
Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling the Phytopathogens
2.2. Differentiation of Pectobacterium spp.
2.3. Testing the Strains for Phage Susceptibility
2.4. Phage Propagation and Purification
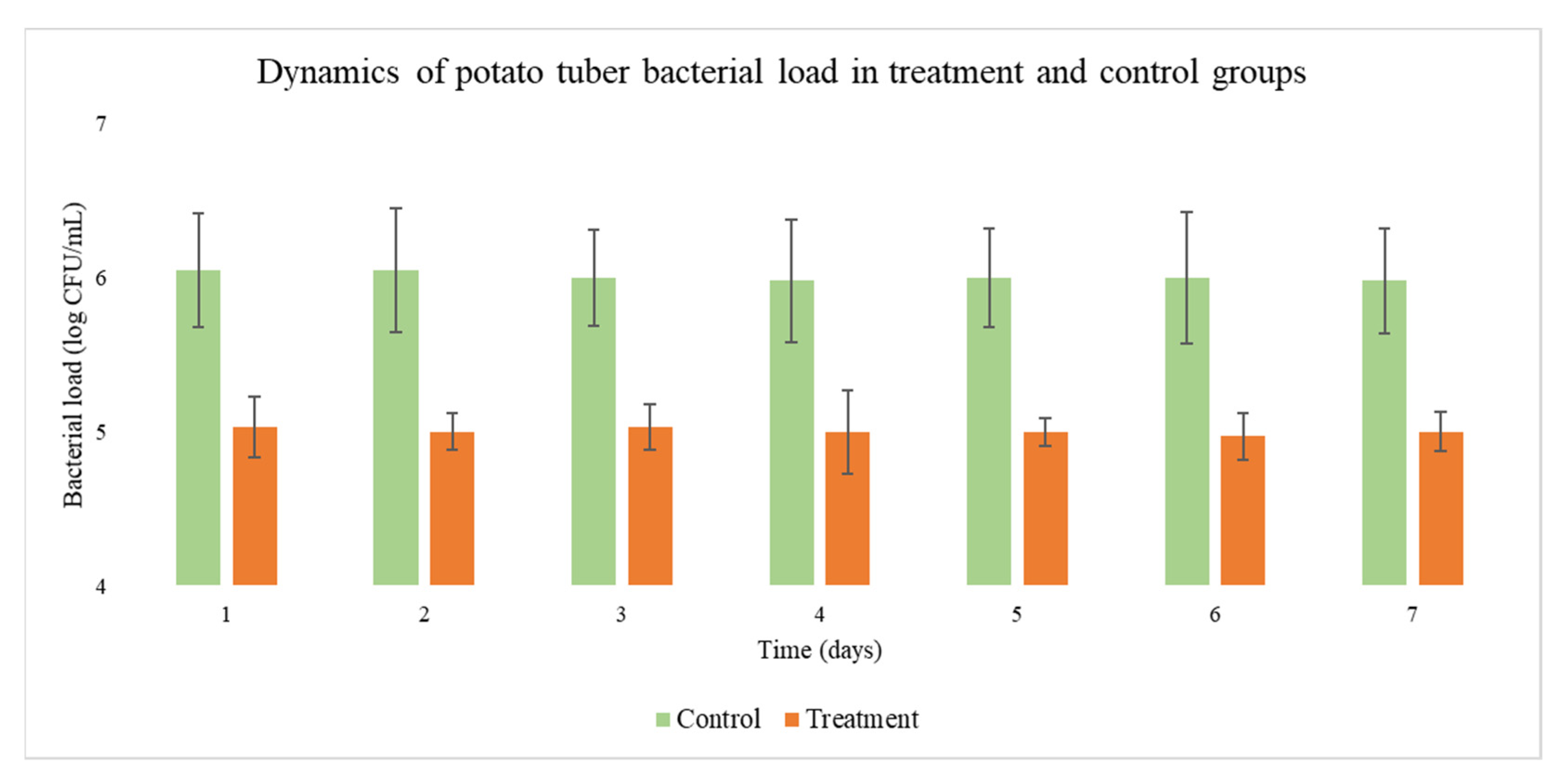
2.5. Application of Phage Cocktails in the Warehouse
2.6. Control of Phage and Bacterial Concentration in the Warehouse Environment
2.7. Statistical Analysis
3. Results
3.1. Phytopathogens Causing Soft Rot of Potato in Northern Moscow Region in 2019
3.2. Construction of the Pathogen-Adapted Phage Cocktail
3.3. Construction of the Pathogen-Adapted Phage Cocktail
3.4. Probing the Phage-Resistant Bacteria Causing Soft Rot
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charkowski, A.O. The changing face of bacterial soft-rot diseases. Annu. Rev. Phytopathol. 2018, 56, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Svircev, A.; Roach, D.; Castle, A. Framing the future with bacteriophages in agriculture. Viruses 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttimer, C.; McAuliffe, O.; Ross, R.P.; Hill, C.; O’Mahony, J.; Coffey, A. Bacteriophages and bacterial plant diseases. Front. Microbiol. 2017, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtappels, D.; Fortuna, K.; Lavigne, R.; Wagemans, J. The future of phage biocontrol in integrated plant protection for sustainable crop production. Curr. Opin. Biotechnol. 2021, 68, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Carstens, A.; Djurhuus, A.; Kot, W.; Jacobs-Sera, D.; Hatfull, G.; Hansen, L. Unlocking the potential of 46 new bacteriophages for biocontrol of Dickeya solani. Viruses 2018, 10, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.A.; Jee, S.; Lee, D.H.; Roh, E.; Jung, K.; Oh, C.; Heu, S. Biocontrol of Pectobacterium carotovorum subsp. carotovorum using bacteriophage PP1. J. Microbiol. Biotechnol. 2013, 23, 1147–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstens, A.B.; Djurhuus, A.M.; Kot, W.; Hansen, L.H. A novel six-phage cocktail reduces Pectobacterium atrosepticum soft rot infection in potato tubers under simulated storage conditions. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Voronina, M.V.; Bugaeva, E.N.; Vasiliev, D.N.; Kabanova, A.P.; Barannik, A.P.; Shneider, M.M.; Kulikov, E.E.; Korzhenkov, A.A.; Toschakov, S.V.; Ignatov, A.N.; et al. Characterization of Pectobacterium carotovorum subsp. carotovorum bacteriophage PP16 prospective for biocontrol of potato soft rot. Microbiology 2019, 88, 451–460. [Google Scholar] [CrossRef]
- Zaczek-Moczydłowska, M.A.; Young, G.K.; Trudgett, J.; Plahe, C.; Fleming, C.C.; Campbell, K.; O’Hanlon, R. Phage cocktail containing Podoviridae and Myoviridae bacteriophages inhibits the growth of Pectobacterium spp. under in vitro and in vivo conditions. PLoS ONE 2020, 15, e0230842. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, E.M.; van Vaerenbergh, J.; Vandenheuvel, D.; Dunon, V.; Ceyssens, P.J.; de Proft, M.; Kropinski, A.M.; Noben, J.P.; Maes, M.; Lavigne, R. T4-related bacteriophage LIMEstone isolates for the control of soft rot on potato caused by “Dickeya solani”. PLoS ONE 2012, 7, e33227. [Google Scholar] [CrossRef] [Green Version]
- Czajkowski, R.; Smolarska, A.; Ozymko, Z. The viability of lytic bacteriophage ΦD5 in potato-associated environments and its effect on Dickeya solani in potato (Solanum tuberosum L.) plants. PLoS ONE 2017, 12, e0183200. [Google Scholar] [CrossRef] [Green Version]
- Hélias, V.; Hamon, P.; Huchet, E.; Wolf, J.V.D.D.; Andrivon, D. Two new effective semiselective crystal violet pectate media for isolation of Pectobacterium and Dickeya. Plant Pathol. 2012, 61, 339–345. [Google Scholar] [CrossRef]
- Humphris, S.N.; Cahill, G.; Elphinstone, J.G.; Kelly, R.; Parkinson, N.M.; Pritchard, L.; Toth, I.K.; Saddler, G.S. Detection of the bacterial potato pathogens Pectobacterium and Dickeya spp. Using Conventional and Real-Time PCR. Methods Mol. Biol. 2015, 1302, 1–16. [Google Scholar] [CrossRef]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Isolation and characterization of novel soilborne lytic bacteriophages infecting Dickeya spp. biovar 3 (‘D. solani’). Plant Pathol. 2014, 63, 758–772. [Google Scholar] [CrossRef]
- Evseev, P.V.; Lukianova, A.A.; Shneider, M.M.; Korzhenkov, A.A.; Bugaeva, E.N.; Kabanova, A.P.; Miroshnikov, K.K.; Kulikov, E.E.; Toshchakov, S.V.; Ignatov, A.N.; et al. Origin and evolution of studiervirinae bacteriophages infecting Pectobacterium: Horizontal transfer assists adaptation to new niches. Microorganisms 2020, 8, 1707. [Google Scholar] [CrossRef]
- Lukianova, A.A.; Shneider, M.M.; Evseev, P.V.; Shpirt, A.M.; Bugaeva, E.N.; Kabanova, A.P.; Obraztsova, E.A.; Miroshnikov, K.K.; Senchenkova, S.N.; Shashkov, A.S.; et al. Morphologically different Pectobacterium brasiliense bacteriophages PP99 and PP101: Deacetylation of O-polysaccharide by the tail spike protein of phage PP99 accompanies the infection. Front. Microbiol. 2020, 10, 3147. [Google Scholar] [CrossRef] [Green Version]
- Clokie, M.R.J.; Kropinski, A.M. (Eds.) Bacteriophages Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Humana Press Inc.: New York, NY, USA, 2009; ISBN 978-1-58829-682-5. [Google Scholar]
- Colombet, J.; Robin, A.; Lavie, L.; Bettarel, Y.; Cauchie, H.M.; Sime-Ngando, T. Virioplankton “pegylation”: Use of PEG (polyethylene glycol) to concentrate and purify viruses in pelagic ecosystems. J. Microbiol. Methods 2007, 71, 212–219. [Google Scholar] [CrossRef] [PubMed]
- Shirshikov, F.V.; Korzhenkov, A.A.; Miroshnikov, K.K.; Kabanova, A.P.; Barannik, A.P.; Ignatov, A.N.; Miroshnikov, K.A. Draft genome sequences of new genomospecies “Candidatus Pectobacterium maceratum” strains, which cause soft rot in plants. Genome Announc. 2018, 6, e00260-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portier, P.; Pédron, J.; Taghouti, G.; Fischer-Le Saux, M.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef]
- Dees, M.W.; Lebecka, R.; Perminow, J.I.S.; Czajkowski, R.; Grupa, A.; Motyka, A.; Zoledowska, S.; Śliwka, J.; Lojkowska, E.; Brurberg, M.B. Characterization of Dickeya and Pectobacterium strains obtained from diseased potato plants in different climatic conditions of Norway and Poland. Eur. J. Plant Pathol. 2017, 148, 839–851. [Google Scholar] [CrossRef]
- Malko, A.; Frantsuzov, P.; Nikitin, M.; Statsyuk, N.; Dzhavakhiya, V.; Golikov, A. Potato Pathogens in Russia’s Regions: An Instrumental survey with the use of Real-Time PCR/RT-PCR in Matrix Format. Pathogens 2019, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Motyka-Pomagruk, A.; Zoledowska, S.; Sledz, W.; Lojkowska, E. The occurrence of bacteria from different species of Pectobacteriaceae on seed potato plantations in Poland. Eur. J. Plant Pathol. 2020. [Google Scholar] [CrossRef]
- Ngoc Ha, V.T.; Voronina, M.V.; Kabanova, A.P.; Shneider, M.M.; Korzhenkov, A.A.; Toschakov, S.V.; Miroshnikov, K.K.; Miroshnikov, K.A.; Ignatov, A.N. First report of Pectobacterium parmentieri causing stem rot disease of potato in Russia. Plant Dis. 2019, 103, 144. [Google Scholar] [CrossRef]
- Zoledowska, S.; Motyka, A.; Zukowska, D.; Sledz, W.; Lojkowska, E. Population structure and biodiversity of Pectobacterium parmentieri isolated from potato fields in temperate climate. Plant Dis. 2018. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.; Abedon, S.T. Phage therapy pharmacology. Phage cocktails. Adv. Appl. Microbiol. 2012, 78, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Tsonos, J.; Vandenheuvel, D.; Briers, Y.; De Greve, H.; Hernalsteens, J.P.; Lavigne, R. Hurdles in bacteriophage therapy: Deconstructing the parameters. Vet. Microbiol. 2014, 171, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.J.; Trauner, A.; Komitopoulou, E.; Salmond, G.P.C. Exploitation of a new flagellatropic phage of Erwinia for positive selection of bacterial mutants attenuated in plant virulence: Towards phage therapy. J. Appl. Microbiol. 2010, 108, 676–685. [Google Scholar] [CrossRef]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [Green Version]
- Dees, M.W.; Lysøe, E.; Rossmann, S.; Perminow, J.; Brurberg, M.B. Pectobacterium polaris sp. nov., isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2017, 67, 5222–5229. [Google Scholar] [CrossRef]
- Staicu, L.C.; Ackerson, C.J.; Cornelis, P.; Ye, L.; Berendsen, R.L.; Hunter, W.J.; Noblitt, S.D.; Henry, C.S.; Cappa, J.J.; Montenieri, R.L.; et al. Pseudomonas moraviensis subsp. stanleyae, a bacterial endophyte of hyperaccumulator Stanleya pinnata, is capable of efficient selenite reduction to elemental selenium under aerobic conditions. J. Appl. Microbiol. 2015, 119, 400–410. [Google Scholar] [CrossRef]
- Niu, B.; Paulson, J.N.; Zheng, X.; Kolter, R. Simplified and representative bacterial community of maize roots. Proc. Natl. Acad. Sci. USA 2017, 114, E2450–E2459. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Potato cv | CVP | Maceration | Species | PP16 | PP47 | PP101 | Q50 | Possum |
|---|---|---|---|---|---|---|---|---|---|
| 2_1 | Bellarosa | + 1 | + | Pve | + | + | − | − | − |
| 2_2 | Bellarosa | + | + | Pod | − | − | − | − | − |
| 2_3 | Bellarosa | + | + | Pve | + | + | − | − | − |
| 2_4 | Bellarosa | + | + | Pve | + | + | − | − | − |
| 2_5 | Bellarosa | + | + | Pve | − | + | + | + | − |
| 4 | Lady Claire | + | + | Pbr | + | − | + | + | − |
| 4_82 | Gala | + | + | Pbr | − | + | + | + | − |
| 5_1 | Gala | + | + | Pbr | − | + | + | + | − |
| 5_2 | Gala | + | + | Pbr | − | + | + | + | − |
| 6_1 | Arsenal | + | + | Pve | + | + | + | − | − |
| 6_2 | Arsenal | + | + | Pve | + | + | + | − | − |
| 8_1 | Isle of Jura | + | + | Pve | + | + | − | + | + |
| 8_2 | Isle of Jura | + | + | Pve | + | + | − | + | + |
| 9_1 | Molly | + | + | Pbr | + | − | + | + | − |
| 9_2 | Molly | + | + | Pbr | + | − | + | + | − |
| 11_1 | Isle of Jura | + | + | Ppo | − | − | − | + | − |
| 11_2 | Isle of Jura | + | + | Pve | − | − | − | − | + |
| 19 | Gala | + | + | Ppo | − | − | − | + | − |
| 20 | Gala | + | + | Ppo | − | − | − | + | − |
| Phage | Morphology 1 | Genus | Propagation Strain 2 | Infection Range | Genome NCBI Accession Number | Genome, bp | Reference |
|---|---|---|---|---|---|---|---|
| PP16 | P | Kotilavirus | Pve F002 | Pve + Pbr | NC_031068 | 44,268 | [8] |
| PP47 | P | Pektosvirus | Pbr F157 | Pbr + Pve | NC_047801 | 40,844 | [15] |
| PP101 | M | Suwonvirus | Pbr F152 | Pbr + Pve | NC_047791 | 53,333 | [16] |
| Q51 | M | Kolesnikvirus | Pve F018 | Ppo + Pve + Pbr | MK_053931 | 84,303 | This work |
| Possum | P | Enquatrovirus | Pve F131 | Pat + Pve | MN_812691 | 73,090 | This work |
| Isolate | CVP Pit Formation | Maceration of Potato Slices | Species |
|---|---|---|---|
| 7211 | + | + | P. carotovorum |
| 7231 | + | + | Pantoea agglomerans |
| 7232 | + | + | P. polaris |
| 7233 | + | + | P. polaris |
| 7241 | + | + | P. polaris |
| 7242 | + | + | P. polaris |
| 7243 | + | + | P. carotovorum |
| 7413 | + | + | Chryseobacterium sp. |
| 7522 | + | + | P. carotovorum |
| 7523 | + | + | Pseudomonas moraviensis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bugaeva, E.N.; Voronina, M.V.; Vasiliev, D.M.; Lukianova, A.A.; Landyshev, N.N.; Ignatov, A.N.; Miroshnikov, K.A. Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study. Viruses 2021, 13, 1095. https://doi.org/10.3390/v13061095
Bugaeva EN, Voronina MV, Vasiliev DM, Lukianova AA, Landyshev NN, Ignatov AN, Miroshnikov KA. Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study. Viruses. 2021; 13(6):1095. https://doi.org/10.3390/v13061095
Chicago/Turabian StyleBugaeva, Eugenia N., Maya V. Voronina, Dmitry M. Vasiliev, Anna A. Lukianova, Nikolay N. Landyshev, Alexander N. Ignatov, and Konstantin A. Miroshnikov. 2021. "Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study" Viruses 13, no. 6: 1095. https://doi.org/10.3390/v13061095
APA StyleBugaeva, E. N., Voronina, M. V., Vasiliev, D. M., Lukianova, A. A., Landyshev, N. N., Ignatov, A. N., & Miroshnikov, K. A. (2021). Use of a Specific Phage Cocktail for Soft Rot Control on Ware Potatoes: A Case Study. Viruses, 13(6), 1095. https://doi.org/10.3390/v13061095