
Hepatitis C Virus Subtypes Novel 6g-Related Subtype and 6w Could Be Indigenous in Southern Taiwan with Characteristic Geographic Distribution

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Diagnostic Procedures
Study Design
2.3. Genotyping by Abbott GT II and PLUS Assays
2.4. Genotyping by 5′UTR/Core Sequencing
2.5. Subtyping by E1 and NS5B Sequencing
2.6. Phylogenetic Analysis by E1 and NS5B Sequencing
3. Results
4. Discussions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Childs, K.; Davis, C.; Cannon, M.; Montague, S.; Filipe, A.; Tong, L.; Simmonds, P.; Smith, D.; Thomson, E.C.; Dusheiko, G.; et al. Suboptimal SVR rates in African patients with atypical genotype 1 subtypes: Implications for global elimination of hepatitis C. J. Hepatol. 2019, 71, 1099–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, D.T.; Tran, T.T.T.; Nghiem, N.M.; Le, P.T.; Vo, Q.M.; Day, J.; Rahman, M.; Le, H.M. Effectiveness of sofosbuvir based direct-acting antiviral regimens for chronic hepatitis C virus genotype 6 patients: Real-world experience in Vietnam. PLoS ONE 2020, 15, e0233446. [Google Scholar] [CrossRef]
- Chen, J.J.; Lee, P.L.; Chiu, H.C.; Tung, H.D.; Chiu, Y.C.; Cheng, P.N. Real-world effectiveness and safety of ledipasvir/sofosbuvir for genotype 6 chronic hepatitis C patients in Taiwan. J. Gastroenterol. Hepatol. 2020, 35, 467–472. [Google Scholar] [CrossRef]
- Mettikanont, P.; Bunchorntavakul, C.; Reddy, K.R. Systematic review: Epidemiology and response to direct-acting antiviral therapy in genotype 6 chronic hepatitis C virus infection. Aliment. Pharmacol. Ther. 2019, 49, 492–505. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Davidson, F.; Simmonds, P. Hepatitis C virus variants and the role of genotyping. J. Hepatol. 1995, 23 (Suppl. 2), 26–31. [Google Scholar]
- Pybus, O.G.; Barnes, E.; Taggart, R.; Lemey, P.; Markov, P.V.; Rasachak, B.; Syhavong, B.; Phetsouvanah, R.; Sheridan, I.; Humphreys, I.S.; et al. Genetic history of hepatitis C virus in East Asia. J. Virol. 2009, 83, 1071–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.B.; Bukh, J.; Kuiken, C.; Muerhoff, A.S.; Rice, C.M.; Stapleton, J.T.; Simmonds, P. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: Updated criteria and genotype assignment web resource. Hepatology 2014, 59, 318–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasitthankasem, R.; Vongpunsawad, S.; Siripon, N.; Suya, C.; Chulothok, P.; Chaiear, K.; Rujirojindakul, P.; Kanjana, S.; Theamboonlers, A.; Tangkijvanich, P.; et al. Genotypic distribution of hepatitis C virus in Thailand and Southeast Asia. PLoS ONE 2015, 10, e0126764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thong, V.D.; Akkarathamrongsin, S.; Poovorawan, K.; Tangkijvanich, P.; Poovorawan, Y. Hepatitis C virus genotype 6: Virology, epidemiology, genetic variation and clinical implication. World J. Gastroenterol. 2014, 20, 2927–2940. [Google Scholar] [CrossRef] [PubMed]
- International Committee on Taxonomy of Viruses(ICTV). HCV Classification. Available online: https://talk.ictvonline.org/ictv_wikis/flaviviridae/w/sg_flavi/56/hcv-classification (accessed on 23 April 2021).
- Chen, C.H.; Sheu, J.C.; Wang, J.T.; Huang, G.T.; Yang, P.M.; Lee, H.S.; Lee, C.Z.; Chen, D.S. Genotypes of hepatitis C virus in chronic liver disease in Taiwan. J. Med. Virol. 1994, 44, 234–236. [Google Scholar] [CrossRef]
- Kao, J.H.; Chen, P.J.; Lai, M.Y.; Yang, P.M.; Sheu, J.C.; Wang, T.H.; Chen, D.S. Genotypes of hepatitis C virus in Taiwan and the progression of liver disease. J. Clin. Gastroenterol. 1995, 21, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.; Lu, S.N.; Hung, C.H.; Tung, W.C.; Wang, J.H.; Tung, H.D.; Chen, C.H.; Hu, T.H.; Changchien, C.S.; Chen, W.J. Hepatitis C virus genotypes in southern Taiwan: Prevalence and clinical implications. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 767–774. [Google Scholar] [CrossRef]
- Chen, C.H.; Yang, P.M.; Huang, G.T.; Lee, H.S.; Sung, J.L.; Sheu, J.C. Estimation of seroprevalence of hepatitis B virus and hepatitis C virus in Taiwan from a large-scale survey of free hepatitis screening participants. J. Formos. Med. Assoc. 2007, 106, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Omata, M.; Kanda, T.; Yokosuka, O.; Crawford, D.; Al-Mahtab, M.; Wei, L.; Ibrahim, A.; Lau, G.K.; Sharma, B.C.; Hamid, S.S.; et al. Features of hepatitis C virus infection, current therapies and ongoing clinical trials in ten Asian Pacific countries. Hepatol. Int. 2015, 9, 486–507. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Lin, H.J.; Chen, Y.J.; Lee, C.M.; Wang, S.F.; Chang, K.Y.; Chen, T.L.; Liu, H.F.; Chen, Y.M. Molecular epidemiology of HCV genotypes among injection drug users in Taiwan: Full-length sequences of two new subtype 6w strains and a recombinant form_2b6w. J. Med. Virol. 2010, 82, 57–68. [Google Scholar] [CrossRef]
- Liu, J.Y.; Lin, H.H.; Liu, Y.C.; Lee, S.S.; Chen, Y.L.; Hung, C.C.; Ko, W.C.; Huang, C.K.; Lai, C.H.; Chen, Y.S.; et al. Extremely high prevalence and genetic diversity of hepatitis C virus infection among HIV-infected injection drug users in Taiwan. Clin. Infect. Dis. 2008, 46, 1761–1768. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.J.; Tung, H.D.; Lee, P.L.; Kuo, H.T.; Sheu, M.J.; Cheng, C.T.; Chuang, T.W.; Kao, H.J.; Lu, N.M.; Wu, L.C.; et al. High prevalence of genotype 6 hepatitis C virus infection in Southern Taiwan using Abbott genotype assays. J. Formos. Med. Assoc. 2020, 119, 413–419. [Google Scholar] [CrossRef]
- Tung, H.D.; Lee, P.L.; Chen, J.J.; Kuo, H.T.; Sheu, M.J.; Cheng, C.T.; Chuang, T.W.; Kao, H.J.; Hou, C.Y.; Tsai, H.H.; et al. Geographic variation of genotype 6 hepatitis C virus infection in an endemic area of southern Taiwan. J. Formos. Med. Assoc. 2020, 119, 1876–1880. [Google Scholar] [CrossRef]
- Bukh, J.; Purcell, R.H.; Miller, R.H. Sequence analysis of the 5’ noncoding region of hepatitis C virus. Proc. Natl. Acad. Sci. USA 1992, 89, 4942–4946. [Google Scholar] [CrossRef] [Green Version]
- Pohjanpelto, P.; Lappalainen, M.; Widell, A.; Asikainen, K.; Paunio, M. Hepatitis C genotypes in Finland determined by RFLP. Clin. Diagn. Virol. 1996, 7, 7–16. [Google Scholar] [CrossRef]
- Mellor, J.; Holmes, E.C.; Jarvis, L.M.; Yap, P.L.; Simmonds, P. Investigation of the pattern of hepatitis C virus sequence diversity in different geographical regions: Implications for virus classification. The International HCV Collaborative Study Group. J. Gen. Virol. 1995, 76, 2493–2507. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Yu, M.L.; Chuang, W.L.; Chen, S.C.; Dai, C.Y.; Hou, C.; Wang, J.H.; Lu, S.N.; Huang, J.F.; Lin, Z.Y.; Hsieh, M.Y.; et al. Changing prevalance of hepatitis C virus genotypes: Molicular epidemiology and clinical implications in the hepatitis C virus hyperendemic areas and a tertiary referral center in Taiwan. J. Med. Virol. 2001, 65, 58–65. [Google Scholar] [CrossRef]
- Hedskog, C.; Parhy, B.; Chang, S.; Zeuzem, S.; Moreno, C.; Shafran, S.D.; Borgia, S.M.; Asselah, T.; Alric, L.; Abergel, A.; et al. Identification of 19 Novel Hepatitis C Virus Subtypes-Further Expanding HCV Classification. Open Forum Infect. Dis. 2019, 6, ofz076. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.S.; Lee, H.F.; Hsiau, H.L.; Lu, H.Y.; Chou, W.H.; Lu, C.F.; Chen, H.Y.; Lee, F.N.; Chen, P.Y.; Tam, K.M. Genotype distribution of hepatitis C virus infection in Taiwan. J. Med. Virol. 1994, 44, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Stuyver, L.; Wyseur, A.; van Arnhem, W.; Hernandez, F.; Maertens, G. Second-generation line probe assay for hepatitis C virus genotyping. J. Clin. Microbiol. 1996, 34, 2259–2266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmonds, P.; Bukh, J.; Combet, C.; Deleage, G.; Enomoto, N.; Feinstone, S.; Halfon, P.; Inchauspe, G.; Kuiken, C.; Maertens, G.; et al. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology 2005, 42, 962–973. [Google Scholar] [CrossRef]
- Hraber, P.T.; Fischer, W.; Bruno, W.J.; Leitner, T.; Kuiken, C. Comparative analysis of hepatitis C virus phylogenies from coding and non-coding regions: The 5’ untranslated region (UTR) fails to classify subtypes. Virol. J. 2006, 3, 103. [Google Scholar] [CrossRef] [Green Version]
- Murphy, D.G.; Willems, B.; Deschenes, M.; Hilzenrat, N.; Mousseau, R.; Sabbah, S. Use of sequence analysis of the NS5B region for routine genotyping of hepatitis C virus with reference to C/E1 and 5’ untranslated region sequences. J. Clin. Microbiol. 2007, 45, 1102–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallory, M.A.; Lucic, D.; Ebbert, M.T.; Cloherty, G.A.; Toolsie, D.; Hillyard, D.R. Evaluation of the Abbott RealTime HCV genotype II plus RUO (PLUS) assay with reference to core and NS5B sequencing. J. Clin. Virol. 2017, 90, 26–31. [Google Scholar] [CrossRef]
- Hsieh, M.H.; Tsai, J.J.; Hsieh, M.Y.; Huang, C.F.; Yeh, M.L.; Yang, J.F.; Chang, K.; Lin, W.R.; Lin, C.Y.; Chen, T.C.; et al. Hepatitis C virus infection among injection drug users with and without human immunodeficiency virus co-infection. PLoS ONE 2014, 9, e94791. [Google Scholar] [CrossRef]
- Lee, H.C.; Ko, N.Y.; Lee, N.Y.; Chang, C.M.; Ko, W.C. Seroprevalence of viral hepatitis and sexually transmitted disease among adults with recently diagnosed HIV infection in Southern Taiwan, 2000-2005: Upsurge in hepatitis C virus infections among injection drug users. J. Formos. Med. Assoc. 2008, 107, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.M.; Lan, Y.C.; Lai, S.F.; Yang, J.Y.; Tsai, S.F.; Kuo, S.H. HIV-1 CRF07_BC infections, injecting drug users, Taiwan. Emerg. Infect. Dis. 2006, 12, 703–705. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.H.; Shih, Y.L.; Liu, Y.C.; Lee, S.S.; Huang, C.K.; Chen, Y.L.; Chin, C.; Lai, C.H.; Tsai, H.C.; Guo, Y.C.; et al. An epidemic of HIV type I CRF07_BC infection among injection drug users in Taiwan. J. Acquir. Immune Defic. Syndr. 2006, 42, 248–255. [Google Scholar] [CrossRef]
- Tee, K.K.; Pybus, O.G.; Liao, H.; Uenishi, R.; Hase, S.; Kamarulzaman, A.; Li, X.J.; Takebe, Y. Chronology of the HIV-1 CRF07_BC expansion in East Asia. AIDS 2008, 22, 156–158. [Google Scholar] [CrossRef]
- Chen, Y.M.; Kuo, S.H. HIV-1 in Taiwan. Lancet 2007, 369, 623–625. [Google Scholar] [CrossRef]
- Tee, K.K.; Pybus, O.G.; Li, X.J.; Han, X.; Shang, H.; Kamarulzaman, A.; Takebe, Y. Temporal and spatial dynamics of human immunodeficiency virus type 1 circulating recombinant forms 08_BC and 07_BC in Asia. J. Virol. 2008, 82, 9206–9215. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Fu, Y.; Lu, L.; Ji, W.; Yu, J.; Hagedorn, C.H.; Zhang, L. Complete genomic sequences for hepatitis C virus subtypes 6e and 6g isolated from Chinese patients with injection drug use and HIV-1 co-infection. J. Med. Virol. 2006, 78, 1061–1069. [Google Scholar] [CrossRef]
- Lu, L.; Nakano, T.; Li, C.; Fu, Y.; Miller, S.; Kuiken, C.; Robertson, B.H.; Hagedorn, C.H. Hepatitis C virus complete genome sequences identified from China representing subtypes 6k and 6n and a novel, as yet unassigned subtype within genotype 6. J. Gen. Virol. 2006, 87, 629–634. [Google Scholar] [CrossRef]
- Tokita, H.; Okamoto, H.; Iizuka, H.; Kishimoto, J.; Tsuda, F.; Lesmana, L.A.; Miyakawa, Y.; Mayumi, M. Hepatitis C virus variants from Jakarta, Indonesia classifiable into novel genotypes in the second (2e and 2f), tenth (10a) and eleventh (11a) genetic groups. J. Gen. Virol. 1996, 77, 293–301. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, C.; Yin, X.; Guo, X.; Wu, S.; Hou, J. Hepatitis C virus genotypes and subtypes circulating in Mainland China. Emerg. Microbes Infect. 2017, 6, e95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Y.; Wu, T.; Wang, M.; Lu, L.; Li, C.; Zhou, Y.; Fu, Y.; Chen, G. Conservation in China of a novel group of HCV variants dating to six centuries ago. Virology 2014, 464-465, 21–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Xing, Z.; Yuan, M.; Ge, J.; Yuan, G.; Liang, K.; Wu, B.; Xiao, F.; Li, C.; Zhou, Y.; et al. Analysis of HCV Isolates Among the Li Ethnic in Hainan Island of South China Reveals Their HCV-6 Unique Evolution and a New Subtype. Cell Physiol. Biochem. 2018, 50, 1832–1839. [Google Scholar] [CrossRef]
- Wu, T.; Xiong, L.; Wang, F.; Xu, X.; Wang, J.; Lin, F.; Li, C.; Lu, L.; Zhou, Y. A Unique Pattern of HCV Genotype Distribution on Hainan Island in China Revealed by Evolutionary Analysis. Cell Physiol. Biochem. 2016, 39, 316–330. [Google Scholar] [CrossRef]
- Li, C.; Barnes, E.; Newton, P.N.; Fu, Y.; Vongsouvath, M.; Klenerman, P.; Okamoto, H.; Abe, K.; Pybus, O.G.; Lu, L. An expanded taxonomy of hepatitis C virus genotype 6: Characterization of 22 new full-length viral genomes. Virology 2015, 476, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Chao, D.T.; Abe, K.; Nguyen, M.H. Systematic review: Epidemiology of hepatitis C genotype 6 and its management. Aliment. Pharmacol. Ther. 2011, 34, 286–296. [Google Scholar] [CrossRef]
- Lu, S.N.; Su, W.W.; Yang, S.S.; Chang, T.T.; Cheng, K.S.; Wu, J.C.; Lin, H.H.; Wu, S.S.; Lee, C.M.; Changchien, C.S.; et al. Secular trends and geographic variations of hepatitis B virus and hepatitis C virus-associated hepatocellular carcinoma in Taiwan. Int. J. Cancer 2006, 119, 1946–1952. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | Districts | GT1a | GT1b | GT2 | GT3 | GT4 | GT6 | MT | T (n) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Baihe | 0 | 6 | 5 | 0 | 0 | 5 | 0 | 16 |
| Dongshan | 1 | 21 | 36 | 0 | 0 | 21 | 1 | 80 | |
| Liouying | 4 | 53 | 75 | 0 | 0 | 65 | 1 | 198 | |
| Liujia | 2 | 30 | 70 | 1 | 0 | 67 | 1 | 171 | |
| Xiaying | 3 | 82 | 83 | 2 | 0 | 65 | 3 | 238 | |
| Guantian | 5 | 31 | 48 | 1 | 1 | 41 | 1 | 128 | |
| Danei | 1 | 5 | 7 | 1 | 0 | 2 | 0 | 16 | |
| 2 | Xuejia | 2 | 27 | 20 | 0 | 0 | 3 | 0 | 52 |
| Madou | 12 | 25 | 66 | 0 | 1 | 23 | 1 | 128 | |
| Jiangjun | 2 | 19 | 82 | 0 | 0 | 24 | 1 | 128 | |
| Chiali | 6 | 45 | 63 | 0 | 1 | 22 | 0 | 137 | |
| Xigang | 1 | 17 | 22 | 0 | 0 | 7 | 0 | 47 | |
| Qigu | 0 | 26 | 38 | 0 | 0 | 7 | 0 | 71 | |
| 3 | Nanxi | 0 | 5 | 6 | 0 | 0 | 1 | 0 | 12 |
| Yujing | 1 | 9 | 13 | 0 | 0 | 1 | 0 | 24 | |
| Nanhua | 1 | 3 | 3 | 0 | 0 | 0 | 0 | 7 | |
| Shanshang | 1 | 1 | 5 | 0 | 0 | 0 | 0 | 7 | |
| Zuozhen | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 2 | |
| Shanhua | 6 | 12 | 20 | 0 | 0 | 11 | 0 | 49 | |
| Xinshi | 2 | 16 | 23 | 1 | 0 | 5 | 1 | 48 | |
| Xinhua | 0 | 28 | 49 | 0 | 1 | 9 | 1 | 88 | |
| Guanmiao | 1 | 5 | 10 | 0 | 0 | 3 | 0 | 19 | |
| Anding | 0 | 4 | 7 | 0 | 0 | 3 | 0 | 14 | |
| Yongkang | 10 | 75 | 116 | 7 | 0 | 22 | 1 | 231 | |
| Guiren | 3 | 11 | 24 | 0 | 0 | 2 | 0 | 40 | |
| Annan | 8 | 42 | 69 | 1 | 1 | 20 | 0 | 141 | |
| Anping | 2 | 27 | 51 | 0 | 1 | 3 | 0 | 84 | |
| North | 6 | 35 | 36 | 1 | 0 | 16 | 0 | 94 | |
| East | 2 | 23 | 33 | 2 | 0 | 8 | 0 | 68 | |
| W. central | 1 | 16 | 21 | 0 | 0 | 4 | 0 | 42 | |
| South | 3 | 13 | 35 | 1 | 1 | 2 | 0 | 55 | |
| Rende | 3 | 8 | 9 | 0 | 0 | 2 | 1 | 23 | |
| 4 | Houbi | 16 | 44 | 39 | 0 | 0 | 1 | 0 | 100 |
| Xinying | 12 | 146 | 132 | 1 | 0 | 43 | 1 | 335 | |
| Yanshui | 2 | 44 | 57 | 0 | 0 | 10 | 1 | 114 | |
| Beimen | 0 | 2 | 13 | 0 | 0 | 0 | 0 | 15 | |
| Total | 119 | 956 | 1388 | 19 | 7 | 518 | 15 | 3022 |
| Region-1 | Region-2 | Region-3 | Region-4 | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | n | % | |
| GT 1a | 16 | 1.9% | 23 | 4.1% | 50 | 4.8% | 30 | 5.3% | 119 | 3.9% |
| GT 1b | 228 | 26.9% | 159 | 28.2% | 333 | 31.8% | 236 | 41.8% | 956 | 31.6% |
| GT 2 | 324 | 38.3% | 291 | 51.7% | 532 | 50.8% | 241 | 42.7% | 1388 | 45.9% |
| GT 3 | 5 | 0.6% | 0 | 13 | 1.2% | 1 | 0.2% | 19 | 0.6% | |
| GT 4 | 1 | 0.1% | 2 | 0.4% | 4 | 0.4% | 0 | 7 | 0.2% | |
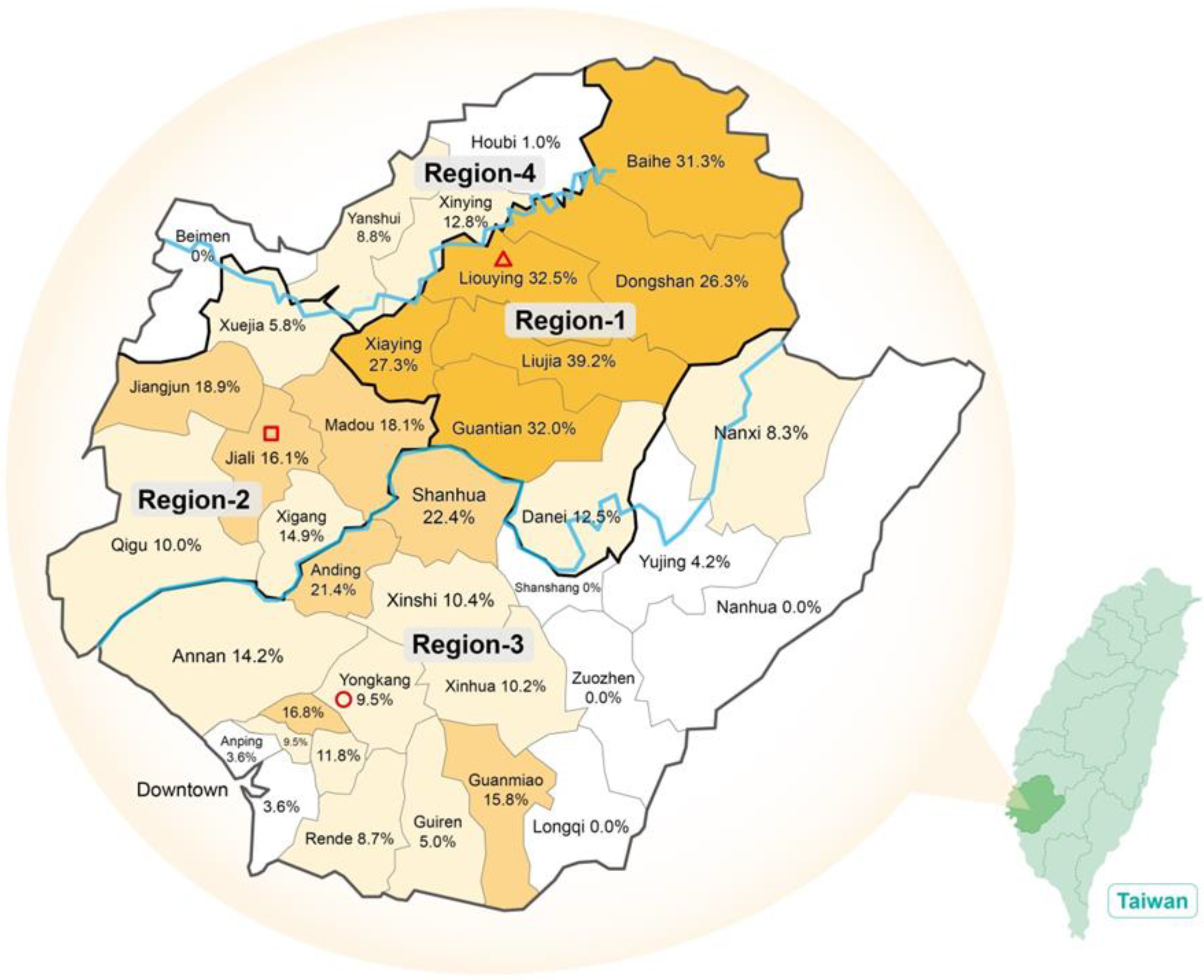
| GT 6 | 266 | 31.4% | 86 | 15.3% | 112 | 10.7% | 54 | 9.6% | 518 | 17.1% |
| Mixed GT | 7 | 0.8% | 2 | 0.4% | 4 | 0.4% | 2 | 0.4% | 15 | 0.5% |
| Total | 847 | 28.0% | 563 | 18.6% | 1048 | 34.7% | 564 | 18.7% | 3022 | |
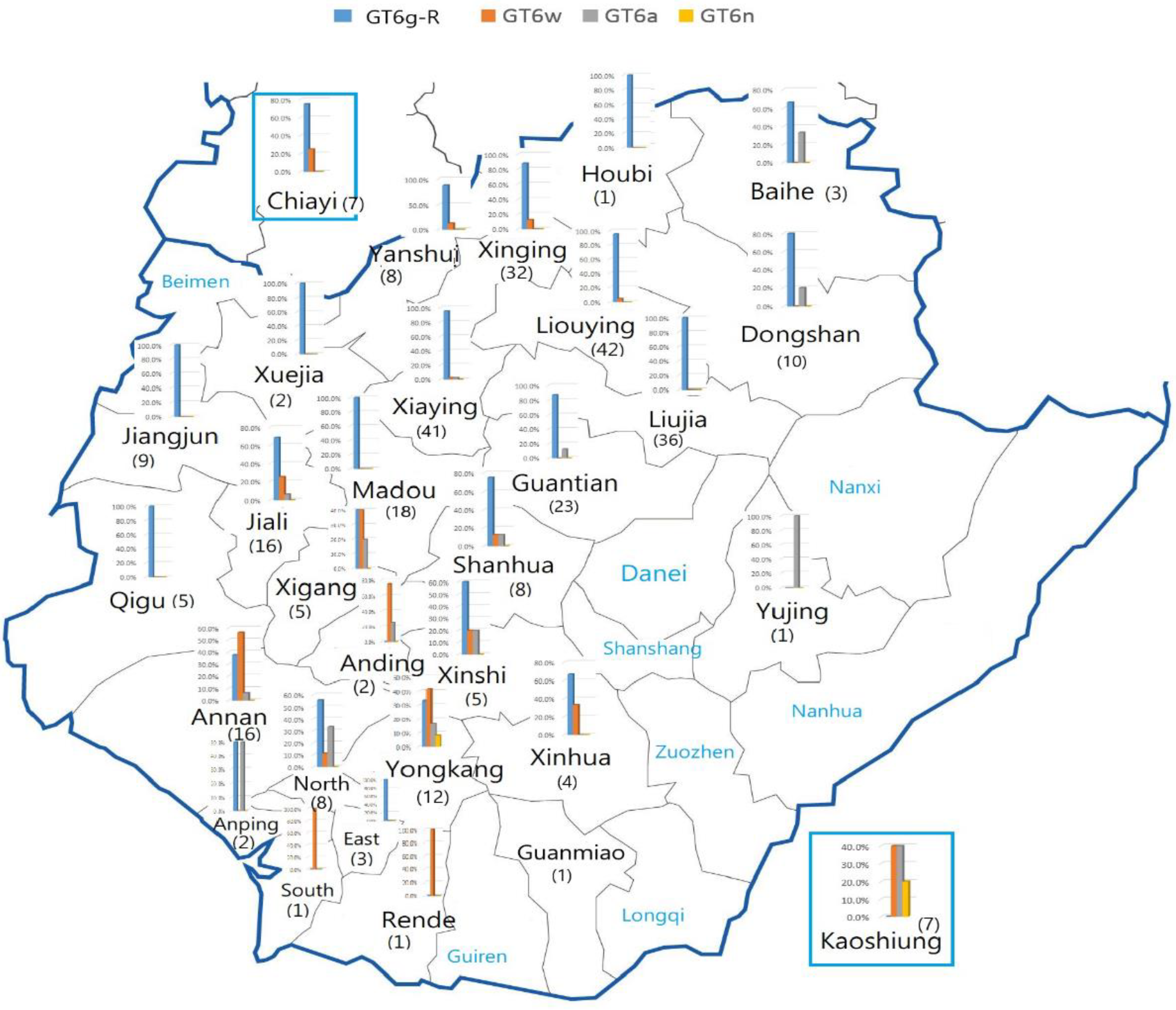
| Region | Districts | GT6a | % | GT6g | % | GT6n | % | GT6w | % | ID | % | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Baihe | 1 | 33.3% | 2 | 66.7% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 3 |
| Dongshan | 1 | 10.0% | 4 | 40.0% | 0 | 0.0% | 0 | 0.0% | 5 | 50.0% | 10 | |
| Liouying | 0 | 0.0% | 38 | 90.5% | 0 | 0.0% | 2 | 4.8% | 2 | 4.8% | 42 | |
| Liujia | 0 | 0.0% | 34 | 94.4% | 0 | 0.0% | 0 | 0.0% | 2 | 5.6% | 36 | |
| Xiaying | 1 | 2.4% | 39 | 95.1% | 0 | 0.0% | 1 | 2.4% | 0 | 0.0% | 41 | |
| Guantian | 3 | 13.0% | 19 | 82.6% | 0 | 0.0% | 0 | 0.0% | 1 | 4.3% | 23 | |
| 6 | 3.9% | 136 | 87.7% | 0 | 0.0% | 3 | 1.9% | 10 | 6.5% | 155 | ||
| 2 | Xuejia | 0 | 0.0% | 2 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 2 |
| Madou | 0 | 0.0% | 18 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 18 | |
| Jiangjun | 0 | 0.0% | 9 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 9 | |
| Jiali | 0 | 0.0% | 11 | 68.8% | 1 | 6.3% | 4 | 25.0% | 0 | 0.0% | 16 | |
| Xigang | 1 | 20.0% | 2 | 40.0% | 0 | 0.0% | 2 | 40.0% | 0 | 0.0% | 5 | |
| Qigu | 0 | 0.0% | 4 | 80.0% | 0 | 0.0% | 0 | 0.0% | 1 | 20.0% | 5 | |
| 1 | 1.8% | 46 | 83.6% | 1 | 1.8% | 6 | 10.9% | 1 | 1.8% | 55 | ||
| 3 | Yujing | 1 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 1 |
| Shanhua | 1 | 12.5% | 6 | 75.0% | 0 | 0.0% | 1 | 12.5% | 0 | 0.0% | 8 | |
| Xinhua | 0 | 0.0% | 2 | 50.0% | 0 | 0.0% | 1 | 25.0% | 1 | 25.0% | 4 | |
| Xinshi | 1 | 20.0% | 3 | 60.0% | 0 | 0.0% | 1 | 20.0% | 0 | 0.0% | 5 | |
| Guanmiao | 1 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 1 | |
| Yongkang | 2 | 16.7% | 3 | 25.0% | 1 | 8.3% | 4 | 33.3% | 2 | 16.7% | 12 | |
| Anding | 1 | 50.0% | 0 | 0.0% | 0 | 0.0% | 1 | 50.0% | 0 | 0.0% | 2 | |
| Annan | 1 | 6.3% | 5 | 31.3% | 0 | 0.0% | 9 | 56.3% | 1 | 6.3% | 16 | |
| Anping | 1 | 50.0% | 1 | 50.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 2 | |
| North | 2 | 25.0% | 5 | 62.5% | 0 | 0.0% | 1 | 12.5% | 0 | 0.0% | 8 | |
| East | 0 | 0.0% | 3 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 3 | |
| W. central | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 2 | 100.0% | 2 | |
| South | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 1 | 100.0% | 0 | 0.0% | 1 | |
| Rende | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 1 | 100.0% | 0 | 0.0% | 1 | |
| 11 | 16.7% | 28 | 42.4% | 1 | 1.5% | 20 | 30.3% | 6 | 9.1% | 66 | ||
| 4 | Houbi | 0 | 0.0% | 1 | 100.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 1 |
| Xinying | 4 | 12.5% | 22 | 68.8% | 0 | 0.0% | 2 | 6.3% | 4 | 12.5% | 32 | |
| Yanshui | 0 | 0.0% | 6 | 75.0% | 0 | 0.0% | 1 | 12.5% | 1 | 12.5% | 8 | |
| 4 | 9.8% | 29 | 70.7% | 0 | 0.0% | 3 | 7.3% | 5 | 12.2% | 41 | ||
| Chiayi | 0 | 0.0% | 6 | 85.7% | 0 | 0.0% | 1 | 14.3% | 0 | 0.0% | 7 | |
| Kaohsiung | 2 | 28.6% | 0 | 0.0% | 1 | 14.3% | 2 | 28.6% | 2 | 28.6% | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tung, H.-D.; Lee, P.-L.; Chen, J.-J.; Kuo, H.-T.; Sheu, M.-J.; Cheng, C.-T.; Chuang, T.-W.; Kao, H.-J.; Wu, Y.-H.; Pang, M.-G.; et al. Hepatitis C Virus Subtypes Novel 6g-Related Subtype and 6w Could Be Indigenous in Southern Taiwan with Characteristic Geographic Distribution. Viruses 2021, 13, 1316. https://doi.org/10.3390/v13071316
Tung H-D, Lee P-L, Chen J-J, Kuo H-T, Sheu M-J, Cheng C-T, Chuang T-W, Kao H-J, Wu Y-H, Pang M-G, et al. Hepatitis C Virus Subtypes Novel 6g-Related Subtype and 6w Could Be Indigenous in Southern Taiwan with Characteristic Geographic Distribution. Viruses. 2021; 13(7):1316. https://doi.org/10.3390/v13071316
Chicago/Turabian StyleTung, Hung-Da, Pei-Lun Lee, Jyh-Jou Chen, Hsing-Tao Kuo, Ming-Jen Sheu, Chun-Ta Cheng, Tang-Wei Chuang, Hsu-Ju Kao, Yu-Hsun Wu, Mai-Gio Pang, and et al. 2021. "Hepatitis C Virus Subtypes Novel 6g-Related Subtype and 6w Could Be Indigenous in Southern Taiwan with Characteristic Geographic Distribution" Viruses 13, no. 7: 1316. https://doi.org/10.3390/v13071316
APA StyleTung, H.-D., Lee, P.-L., Chen, J.-J., Kuo, H.-T., Sheu, M.-J., Cheng, C.-T., Chuang, T.-W., Kao, H.-J., Wu, Y.-H., Pang, M.-G., Lin, C.-H., Hou, C.-Y., Tsai, H.-H., Wu, L.-C., & Lee, C. (2021). Hepatitis C Virus Subtypes Novel 6g-Related Subtype and 6w Could Be Indigenous in Southern Taiwan with Characteristic Geographic Distribution. Viruses, 13(7), 1316. https://doi.org/10.3390/v13071316