for improving nanopore sequencing read accuracy. Genome Biol. 2018, 19, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Faria, J.C.; Aragã, F.J.; Souza, T.L.; Quintela, E.D.; Kitajima, E.W.; Ribeiro, S.D. Golden mosaic of common beans in Brazil: Management with a transgenic approach. APS Features 2016. [Google Scholar] [CrossRef]
- Hoenen, T.; Groseth, A.; Rosenke, K.; Fischer, R.J.; Hoenen, A.; Judson, S.D.; Martellaro, C.; Falzarano, D.; Marzi, A.; Squires, R.B.; et al. Nanopore sequencing as a rapidly deployable Ebola outbreak tool. Emerg. Infect. Dis. 2016, 22, 331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilianski, A.; Roth, P.A.; Liem, A.T.; Hill, J.M.; Willis, K.L.; Rossmaier, R.D.; Marinich, A.V.; Maughan, M.N.; Karavis, M.A.; Kuhn, J.H.; et al. Use of unamplified RNA/cDNA-hybrid nanopore sequencing for rapid detection and characterization of RNA viruses. Emerg. Infect. Dis. 2016, 22, 1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boykin, L.M.; Sseruwagi, P.; Alicai, T.; Ateka, E.; Mohammed, I.U.; Stanton, J.A.; Kayuki, C.; Mark, D.; Fute, T.; Erasto, J.; et al. Tree Lab: Portable genomics for early detection of plant viruses and pests in Sub-Saharan Africa. Genes 2019, 10, 632. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Virus Name | Host Plant | ELISA Based Method | Reference |
|---|---|---|---|
| Rice stripe mosaic virus (RSMV) | Rice | ACP-ELISA (antigen-coated plate enzyme-linked immunosorbent assay) | [37] |
| Dot-ELISA | |||
| Tissue print-ELISA | |||
| Rice dwarf virus (RDV) | Rice | PTA-ELISA (Plate-trapped antigen enzyme-linked immunosorbent assay) | [60] |
| Dot-ELISA | |||
| Squash mosaic virus (SQMV), Cowpea severe mosaic virus (CPSMV), Cucumber mosaic virus (CMV), Cowpea aphid borne mosaic virus (CABMV), Papaya lethal yellowing virus (PLYV) | Squash Cowpea Papaya | PTA-ELISA | [61] |
| Tomato yellow leaf curl Thailand virus (TYLCTHV) | Tomato, Pepper, Eggplant, Okra, Cucurbit | TAS-ELISA (triple antibody sandwich enzyme-linked immunosorbent assays) | [62] |
| Beet soil-borne virus (BSBV) | sugar beet | TAS-ELISA | [65] |
| Tomato yellow leaf curl Thailand virus, TYLCTHV | Tomato | TAS-ELISA | [62] |
| beet necrotic yellow vein virus (BNYVV) | sugar beet | TAS-ELISA | [66] |
| Potato virus Y (PVY) | Potato | TAS-ELISA | [64] |
| Potato virus S (PVS) | Potato | DAS-ELISA (double-antibody sandwich ELISA) direct ELISA | [36] |
| Tissue print-ELISA | |||
| Dot-ELISA | |||
| Rice-infecting viruses | Rice | DAS-ELISA | [67] |
| Badnaviruses | West African Yam | PAS-ELISA (Protein A-Sandwich ELISA) | [63] |
| Comovirus, Cowpea severe mosaic virus (CPSMV), Squash mosaic virus (SQMV) | Cowpea, Squash | IP-ELISA (Immune Precipitation ELISA) | [68] [69] |
| Virus Name | Host Plants | Location | Primer Sequence | Reference |
|---|---|---|---|---|
| Pepino mosaic virus | Tomato | PepMV Ch1 genome | 5′TTTGAGTATATCCCTGGTTC3′ 5′GTTAGCTAAACTACGTACAT3′ | [84] |
| Pepino mosaic virus | Tomato | PepMV Ch2 genome | 5′TTTGAGTATATCCCTGGTTC3′ 5′AATGGGATCCAAGCATTCCAG3′ | |
| Tomato brown rugose fruit virus (ToBRFV) | Tomato | Tobamoviruses Universal primer | 5′GGGAATCAGTTTCAAACRCA3′ 5′GGGGGGATTCGAACCYCT3′ | [85] |
| tomato brown rugose fruit virus (ToBRFV) | Tomato | ToBRFV genome | 5′GAAGTCCCGATGTCTGTAAGG3′ 5′GTGCCTACGGATGTGTATGA3′ | [86] |
| Potato virus M (PVM) | Pepino | Coat protein | 5′TAAGGTAAATCTGAAATAGTG3′ 5′GCCACCCTGGTTACGTGCTT3′ | [81] |
| Pepino mosaic virus (PepMV) | Pepino | Coat protein | 5′ATGCCTGACACAACACCTGT3′ 5′TTAAAGTTCAGGGGGTGCG3′ | |
| Tomato mosaic virus (ToMV) | Pepino | Coat protein | 5′ATGTCTTACTCAATCACTTC3′ 5′TTAAGATGCAGGTGCAGAGG3′ | |
| Potato virus S (PVS) | Pepino | Partial Coat protein | 5′ARCACCTTTAGGTTCACAG3′ 5′TTGARAAWCGCGAGTATG3′ | |
| Tobacco mosaic virus (TMV) | Capsicum | Replicase gene | 5′CGACATCAGCCGATGCAGC3′ 5′ACCGTTTTCGAACCGAGACT3′ | [87] |
| Tomato mosaic virus (ToMV) | Tomato | 5′CGAGAGGGGCAACAAACAT3′ 5′ACCTGTCTCCATCTCTTTGG3′; | ||
| Tomato mosaic virus (ToMV) | Tomato, Tobacco | Genome sequence | 5′GGGCCATATGGGGGCCA3′ 5′ATCCCCGGGT3′ | [88] |
| Cucumber mosaic virus (CMV) | Cucurbit, pumpkin, snake gourd, bitter gourd | Coat protein | 5′GAGTTCTTCCGCGTCCCGCT’3 5′AAACCTAGGAGATGGTTTCA’3 | [89] |
| Papaya ringspot virus (PRSV) | 5′GCAATGATAGARTCATGGGG’3 5′AAGCGGTGGCGCAGCCACACT’3 | |||
| Zucchini yellow mosaic virus (ZYMV) | 5′ATAGCTGAGACAGCACT’3 5CGGCAGCRAAACGATAACCT3 | |||
| Cucumber Green Mottle Mosaic Virus (CGMMV) | 5′TAAGCGGCATTCTAAACCTCCA’3 5′CACTATGCACTTTGGTGTGC’3 | |||
| Begomovirus | 5′ATGKYGAAGCGACCAGCMGA’3 5′CGCCCKCMGAYTGGGMTTTTCTT’3 | |||
| Tobacco mosaic virus (TMV) | Tomato, Bell paper | Movement protein | 5′TGAAAATGAATCATTGTCTG3′ 5′ACTCATCAACAACTTCTTCC3′ | [90] |
| Tomato mosaic virus (ToMV) | 5′TGAAAATGAATCATTGTCTA3′ 5′CATCTTCAATCAAATTATC3′ | |||
| Tomato leaf curl New Delhi virus (ToLCNDV) | Tomato, Eggplant, Cucurbits | Begomovirus universal primer | 5′TAATATTACCKG WKGVCSC3′ 5′TGGACYTTRCAWGGBCCTTCACA3′ | [91] |
| Tomato leaf curl Gujarat virus (ToLCGV) | 5′TAATATTACCKG WKGVCSC3′ 5′TGGACYTTRCAWGGBCCTTCACA3′ | |||
| Tomato leaf curl Palampur virus (ToLCPalV) | 5′TAATATTACCKG WKGVCSC3′ 5′TGGACYTTRCAWGGBCCTTCACA3′ | |||
| Tomato leaf curl Bangalore virus (ToLCBV) | 5′TAATATTACCKG WKGVCSC3′ 5′TGGACYTTRCAWGGBCCTTCACA3′ | |||
| Tomato leaf curl New Delhi virus (ToLCNDV) | Tomato, Eggplant, Cucurbits | Begomovirus universal primer | 5′ACNGGNAA RACNATGTGGGC3′ 5′GGNAARTHTGGATGGA3′ | [1] |
| Tomato leaf curl Gujarat virus (ToLCGV) | 5′ACNGGNAA RACNATGTGGGC3′ 5′GGNAARTHTGGATGGA3′ | |||
| Tomato leaf curl Palampur virus (ToLCPalV) | 5′ACNGGNAA RACNATGTGGGC3′ 5′GGNAARTHTGGATGGA3′ | |||
| Tomato leaf curl Bangalorevirus (ToLCBV) | 5′ACNGGNAA RACNATGTGGGC3′ 5′GGNAARTHTGGATGGA3′ | |||
| Tomato leaf curl New Delhi virus (ToLCNDV) | Tomato | Begomovirus coat protein | 5′ATGKYGAAGCGACCAGCMGA3′ 5′CGCCCKCMGAYTGGGMTTTTCTT3′ | [92] |
| Tomato leaf curl Gujarat virus (ToLCGV) | 5′ATGKYGAAGCGACCAGCMGA3′ 5′CGCCCKCMGAYTGGGMTTTTCTT3′ | |||
| Tomato leaf curl Palampur virus (ToLCPalV) | 5′ATGKYGAAGCGACCAGCMGA3′ 5′CGCCCKCMGAYTGGGMTTTTCTT3′ | |||
| Tomato leaf curl Bangalorevirus (ToLCBV) | 5′ATGKYGAAGCGACCAGCMGA3′ 5′CGCCCKCMGAYTGGGMTTTTCTT3′ | |||
| Citrus tristeza virus (CTV) | Citrus | Coat protein | 5′-CGCCATAACTCAAGTTGCG-3′ 5′-GTGGCCAATAGGTCCGTAGA-3′ | [93] |
| Cucumber mosaic virus (CMV) | Tomato | Coat protein | 5′GAGTTCTTCCGCGTCCCGCT3′ 5′AAACCTAGG AGATGGTTTCA3′ | [94] |
| Tobacco mosaic virus (TMV) | Universal primer tobamovirus | 5′ATTTAAGTGGAGGGAAAACCACT3′ 5′GTYGTTGATGAGTTCGTGGA3′ | ||
| Tobacco mosaic virus (TMV) | Cucumber green mottle mosaic virus | 5′TAAG CGGCATTCTAAACCTCCA3′ 5′CACTATGCACTTTG GTGTGC3′ | ||
| Tomato mosaic virus (ToMV) | Universal primer tobamovirus | 5′ATTTAAGTGGAGGGAAAACCACT3′ 5′GTYGTTGATGAGTTCGTGGA3′ | ||
| Tomato mosaic virus (ToMV) | Cucumber green mottle mosaic virus | 5′TAAG CGGCATTCTAAACCTCCA3′ 5′CACTATGCACTTTG GTGTGC3′ | ||
| Potato virus Y (PVY) | Universal potyvirus primer | 5′GGBAAYAATAGTGGNCAACC3′ 5′GGGGAGGTGCCGTTCTCDATRCACCA3′ | ||
| Potato virus Y (PVY) | Universal potyvirus primer | 5′GTITGYGTIGAYGAYTTYAAYAA3′ 5′TCIACIACIGTIGAIGGYTGNCC3′ | ||
| Potato virus Y (PVY) | Papaya ringspot virus coat protein | 5′GCAATGATAGARTC ATGGGG3′ 5′AAGCGGTGGCGCAGCCACACT3′ | ||
| Zucchini yellow mosaic | 5′ATAGCTGAGACA GCACT3′ 5′CGGCAGCRAAACGATAACCT3 | |||
| Begomovirus | Clove basil | Coat protein | 5′ATGGCGAAGCGACCAG3′ 5′TTAATTTGTGACCGAATCAT3 | [95] |
| Potyvirus | 5′GTITGYGTIGAYGAYTTYAAYAA3′ 5′TCIACIACIGTIGAIGGYTGNCC3′ | |||
| Cucumber mosaic virus (CMV) | 5′GCATTCTAGATGGACAAATCTGAATC3 5′GCATGGTACCTCAAACTGGGAGCAC3′ | |||
| Apple chlorotic leaf spot virus (ACLSV) | Pear | Coat protein | 5′CCGAATTCATGGCAGCAGTTCTGAATC3′ 5′GAGAGCTCCTAGATGCAAAGATCAG3′ | [96] |
| Apple stem pitting virus (ASPV) | 5′ATGTCTGGAACCTCATGCTGCAA3′ 5′TTGGGATCAACTTTACTAAAAAGCATAA3′ | |||
| Tobacco mosaic virus (TMV) | Tomato | Coat protein | 5′TCTTGTCATCAGCGTGGGC3′ 5′CCAGAGGTCCAAACCAAACCA3′ | [97] |
| Cucumber mosaic virus (CMV) | 5′TGGACAAATCTGAATCAACCAGTG3′ 5′GTACTAGCTCGTCCGTCTCG3′ | |||
| Potato virus X (PVX) | 5′TCAGCACCAGCTAGCACAAC3′ 5′TGGTGGGAGAGTGACAACAGC3′ | |||
| Potato virus Y (PVY) | 5′ATACTCGGGCAACTCAATCAC3′ 5′GCTTCTGCAACATCTGAGAAATG3′ | |||
| omato spotted wilt virus (TSWV) | 5′CAGAATCTGGTAGCATTCAACTTCA3′ 5′ACTTTTCCTAAGGCTTCCCTG3′ | |||
| Tomato chlorosis virus (ToCV) | 5′AGGGACCTCAGTTAAAGCAGC3′ 5′TCATGACTTCTGGCGTACCG3′ | |||
| Tobamovirus | Laboratory cultivated | Replicase gene | 5′TKGAYGGNGTBCCNGGNTGYGG3′ 5′ACNGAVTBNABCTGTAATTGCTAT3′ | [98] |
| 5′TKGAYGGIGTBCCIGGITGYGG3′ 5′ACIGAVTBIABCTGTAATTGCTAT3′ |
| Host Plant | Disease | Viruses Identified/Findings | Platform Used | Reference |
|---|---|---|---|---|
| Papaya | Papaya ringspot disease | Moroccan watermelon mosaic virus (MWMV) Cowpea mild mottle virus (CpMMV) Papaya mottle-associated virus (PaMV) | Illumina MiSeq | [137] |
| Tomato | Begomovirus disease | Distinct ssDNA virus/subviral agents | Illumina HiSeq | [140] |
| China rose | China rose mosaic disease | Prunus necrotic ringspot virus (PNRSV) | Illumina HiSeq | [131] |
| Grapevine | Leafroll disease | Grapevine leafroll-associated virus 1 (GLRaV-1) and GLRaV-3 | Illumina HiSeq | [124] |
| Grapevine | Grapevine rupestris vein feathering virus (GRVFV | Distinct GRVFV molecular variant | Illumina MiSeq | [141] |
| Tomato, Lettuce | Phytovirome | Virion-associated nucleic acids (VANA) | Illumina HiSeq | [142] |
| Sweet potato | Weet potato virus disease | Begomovirus, Potyvirus and Crinivirus | Illumina MiSeq | [138] |
| Common bean | Virus symptomatic bean plants | Pelargonium vein banding virus (PVBV) Bean common mosaic necrosis virus (BCMNVA) (Potyviridae), Aphid lethal paralysis virus (ALPV) | Illumina HiSeq | [143] |
| Cowpea | Virion-associated nucleic acids (VANA) | Bean common mosaic virus (BCMV) Cowpea mottle virus (CMV) | 454 (GS FLX) | [139] |
| Malus, Prunus, Pyrus | Viruses and viriods | Genotypes of Apple stem pitting virus (ASPV) | Illumina HiSeq | [144] |
| Cucumber | Aphid lethal paralysis virus | Aphid lethal paralysis virus (ALPV) | Illumina MiSeq | [145] |
| Various plant species | Mixed viral infections | Viral/viroid species, putative novel viral species Cytorhabdovirus: CCyV1) | Illumina MiSeq | [146] |
| Common Bean | Bean common mosaic and necrosis disease | Bean common mosaic necrosis virus (BCMNV) Cucumber mosaic virus (CMV) Phaseolus vulgaris alphaendorna viruses 1 and 2 (PvEV1 and 2) | Illumina MiSeq | [147] |
| Common Bean | Virus symptomatic plants. | Pelargonium vein banding virus (PVBV) Bean common mosaic necrosis virus (BCMNVA) | Illumina HiSeq | [143] |
| Apricot | Mixed viral infections | Cherry virus A (CVA) Little cherry virus 1 (LChV-1) | Illumina HiSeq | [148] |
| Prunus | Various viral and viroids infections | Apricot latent virus (ApLV) Apricot vein clearing associated virus (AVCaV) Asian Prunus Virus 2 (APV2) Nectarine stem pitting-associated virus (NSPaV) | Illumina MiSeq | [149] |
| Sweet Cherry | Little cherry viral infection | Little cherry virus 1 (LChV1) | Ion Torrent | [150] |
| Maize | Maize lethal necrosis | Maize chlorotic mottle virus (MCMV) Sugarcane mosaic virus (SCMV), Wheat streak mosaic virus (WSMV) Johnson grass mosaic virus (JGMV). | Illumina MiSeq | [151] |
| Apple | Apple rubbery wood disease | Apple rubbery wood virus (ARWV) 1 and 2 | Illumina HiSeq | [152] |
| Strawberry | Norovirus (NoV) gastroenteritis | Norovirus (NoV) | Illumina HiSeq | [153] |
| Peach, Prunus | Different disease phenotypes | Peach virus D, Viral synergisms involving genera in the Betaflexiviridae, Closteroviridae, and Luteoviridae families | Illumina HiSeq. | [154] |
| Tobacco | Different viral diseases | Cucumber mosaic virus (CMV) Potato virus Y (PVX) Tobacco mosaic virus (TMV) Tobacco vein banding Mosaic virus (TVBMV) Pepper mottle virus (PMV) Brassica yellow virus (BYV) Chilli venial mottle virus (CVMV) | Illumina HiSeq | [155] |
| Prunus | Prunus necrotic ringspot infection | Prunus necrotic ringspot virus (PNRSV) | Illumina MiSeq | [156] |
| Prunus | Mixed virus infections | Prunus necrotic ringspot virus (PNRSV) Prune dwarf virus (PDV) Apple mosaic virus (ApMV) American plum line pattern virus (APLPV) Ilarvirus-like RNA2 amplicon sequences | Illumina MiSeq | [157] |
| Grapevine | Virome of Grapewine | Grapevine fanleaf virus (GFLV) | Illumina HiSeq | [105] |
| Soybean | Pathogen analysis | Bean yellow mosaic virus (BYMV) and other pathogens | Illumina HiSeq | [158] |
| Grapevine | Genetic variability of infected viruses | Grapevine Syrah virus 1 (GSyV-1) Grapevine Cabernet Sauvignon reovirus (GCSV) Grapevine Red Globe virus (GRGV) Grapevine vein clearing virus (GVCV) | Illumina HiSeq | [159] |
| Grapevine | Mixed viral infections | Grapevine rupestris vein feathering virus (GRVFV) Grapevine yellow speckle viroid 1 (GSYVd-1) | Illumina HiSeq | [160] |
| Prunus | Stem-pitting symptoms | Marafivirus. Luteovirus-like viruses | Illumina HiSeq | [14] |
| Vanilla | Viral diseases | Potexvirus, Vanilla virus X (VVX) Vanilla latent virus (VLV) | 454 pyrosequencing | [161] |
| Virus Name | Host Plant | Remark | Reference |
|---|---|---|---|
| Potato virus Y (PVY) | Potato plant | Potato virus Y genetic diversity analyzed | [178] |
| Arabis mosaic virus (ArMV) | Potato plant | First report of ArMV in potato plants | [179] |
| Sri Lankan cassava mosaic virus (SLCMV) | Cassava plant | Complete genome sequencing of SLCMV | [180] |
| Sowthistle yellow vein virus (SYVV) | Sowthistle plant | Historically important plant rhabdovirus | [181] |
| Tomato severe rugose virus (ToSRV) | Tomato weed (Physalis angulata L.) | First report on infection of P. angulata by ToSRV | [182] |
| Cowpea bright yellow mosaic virus (CABMV) | Cowpea plant | Cowpea bright yellow mosaic virus identified | [183] |
| Wheat streak mosaic virus (WSMV) | Wheat plant | Wheat streak mosaic virus identified | [184] |
| Cassava mosaic disease (CMV) | Cassava plant | Cassava mosaic virus identified | [185] |
| Tomato yellow leaf curl virus (TYLCV) | Tomato plant | Tomato yellow leaf curl virus identified | [186] |
| Watermelon chlorotic stunt virus (WmCSV) | Watermelon plant | Chlorotic stunt virus identified | |
| Tomato brown rugose fruit virus (ToBRFV) | Tomato plant | Tomato brown rugose fruit virus identified | |
| Cucumber green mottle mosaic virus (CGMMV) | Tomato plant | Cucumber green mottle mosaic virus identified | |
| Zucchini yellow mosaic virus (ZYMV) | Butternut squash | Zucchini yellow mosaic virus | |
| Dioscorea bacilliform virus (DBV) | Water yam plant | Bacilliform virus identified | [187] |
| Yam mild mosaic virus (YMMV) | Mosaic virus identified | ||
| Yam chlorotic necrosis virus (YCNMV) | Chlorotic necrosis virus identified | ||
| Plum pox virus (PPV) | Prunus plant | Plum pox virus identified | [188] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehetre, G.T.; Leo, V.V.; Singh, G.; Sorokan, A.; Maksimov, I.; Yadav, M.K.; Upadhyaya, K.; Hashem, A.; Alsaleh, A.N.; Dawoud, T.M.; et al. Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review. Viruses 2021, 13, 412. https://doi.org/10.3390/v13030412
Mehetre GT, Leo VV, Singh G, Sorokan A, Maksimov I, Yadav MK, Upadhyaya K, Hashem A, Alsaleh AN, Dawoud TM, et al. Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review. Viruses. 2021; 13(3):412. https://doi.org/10.3390/v13030412
Chicago/Turabian StyleMehetre, Gajanan T., Vincent Vineeth Leo, Garima Singh, Antonina Sorokan, Igor Maksimov, Mukesh Kumar Yadav, Kalidas Upadhyaya, Abeer Hashem, Asma N. Alsaleh, Turki M. Dawoud, and et al. 2021. "Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review" Viruses 13, no. 3: 412. https://doi.org/10.3390/v13030412
APA StyleMehetre, G. T., Leo, V. V., Singh, G., Sorokan, A., Maksimov, I., Yadav, M. K., Upadhyaya, K., Hashem, A., Alsaleh, A. N., Dawoud, T. M., Almaary, K. S., & Singh, B. P. (2021). Current Developments and Challenges in Plant Viral Diagnostics: A Systematic Review. Viruses, 13(3), 412. https://doi.org/10.3390/v13030412