
Sequence Length of HIV-1 Subtype B Increases over Time: Analysis of a Cohort of Patients with Hemophilia over 30 Years



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Patients and Samples
2.3. RNA/DNA Preparation and FL Gene Amplification
2.4. Phylogenetic Tree Analysis
2.5. Viral Signature Pattern Analysis (VESPA)
2.6. Statistical Analysis
2.7. Nucleotide Sequence Data
3. Results
3.1. Origin of the KSB of Subtype B
3.2. Molecular Epidemiologic Data on the FL HIV-1 Gene
3.3. Korean Signature Pattern Amino Acid Residues
3.4. Sequence Identities of HPs Compared to Plasma Donors O and P
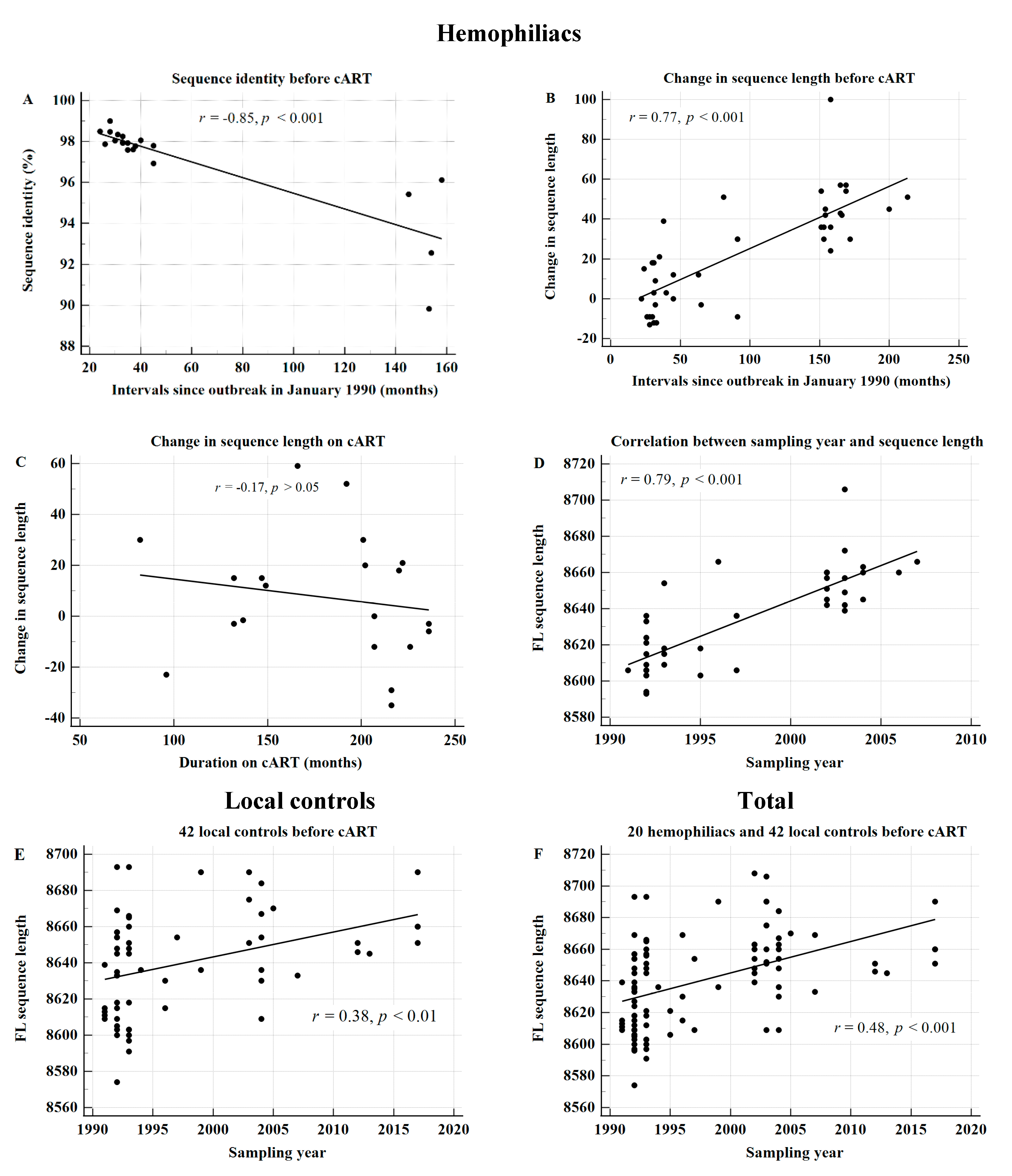
3.5. Sequence Length of HIV-1 KSB Significantly Increases over Time before cART Administration
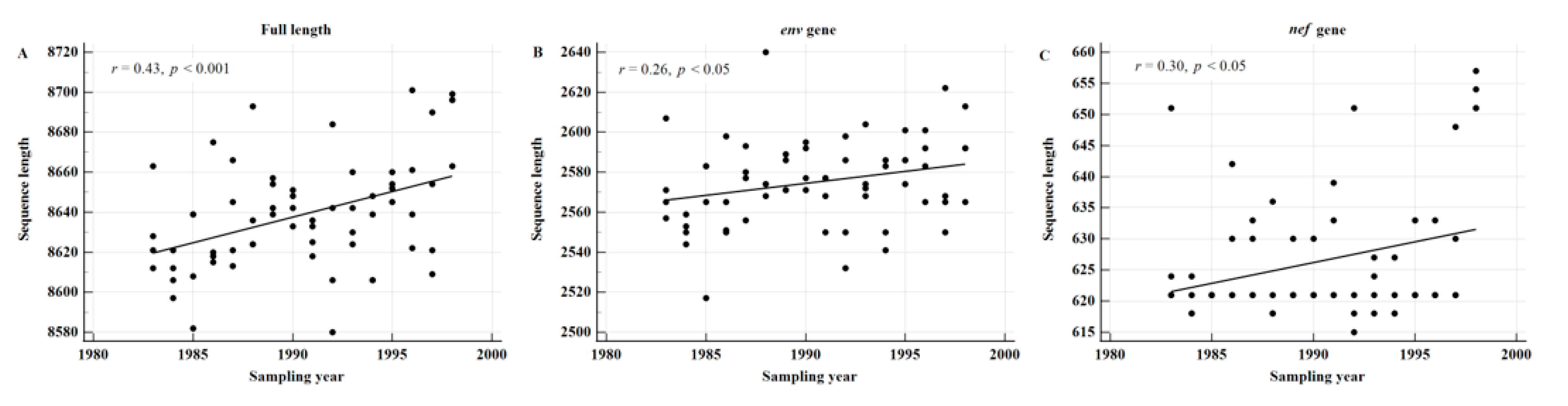
3.6. The Sequence Length in HIV-1 Subtype B Significantly Increases over Time
3.7. Clinical Significance of the Increase in Sequence Length in HIV-1 Subtype B
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cho, Y.K.; Foley, B.T.; Sung, H.; Kim, Y.B.; Kim, J.H. Molecular epidemiologic study of a human immunodeficiency virus 1 outbreak in hemophiliacs B infected through clotting factor 9 after 1990. Vox Sang. 2007, 92, 113–120. [Google Scholar] [CrossRef]
- Cho, Y.K.; Jung, Y.S.; Foley, B.T. Phylogenetic analysis of full-length pol gene from Korean hemophiliacs and plasma donors infected with Korean subclade B of HIV-1. AIDS Res. Hum. Retrovir. 2011, 27, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.K.; Jung, Y.; Lee, J.S.; Foley, B.T. Molecular evidence of HIV-1 transmission in 20 Korean individuals with hemophilia; phylogenetic analysis of vif gene. Haemophilia 2012, 18, 291–299. [Google Scholar] [CrossRef]
- Cho, Y.K.; Kim, J.E.; Foley, B.T. Phylogenetic analysis of the earliest nef gene from hemophiliacs and local controls in Korea. BioRes. Open Access 2012, 1, 41–49. [Google Scholar]
- Cho, Y.K.; Kim, J.E.; Jeong, D.; Foley, B.T. Signature pattern analysis for the full-length env gene of the earliest Korean subclade B of HIV-1: Outbreak among Korean hemophiliacs. Virus Genes 2017, 53, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.K.; Kim, J.E.; Foley, B.T. Genetic analysis of the full-length gag gene from the earliest Korean subclade B of HIV-1: An outbreak among Korean hemophiliacs. Viruses 2019, 11, 545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.K.; Kim, J.E.; WOO, J.H. Korean Red Ginseng increases defective pol gene in peripheral blood mononuclear cells of HIV-1-infected patients; inhibition of its detection during ginseng-based combination therapy. J. Ginseng Res. 2019, 43, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Leitner, T.; Escanilla, D.; Franzén, C.; Uhlén, M.; Albert, J. Accurate reconstruction of a known HIV-1 transmission history by phylogenetic tree analysis. Proc. Natl. Acad. Sci. USA 1996, 93, 10864–10869. [Google Scholar] [CrossRef] [Green Version]
- Novitsky, V.; Moyo, S.; Lei, Q.; DeGruttola, V.; Essex, M. Importance of viral sequence length and number of variable and informative sites in analysis of HIV clustering. AIDS Res. Hum. Retrovir. 2015, 31, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Daniels, R.S.; Kang, C.; Patel, D.; Xiang, Z.; Douglas, N.W.; Zheng, N.N.; Cho, H.W.; Lee, J.S. An HIV type 1 Subtype B founder effect in Korea: gp160 signature patterns infer circulation of CTL-escape strains at the population level. AIDS Res. Hum. Retrovir. 2003, 19, 631–641. [Google Scholar] [CrossRef]
- Korber, B.; Myers, G. Signature pattern analysis: A method for assessing viral sequence relatedness. AIDS Res. Hum. Retrovir. 1992, 8, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Korber, B.T.; Foley, B.; Gaschen, B.; Kuiken, C. Epidemiological and Immunological Implications of the Global Variability of HIV-1; Pataleo, G., Walker, B.D., Eds.; Humana Press: Totowa, NJ, USA, 2001; pp. 1–32. ISBN 978-1-61737-128-8. [Google Scholar]
- Junqueira, D.M.; Almeida, S.E. HIV-1 subtype B: Traces of a pandemic. Virology 2016, 495, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Jang, S.Y.; Park, C.S.; Lee, K.M.; Lee, D.H.; Lee, C.H. Timing and evolution of the most recent common ancestor of the Korean clade HIV subtype B based on nef and vif sequences. J. Microbiol. 2009, 47, 85–90. [Google Scholar] [CrossRef]
- Easterbrook, P.J.; Smith, M.; Mullen, J.; O’Shea, S.; Chrystie, I.; de Ruiter, A.; Tatt, L.D.; Geretti, A.M.; Zuckerman, M. Impact of HIV-1 viral subtype on disease progression and response to antiretroviral therapy. J. Int. AIDS Soc. 2010, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.K.; Kim, J.E.; Foley, B.T. Phylogenetic analysis of near full-length HIV-1 genomic sequences from 21 Korean individuals. AIDS Res. Hum. Retrovir. 2013, 29, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.K.; Sung, H.; Bae, I.G.; Oh, H.B.; Kim, N.J.; Woo, J.H.; Kim, Y.B. Full sequence of HIV type 1 Korean subtype B in an AIDS case with atypical seroconversion: TAAAA at TATA box. AIDS Res. Hum. Retrovir. 2005, 21, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Bio. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.Y.; Ciesielski, C.A.; Myers, G.; Bandea, C.I.; Luo, C.C.; Korber, B.T.; Mullins, J.I.; Schochetman, G.; Berkelman, R.L.; Economou, A.N.; et al. Molecular epidemiology of HIV transmission in a dental practice. Science 1992, 256, 1165–1171. [Google Scholar] [CrossRef] [Green Version]
- Park, C.S.; Kim, M.S.; Lee, S.D.; Kim, S.S.; Lee, K.M.; Lee, C.H. Molecular phylogenetic analysis of HIV-1 vif gene from Korean isolates. J. Microbiol. 2006, 44, 655–659. [Google Scholar]
- Park, C.S.; Lee, D.H.; Lee, K.M.; Lee, C.H. Characterization and signature pattern analysis of Korean clade HIV-1 using nef gene sequences. J. Microbiol. 2008, 46, 88–94. [Google Scholar] [CrossRef]
- Cho, Y.K.; Kim, J.E.; Woo, J.H. Genetic defects in the nef gene are associated with Korean Red Ginseng intake: Monitoring of nef 452 sequence polymorphisms over 20 years. J. Ginseng Res. 2017, 41, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.K.; Kim, J.E.; Lee, S.U.; Foley, B.T.; Choi, B.S. Impact of HIV-1 subtype and Korean Red Ginseng on AIDS progression: Comparison of subtype B and subtype D. J. Ginseng Res. 2019, 43, 312–319. [Google Scholar] [CrossRef] [PubMed]
- Shankarappa, R.; Margolick, J.B.; Gange, S.J.; Rodrigo, A.G.; Upchurch, D.; Farzadegan, H.; Gupta, P.; Rinaldo, C.R.; Learn, G.H.; He, X.; et al. Consistent viral evolutionary changes associated with the progression of human immunodeficiency virus type 1 infection. J. Virol. 1999, 73, 10489–10502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagar, M.; Wu, X.; Lee, S.; Overbaugh, J. Human immunodeficiency virus type 1 V1–V2 envelope loop sequences expand and add glycosylation sites over the course of infection, and these modifications affect antibody neutralization sensitivity. J. Virol. 2006, 80, 9586–9598. [Google Scholar] [CrossRef] [Green Version]
- Bunnik, E.M.; Euler, Z.; Welkers, M.R.; Boeser-Nunnink, B.D.; Grijsen, M.L.; Prins, J.M.; Schuitemaker, H. Adaptation of HIV-1 envelope gp120 to humoral immunity at a population level. Nat. Med. 2010, 16, 995–997. [Google Scholar] [CrossRef]
- Malim, M.H.; Emerman, M. HIV-1 sequence variation, drift, shift, and attenuation. Cell 2001, 104, 469–472. [Google Scholar] [CrossRef] [Green Version]
- Silver, Z.A.; Dickinson, G.M.; Seaman, M.S.; Desrosiers, R.C. A highly unusual V1 region of Env in an elite controller of HIV infection. J. Virol. 2019, 93, e00094-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shioda, T.; Oka, S.; Xin, X.; Liu, H.; Harukuni, R.; Kurotani, A.; Fukushima, M.; Shiino, T.; Takebe, Y.; Lwamoto, A.; et al. In vivo sequence variability of human immunodeficiency virus type 1 envelope gp120: Association of V2 extension with slow disease progression. J. Virol. 1997, 71, 4871–4881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Spira, T.J.; Owen, S.; Lal, R.B.; Saksena, N.K. HIV-1 strains from a cohort of American subjects reveal the presence of a V2 region extension unique to slow progressors and non-progressors. AIDS 2000, 14, 213–223. [Google Scholar] [CrossRef]
- Masciotra, S.; Owen, S.M.; Rudolph, D.; Yang, C.; Wang, B.; Saksena, N.; Spira, T.; Dhawan, S.; Lal, R.B. Temporal relationship between V1V2 variation, macrophage replication, and coreceptor adaptation during HIV-1 disease progression. AIDS 2000, 16, 1887–1898. [Google Scholar] [CrossRef]
- Daniels, R.S.; Wilson, P.; Patel, D.; Longhurst, H.; Pattersom, S. Analysis of full-length HIV type 1 env genes indicates differences between the virus infecting T cells and dendritic cells in peripheral blood of infected patients. AIDS Res. Hum. Retrovir. 2004, 20, 409–413. [Google Scholar] [CrossRef]
- Hirao, K.; Andrews, S.; Kuroki, K.; Kusaka, H.; Tadokoro, T.; Kita, S.; Ose, T.; Rowland-Jones, S.L.; Maenaka, K. Structure of HIV-2 Nef reveals features distinct from HIV-1 involved in immune regulation. iScience 2020, 23, 100758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, M.; Münch, J.; Kutsch, O.; Li, H.; Santiago, M.L.; Bibollet-Ruche, F.; Müller-Trutwin, M.C.; Novembre, F.J.; Peeters, M.; Courgnaud, V.; et al. Nef-mediated suppression of T cell activation was lost in a lentiviral lineage that gave rise to HIV-1. Cell 2006, 125, 1055–1067. [Google Scholar] [CrossRef] [Green Version]
- Ariën, K.K.; Troyer, R.M.; Gali, Y.; Colebunders, R.L.; Arts, E.J.; Vanham, G. Replicative fitness of historical and recent HIV-1 isolates suggests HIV-1 attenuation over time. AIDS 2005, 19, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Venner, C.M.; Nankya, I.; Kyeyune, F.; Demers, K.; Kwok, C.; Chen, P.L.; Rwambuya, S.; Munjoma, M.; Chipato, T.; Byamugisha, J.; et al. Infecting HIV-1 subtype predicts disease progression in women of sub-Saharan Africa. EBioMedicine 2016, 13, 305–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, L.D.; de Moura, A.P.S.; Ciandrini, L. Gene length as a regulator for ribosome recruitment and protein synthesis: Theoretical insights. Sci. Rep. 2017, 7, 17409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, A.; Iwatani, Y. APOBEC3G-Mediated G-to-A hypermutation of the HIV-1 Genome: The missing link in antiviral molecular mechanisms. Front. Microbiol. 2016, 7, 2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nested PCR | Primer | Sequences (5′-3′) | |
|---|---|---|---|
| For the full-length gag gene [6] | |||
| First PCR | 503k | 5′-CCKTCTGTTGTGTGACTCTGGTAA-3′ | forward |
| 524 | 5′-CATTGTTTAACTTTTGGGCCATCC -3′ | reverse | |
| Second PCR | 504F | 5′-TCTCTAGCAGTGGCGCCCGAAC-3′ | forward |
| 505 | 5′-GAGACATGGGTGCGAGAGCGT-3′ | forward | |
| 522 | 5′-ACTGTCCTACTTTGATAAAACCTC-3′ | reverse | |
| For the full-length pol gene [2,7] | |||
| First PCR | HXB2 | 5′-GTGGGAGAAATCTATAAAAGATGG-3′ | forward |
| OBP2 | 5′-GAGACTCCCTGACCCAGATG-3′ | reverse | |
| OBP2k | 5′-GAGACTCCCTGACCCAGATG-3′ | reverse | |
| 550 | 5′-CCTAGTGGGATGTGTACTTCTGAA-3′ | reverse | |
| Second PCR | PO1 | 5′-AAAATTGCAGGGCCCCTAGGA-3′ | forward |
| PR3-1 | 5′-GAAGCAGGAGCCGATAGACA-3′ | forward | |
| OBP4 | 5′-CAATCATCACCTGCCATCTG-3′ | reverse | |
| P2 | 5′-AGGAAGGACACCAAATGAAAG-3′ | forward | |
| P16 | 5′-GGATKAGTGCTTTCATAGTGA-3′ | reverse | |
| For vif, vpr, tat, rev, and vpu genes | |||
| First PCR | 545 | 5′-GCAGTACAAATGGCAGTATTCATC-3′ | forward |
| KMK2 | 5′-ATGGGAATTGGTTCAAAGGA-3′ | reverse | |
| Second PCR | 548F | 5′-AGTGACATAAAAGTAGTRCCAAGAA-3′ | forward |
| LA106 | 5′-TTCACTCTCATTGCCACT-3′ | reverse | |
| For the full-length env gene [5] | |||
| First PCR | OWE1 | 5′-TCATCAAGTTTCTCTATCAAAGCA-3′ | forward |
| OWE2 | 5′-TCTGACTGGAAAGCCCACTT-3′ | reverse | |
| Second PCR | OWE3 | 5′-GCAATATTAGCAATAGTTGTGTGG-3′ | forward |
| OWE4 | 5′-ATACTGCTCCCACCCCTTCT-3′ | reverse | |
| For nef gene [4] | |||
| First PCR | Nef5′5 | 5′-AGGATTGTGGAACTTCTGGGAC-3′ | forward |
| LTR3 | 5′-AGGCTCAGATCTGGTCTAAC-3′ | reverse | |
| Second PCR | Nef3 | 5′-ATGGGTGGCAAGTGGTCAAA-3′ | forward |
| N10 | 5′-CGTCCAGAATTCGGAAAGTCCCCAGCGGAAAGT-3′ | reverse | |
| Sequences with Intervals | Gag | Pol | Vif | Vpr | Tat | Rev | Vpu | Env | Nef | FL |
|---|---|---|---|---|---|---|---|---|---|---|
| First sequences (n = 16) | 1503 ± 0 | 3012 ± 0 | 579 | 291 | 215 | 76 | 247 ± 2 | 2563 ± 19 | 622.5 ± 2 | 8620 ± 20 |
| Second sequences (n = 16) | 1504 ± 3 | 3011 ± 3 | 579 | 291 | 215 | 76 | 247 ± 2 | 2595 ± 22 | 627.2 ± 8 | 8659 ± 23 |
| p-value | NS | NS | NS | NS | NS | NS | NS | < 0.001 | < 0.05 | = 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, Y.-K.; Kim, J.-E.; Foley, B.T. Sequence Length of HIV-1 Subtype B Increases over Time: Analysis of a Cohort of Patients with Hemophilia over 30 Years. Viruses 2021, 13, 806. https://doi.org/10.3390/v13050806
Cho Y-K, Kim J-E, Foley BT. Sequence Length of HIV-1 Subtype B Increases over Time: Analysis of a Cohort of Patients with Hemophilia over 30 Years. Viruses. 2021; 13(5):806. https://doi.org/10.3390/v13050806
Chicago/Turabian StyleCho, Young-Keol, Jung-Eun Kim, and Brian T. Foley. 2021. "Sequence Length of HIV-1 Subtype B Increases over Time: Analysis of a Cohort of Patients with Hemophilia over 30 Years" Viruses 13, no. 5: 806. https://doi.org/10.3390/v13050806