
Fucoidan But Not 2′-Fucosyllactose Inhibits Human Norovirus Replication in Zebrafish Larvae


Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. HNoV Virus-Like Particle (VLP) Binding Block Assay
2.3. Zebrafish Maintenance and Injection of Zebrafish Larvae with hNoV in the Yolk
2.4. Tissue Homogenization, RNA Extraction and RT-qPCR for Detection of hNoV
2.5. Statistical Analysis
3. Results
3.1. Both Fucoidan and 2′-FL Were Able to Block the Binding of hNoV VLPs to HBGAs in Type A Saliva
3.2. HNoV GII.P16-GII.4 Replicated in Zebrafish Larvae More Consistently Than GII.P31-GII.17 at 2 dpi
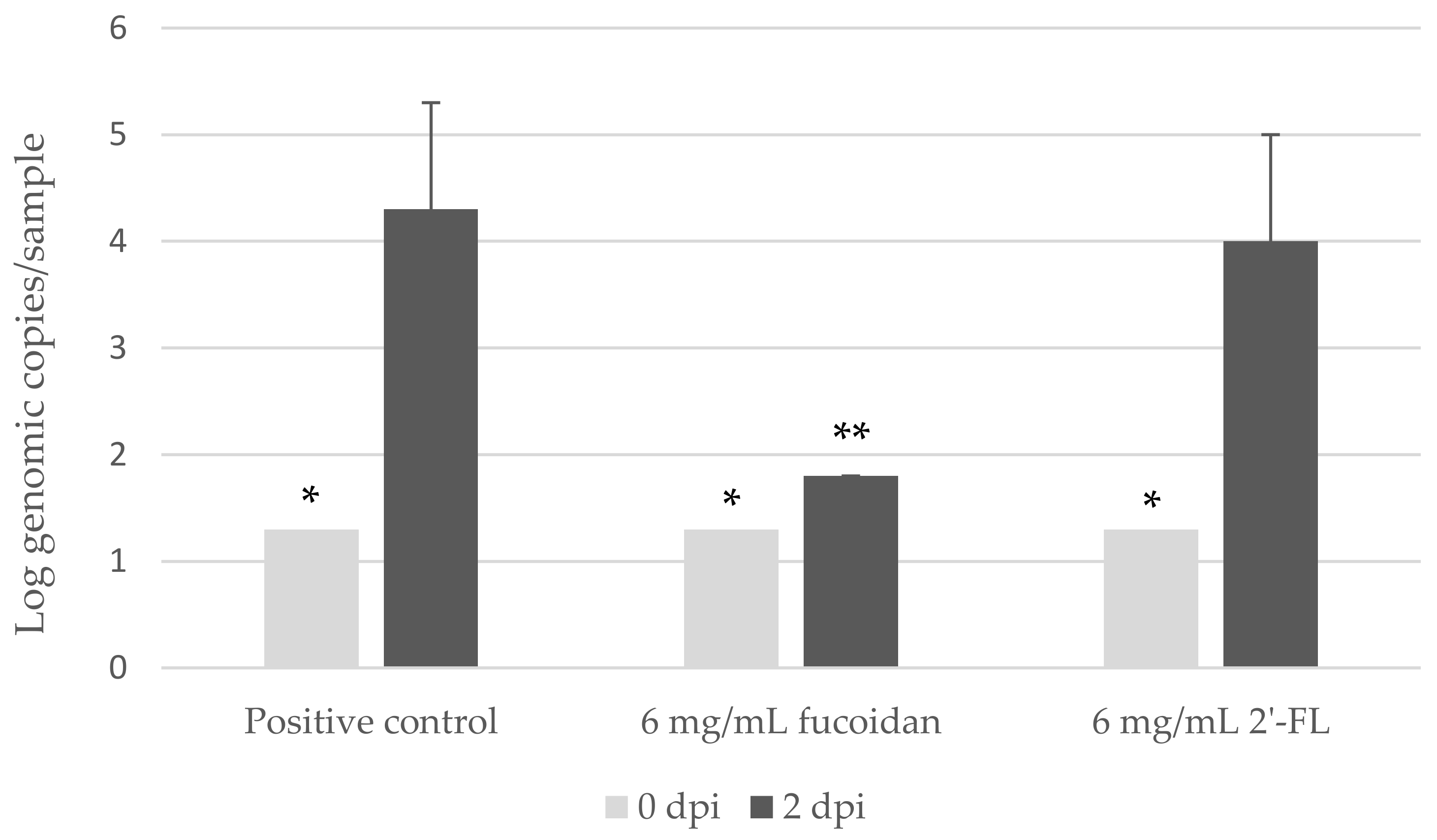
3.3. Fucoidan, but Not 2′-FL, Inhibited hNoV GII.P16-GII.4 Replication in Zebrafish Larvae
3.4. Saliva But Not Human Milk from Type A Secretors Inhibited hNoV GII.P16-GII.4 Replication in Zebrafish Larvae
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- WHO (World Health Organization). WHO Estimates of the Global Burden of Foodborne Diseases: Foodborne Disease Burden Epidemiology Reference Group 2007–2015; WHO: Geneva, Switzerland, 2015; Available online: https://apps.who.int/iris/handle/10665/199350 (accessed on 1 January 2021).
- Iturriza-Gomara, M.; O’Brien, S.J. Foodborne viral infections. Curr. Opin. Infect. Dis. 2016, 29, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Marsh, Z.; Shah, M.P.; Wikswo, M.E.; Barclay, L.; Kisselburgh, H.; Kambhampati, A.; Cannon, J.L.; Parashar, U.D.; Vinjé, J.; Hall, A.J. Epidemiology of Foodborne Norovirus Outbreaks—United States, 2009–2015. Food Saf. 2018, 6, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Chong, P.P.; Atmar, R.L. Norovirus in health care and implications for the immunocompromised host. Curr. Opin. Infect. Dis. 2019, 32, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Hallowell, B.D.; Parashar, U.D.; Hall, A.J. Epidemiologic challenges in norovirus vaccine development. Hum. Vaccines Immunother. 2018, 15, 1279–1283. [Google Scholar] [CrossRef]
- Netzler, N.E.; Enosi Tuipulotu, D.; White, P.A. Norovirus antivirals: Where are we now? Med. Res. Rev. 2018, 39, 860–886. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.; Jiang, X. Norovirus and its histo-blood group antigen receptors: An answer to a historical puzzle. Trends Microbiol. 2005, 13, 285–293. [Google Scholar] [CrossRef]
- Singh, B.K.; Leuthold, M.M.; Hansman, G.S. Structural constraints on human norovirus binding to histo-blood group antigens. MSphere 2016, 1, e00049-16. [Google Scholar] [CrossRef] [Green Version]
- Ali, E.S.; Rajapaksha, H.; Carr, J.M.; Petrovsky, N. Norovirus drug candidates that inhibit viral capsid attachment to human histo-blood group antigens. Antivir. Res. 2016, 133, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Derya, S.M.; Spiegel, H.; Hanisch, F.-G.; Morozov, V.; Schroten, H.; Jennewein, S.; Parschat, K. Biotechnologically produced fucosylated oligosaccharides inhibit the binding of human noroviruses to their natural receptors. J. Biotechnol. 2020, 318, 31–38. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Dai, Y.-C.; Zhong, W.; Tan, M.; Lv, Z.-P.; Zhou, Y.-C.; Jiang, X. Tannic acid inhibited norovirus binding to HBGA receptors, a study of 50 Chinese medicinal herbs. Bioorg. Med. Chem. 2012, 20, 1616–1623. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Tan, M.; Chhabra, M.; Dai, Y.-C.; Meller, J.; Jiang, X. Inhibition of histo-blood group antigen binding as a novel strategy to block norovirus infections. PLoS ONE 2013, 8, e69379. [Google Scholar] [CrossRef] [Green Version]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.-L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef]
- Van Dycke, J.; Ny, A.; Conceição-Neto, N.; Maes, J.; Hosmillo, M.; Cuvry, A.; Goodfellow, I.; Nogueira, T.C.; Verbeken, E.; Matthijnssens, J.; et al. A robust human norovirus replication model in zebrafish larvae. PLoS Pathog. 2019, 15, e1008009. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, J.R.; Jobin, C. Think small: Zebrafish as a model system of human pathology. J. Biomed. Biotechnol. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Burgos, J.S.; Ripoll-Gomez, J.; Alfaro, J.M.; Sastre, I.; Valdivieso, F. Zebrafish as a new model for herpes simplex virus type 1 infection. Zebrafish 2008, 5, 323–333. [Google Scholar] [CrossRef]
- Ding, C.-B.; Zhang, J.-P.; Zhao, Y.; Peng, Z.-G.; Song, D.-Q.; Jiang, J.-D. Zebrafish as a potential model organism for drug test against hepatitis C virus. PLoS ONE 2011, 6, e22921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabor, K.A.; Goody, M.F.; Mowel, W.K.; Breitbach, M.E.; Gratacap, R.L.; Witten, P.E.; Kim, C.H. Influenza A virus infection in zebrafish recapitulates mammalian infection and sensitivity to anti-influenza drug treatment. Dis. Models Mech. 2014, 7, 1227–1237. [Google Scholar] [CrossRef] [Green Version]
- Balla, K.M.; Rice, M.C.; Gagnon, J.A.; Elde, N.C. Linking virus discovery to immune responses visualized during zebrafish infections. Curr. Biol. 2020, 30, 2092–2103. [Google Scholar] [CrossRef]
- Hanisch, F.-G.; Hansman, G.S.; Morozov, V.; Kunz, C.; Schroten, H. Avidity of α-fucose on human milk oligosaccharides and blood group–unrelated oligo/polyfucoses is essential for potent norovirus-binding targets. J. Biol. Chem. 2018, 293, 11955–11965. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lim, C.Y.; Lee, D.B.; Seok, J.H.; Kim, K.H.; Chung, M.S. Inhibitory effects of Laminaria japonica fucoidans against noroviruses. Viruses 2020, 12, 997. [Google Scholar] [CrossRef] [PubMed]
- Koromyslova, A.; Tripathi, S.; Morozov, V.; Schroten, H.; Hansman, G.S. Human norovirus inhibition by a human milk oligosaccharide. Virology 2017, 508, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Schroten, H.; Hanisch, F.-G.; Hansman, G.S. Human norovirus interactions with histo-blood group antigens and human milk oligosaccharides. J. Virol. 2016, 90, 5855–5859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichert, S.; Koromyslova, A.; Singh, B.K.; Hansman, S.; Jennewein, S.; Schroten, H.; Hansman, G.S. Structural basis for norovirus inhibition by human milk oligosaccharides. J. Virol. 2016, 90, 4843–4848. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.D. Multistate outbreak of norwalk-like virus gastroenteritis associated with a common caterer. Am. J. Epidemiol. 2001, 154, 1013–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojima, S.; Kageyama, T.; Fukushi, S.; Hoshino, F.B.; Shinohara, M.; Uchida, K.; Natori, K.; Takeda, N.; Katayama, K. Genogroup-specific PCR primers for detection of Norwalk-like viruses. J. Virol. Methods 2002, 100, 107–114. [Google Scholar] [CrossRef]
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.; Peñaranda, S.; Oberste, M.S.; Vinjé, J.; Koopmans, M. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef]
- International Organization for Standardization. ISO 15216-1 Microbiology of Food and Animal Feed: Horizontal Method for Determination of Hepatitis A Virus and Norovirus in Food Using Real-Time RT-PCR. Part 1: Method for Quantification; International Organization for Standardization: Geneva, Switzerland, 2017. [Google Scholar]
- Nordgren, J.; Svensson, L. Genetic susceptibility to human norovirus infection: An update. Viruses 2019, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic effects of fucoidan: A review on recent studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [Green Version]
- Goehring, K.C.; Marriage, B.J.; Oliver, J.S.; Wilder, J.A.; Barrett, E.G.; Buck, R.H. Similar to those who are breastfed, infants fed a formula containing 2′-fucosyllactose have lower inflammatory cytokines in a randomized controlled trial. J. Nutr. 2016, 146, 2559–2566. [Google Scholar] [CrossRef] [Green Version]
- Schott, M. New Study: Babies Fed Infant Formula with 2′-fl Human Milk Oligosaccharide+ Had Immune Response more like Breastfed Babies. Abbott MediaRoom. 2016. Available online: https://abbott.mediaroom.com/2016-10-26-New-Study-Babies-Fed-Infant-Formula-with-2-FL-Human-Milk-Oligosaccharide-Had-Immune-Response-More-Like-Breastfed-Babies (accessed on 1 January 2021).
- Green, K.Y.; Kaufman, S.S.; Nagata, B.M.; Chaimongkol, N.; Kim, D.Y.; Levenson, E.A.; Tin, C.M.; Yardley, A.B.; Johnson, J.A.; Barletta, A.B.F.; et al. Human norovirus targets enteroendocrine epithelial cells in the small intestine. Nat. Commun. 2020, 11, 2759. [Google Scholar] [CrossRef] [PubMed]
- Almand, E.A.; Moore, M.D.; Jaykus, L.-A. Norovirus binding to ligands beyond histo-blood group antigens. Front. Microbiol. 2017, 8, 2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruvoën-Clouet, N.; Mas, E.; Marionneau, S.; Guillon, P.; Lombardo, D.; Le Pendu, J. Bile-salt-stimulated lipase and mucins from milk of ‘secretor’mothers inhibit the binding of Norwalk virus capsids to their carbohydrate ligands. Biochem. J. 2006, 393, 627–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.; Huang, P.; Zhong, W.; Tan, M.; Farkas, T.; Morrow, A.L.; Newburg, D.S.; Ruiz-Palacios, G.M.; Pickering, L.K. Human milk contains elements that block binding of noroviruses to human histo-blood group antigens in saliva. J. Infect. Dis. 2004, 190, 1850–1859. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| (A) | ||||
| Batch | Sample | Log Genome Copies/Sample | ||
| 0 dpi | 2 dpi | 3 dpi | ||
| Batch 1 | Sample 1 | 1.7 | 4.7 | 4.7 |
| Sample 2 | 1.5 | 4.4 | 4 | |
| Batch 2 | Sample 1 | 1.9 | 4.6 | 4.8 |
| Batch 3 | Sample 1 | <1.3 | 4.8 | 2.5 |
| Batch 4 | Sample 1 | <1.3 | 4.9 | 2.6 |
| (B) | ||||
| Batch | Sample | Log Genome Copies/Sample | ||
| 0 dpi | 2 dpi | 3 dpi | ||
| Batch 1 | Sample 1 | 2.5 | n.a. | 3.4 |
| Sample 2 | 2.5 | n.a. | 2.3 | |
| Batch 2 | Sample 1 | 4.3 | n.a. | 4.4 |
| Batch 3 | Sample 1 | 2.8 | 2.9 | n.a. |
| Batch 4 | Sample 1 | 3 | 4.8 | n.a. |
| (A) | |||
| Concentration of Fucoidan | Zebrafish Larvae Survival Rate at 2 dpi | Log Genome Copies/Sample | |
| 0 dpi | 2 dpi | ||
| 24.0 mg/mL | <90% | <1.3 | n.a. |
| 12.0 mg/mL | <90% | <1.3 | n.a. |
| 6.00 mg/mL | >90% | <1.3 | <1.3 |
| 2.40 mg/mL | >90% | <1.3 | <1.3 |
| 0.24 mg/mL | >90% | <1.3 | 4.3 |
| 0 | >90% | <1.3 | 4.7 |
| (B) | |||
| Concentration of 2′-FL | Zebrafish Larvae Survival Rate at 2 dpi | Log Genome Copies/Sample | |
| 0 dpi | 2 dpi | ||
| 24.0 mg/mL | >90% | 1.4 | 5.3 |
| 6.00 mg/mL | >90% | 1.9 | 3.9 |
| 2.40 mg/mL | >90% | 2.3 | 4.2 |
| 0 | >90% | 1.9 | 4.1 |
| (C) | |||
| Zebrafish Larvae Survival Rate at 2 dpi | Log Genome Copies/Sample | ||
| 0 dpi | 2 dpi | ||
| Saliva (type A secretor) | >90% | 2.4 | 2.2 |
| Control | >90% | 1.9 | 4.1 |
| (D) | |||
| Zebrafish Larvae Survival Rate at 2 dpi | Log Genome Copies/Sample | ||
| 0 dpi | 2 dpi | ||
| Human milk (type A secretor) | >90% | <1.3 | 5.7 |
| Control | >90% | <1.3 | 5.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, M.T.H.; Li, Y.; Eshaghi Gorji, M.; Gong, Z.; Li, D. Fucoidan But Not 2′-Fucosyllactose Inhibits Human Norovirus Replication in Zebrafish Larvae. Viruses 2021, 13, 461. https://doi.org/10.3390/v13030461
Tan MTH, Li Y, Eshaghi Gorji M, Gong Z, Li D. Fucoidan But Not 2′-Fucosyllactose Inhibits Human Norovirus Replication in Zebrafish Larvae. Viruses. 2021; 13(3):461. https://doi.org/10.3390/v13030461
Chicago/Turabian StyleTan, Malcolm Turk Hsern, Yan Li, Mohamad Eshaghi Gorji, Zhiyuan Gong, and Dan Li. 2021. "Fucoidan But Not 2′-Fucosyllactose Inhibits Human Norovirus Replication in Zebrafish Larvae" Viruses 13, no. 3: 461. https://doi.org/10.3390/v13030461