
The Requirement of Glycoprotein C for Interindividual Spread Is Functionally Conserved within the Alphaherpesvirus Genus (Mardivirus), but Not the Host (Gallid)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Cells
2.2. Generation of Recombinant (r)MDVs
2.3. Measurement of Plaque Areas
2.4. Viral Replication Kinetics in Cell Culture
2.5. Animal Experiments
2.6. Viral Replication Kinetics In Vivo
2.7. Monitoring rMDVs in Feather Follicles (FFs)
2.8. Immunofluorescence Assay (IFA) of FFs
2.9. Statistical Analyses
3. Results
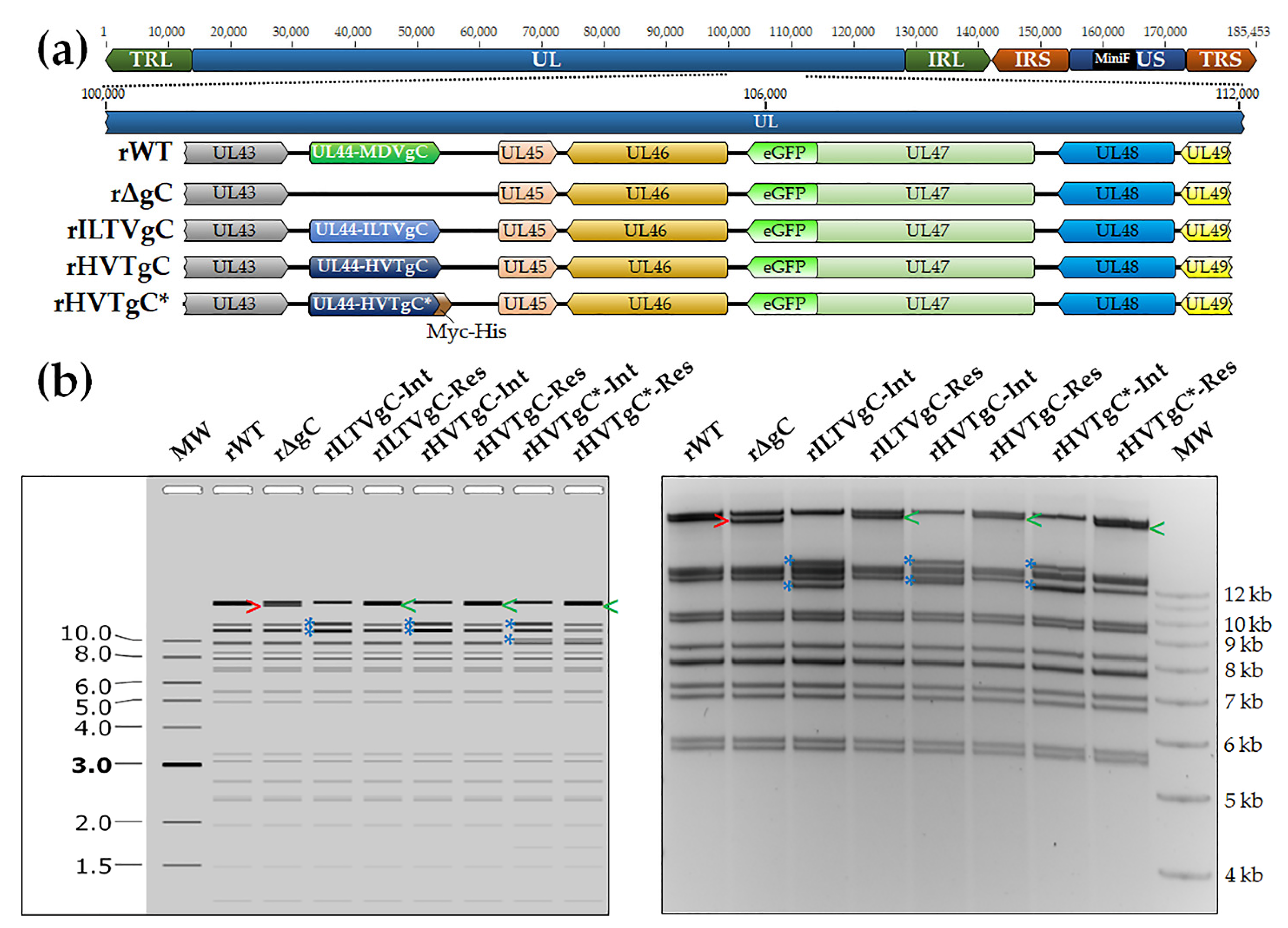
3.1. Generation and Characterization of rMDVs
3.1.1. Generation of rMDV Expressing ILTV or HVT gC
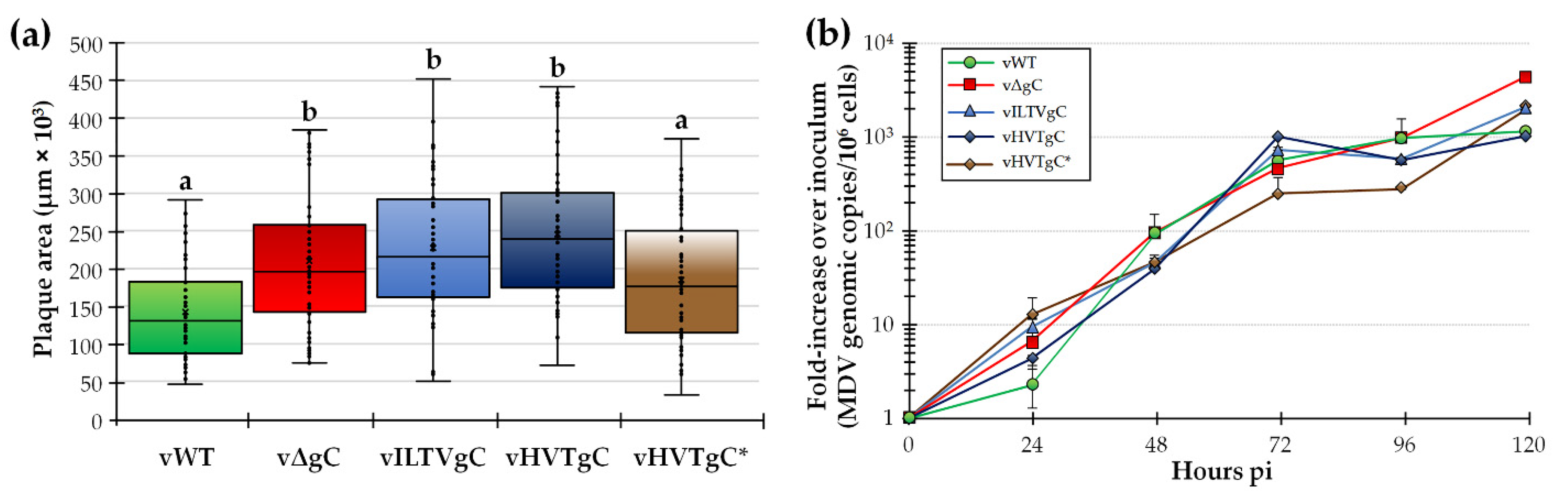
3.1.2. Replication of rMDV Expressing ILTV or HVT gC in Cell Culture
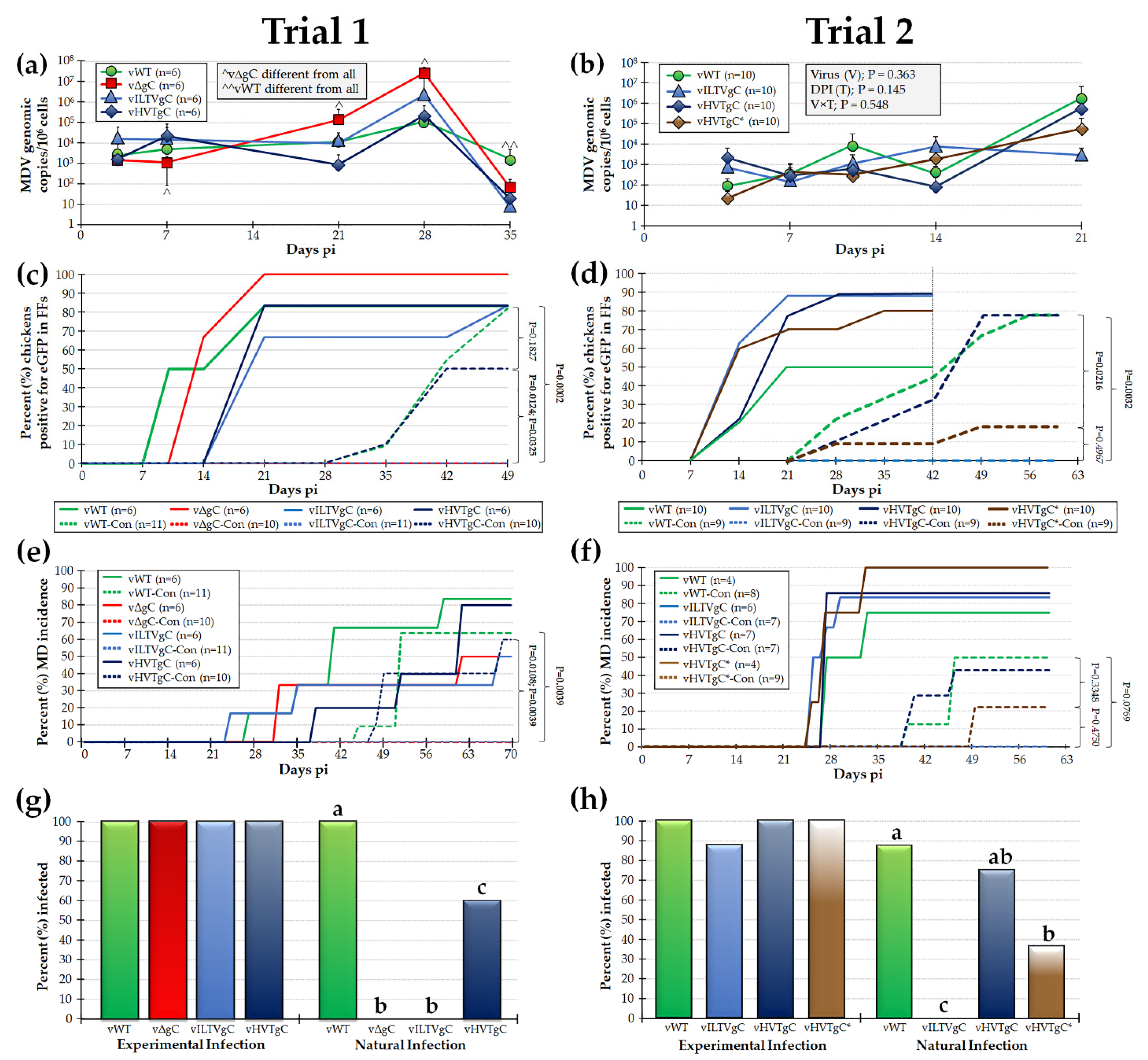
3.2. Characterization of rMDVs in Experimentally Infected Chickens
3.2.1. Replication of rMDVs during Experimental Infection
3.2.2. MD Induction of rMDVs during Experimental Infection
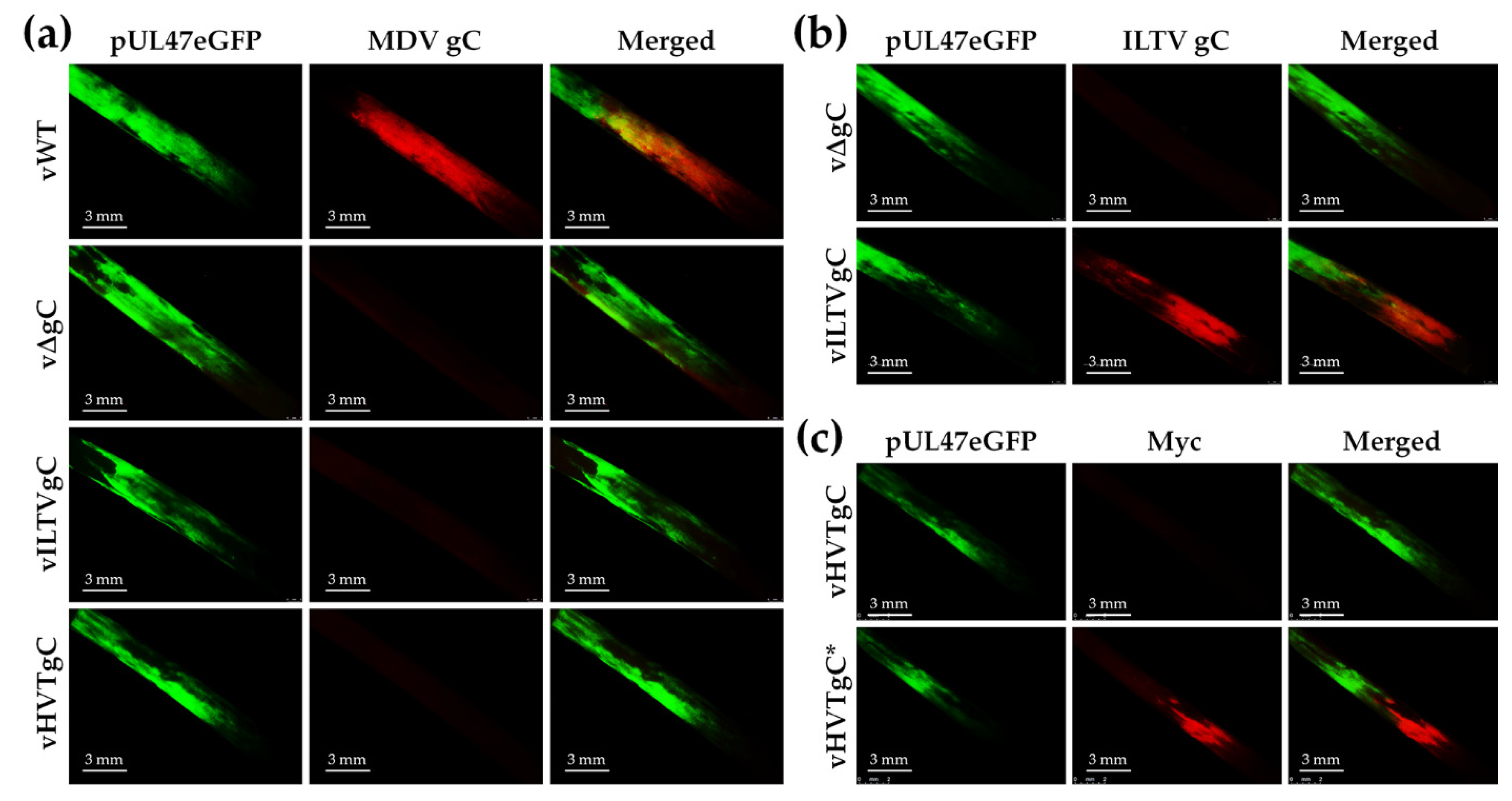
3.2.3. Expression of gC proteins in FFs
3.3. gC Proteins Are Functionally Conserved among the Mardivirus and Not the Gallid Genera
Natural Infection with rMDVs Expressing ILTV or HVT gC
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGeoch, D.J.; Rixon, F.J.; Davison, A.J. Topics in herpesvirus genomics and evolution. Virus Res. 2006, 117, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; McGeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef]
- Gowthaman, V.; Kumar, S.; Koul, M.; Dave, U.; Murthy, T.; Munuswamy, P.; Tiwari, R.; Karthik, K.; Dhama, K.; Michalak, I.; et al. Infectious laryngotracheitis: Etiology, epidemiology, pathobiology, and advances in diagnosis and control—A comprehensive review. Vet. Q. 2020, 40, 140–161. [Google Scholar] [CrossRef] [Green Version]
- Jarosinski, K.W.; Margulis, N.G.; Kamil, J.P.; Spatz, S.J.; Nair, V.K.; Osterrieder, N. Horizontal transmission of Marek’s disease virus requires US2, the UL13 protein kinase, and gC. J. Virol. 2007, 81, 10575–10587. [Google Scholar] [CrossRef] [Green Version]
- Jarosinski, K.W.; Osterrieder, N. Further analysis of Marek’s disease virus horizontal transmission confirms that U(L)44 (gC) and U(L)13 protein kinase activity are essential, while U(S)2 is nonessential. J. Virol. 2010, 84, 7911–7916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarosinski, K.W.; Osterrieder, N. Marek’s disease virus expresses multiple UL44 (gC) variants through mRNA splicing that are all required for efficient horizontal transmission. J. Virol. 2012, 86, 7896–7906. [Google Scholar] [CrossRef] [Green Version]
- Vega-Rodriguez, W.; Ponnuraj, N.; Jarosinski, K.W. Marek’s disease alphaherpesvirus (MDV) RLORF4 is not required for expression of glycoprotein C and interindividual spread. Virology 2019, 534, 108–113. [Google Scholar] [CrossRef]
- Vega-Rodriguez, W.; Xu, H.; Ponnuraj, N.; Akbar, H.; Kim, T.; Jarosinski, K.W. The requirement of glycoprotein C (gC) for interindividual spread is a conserved function of gC for avian herpesviruses. Sci. Rep. 2021, 11, 7753. [Google Scholar] [CrossRef] [PubMed]
- Pavlova, S.P.; Veits, J.; Blohm, U.; Maresch, C.; Mettenleiter, T.C.; Fuchs, W. In vitro and in vivo characterization of glycoprotein C-deleted infectious laryngotracheitis virus. J. Gen. Virol. 2010, 91 Pt 4, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Bulow, V.V.; Biggs, P.M. Differentiation between strains of Marek’s disease virus and turkey herpesvirus by immunofluorescence assays. Avian Pathol. 1975, 4, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Churchill, A.E.; Chubb, R.C.; Baxendale, W. The attenuation, with loss of oncogenicity, of the herpes-type virus of Marek’s disease (strain HPRS-16) on passage in cell culture. J. Gen. Virol. 1969, 4, 557–564. [Google Scholar] [CrossRef]
- Ikuta, K.; Ueda, S.; Kato, S.; Hirai, K. Monoclonal antibodies reactive with the surface and secreted glycoproteins of Marek’s disease virus and herpesvirus of turkeys. J. Gen. Virol. 1983, 64 Pt 12, 2597–2610. [Google Scholar] [CrossRef] [PubMed]
- Ikuta, K.; Ueda, S.; Kato, S.; Hirai, K. Most virus-specific polypeptides in cells productively infected with Marek’s disease virus or herpesvirus of turkeys possess cross-reactive determinants. J. Gen. Virol. 1983, 64 Pt 4, 961–965. [Google Scholar] [CrossRef]
- Tischer, B.K.; Schumacher, D.; Chabanne-Vautherot, D.; Zelnik, V.; Vautherot, J.F.; Osterrieder, N. High-level expression of Marek’s disease virus glycoprotein C is detrimental to virus growth in vitro. J. Virol. 2005, 79, 5889–5899. [Google Scholar] [CrossRef] [Green Version]
- Purchase, H.G.; Burmester, B.R.; Cunningham, C.H. Responses of cell cultures from various avian species to Marek’s disease virus and herpesvirus of turkeys. Am. J. Vet. Res. 1971, 32, 1811–1823. [Google Scholar]
- Mettenleiter, T.C. Glycoprotein gIII deletion mutants of pseudorabies virus are impaired in virus entry. Virology 1989, 171, 623–625. [Google Scholar] [CrossRef]
- Rue, C.A.; Ryan, P. Characterization of pseudorabies virus glycoprotein C attachment to heparan sulfate proteoglycans. J. Gen. Virol. 2002, 83 Pt 2, 301–309. [Google Scholar] [CrossRef]
- Osterrieder, N. Construction and characterization of an equine herpesvirus 1 glycoprotein C negative mutant. Virus Res. 1999, 59, 165–177. [Google Scholar] [CrossRef]
- Allen, G.P.; Coogle, L.D. Characterization of an equine herpesvirus type 1 gene encoding a glycoprotein (gp13) with homology to herpes simplex virus glycoprotein C. J. Virol. 1988, 62, 2850–2858. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, R.J.; Ponce de Leon, M.; Friedman, H.M.; Fries, L.F.; Frank, M.M.; Hastings, J.C.; Cohen, G.H. Complement component C3b binds directly to purified glycoprotein C of herpes simplex virus types 1 and 2. Microb. Pathog. 1987, 3, 423–435. [Google Scholar] [CrossRef]
- Friedman, H.M.; Cohen, G.H.; Eisenberg, R.J.; Seidel, C.A.; Cines, D.B. Glycoprotein C of herpes simplex virus 1 acts as a receptor for the C3b complement component on infected cells. Nature 1984, 309, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Rother, R.P.; Rollins, S.A.; Fodor, W.L.; Albrecht, J.C.; Setter, E.; Fleckenstein, B.; Squinto, S.P. Inhibition of complement-mediated cytolysis by the terminal complement inhibitor of herpesvirus saimiri. J. Virol. 1994, 68, 730–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fodor, W.L.; Rollins, S.A.; Bianco-Caron, S.; Rother, R.P.; Guilmette, E.R.; Burton, W.V.; Albrecht, J.C.; Fleckenstein, B.; Squinto, S.P. The complement control protein homolog of herpesvirus saimiri regulates serum complement by inhibiting C3 convertase activity. J. Virol. 1995, 69, 3889–3892. [Google Scholar] [CrossRef] [Green Version]
- Huemer, H.P.; Larcher, C.; van Drunen Littel-van den Hurk, S.; Babiuk, L.A. Species selective interaction of Alphaherpesvirinae with the “unspecific” immune system of the host. Arch. Virol. 1993, 130, 353–364. [Google Scholar] [CrossRef]
- Huemer, H.P.; Larcher, C.; Coe, N.E. Pseudorabies virus glycoprotein III derived from virions and infected cells binds to the third component of complement. Virus Res. 1992, 23, 271–280. [Google Scholar] [CrossRef]
- Ponnuraj, N.; Tien, Y.T.; Vega-Rodriguez, W.; Krieter, A.; Jarosinski, K.W. The Herpesviridae conserved multifunctional infected-cell protein 27 (ICP27) is important but not required for replication and oncogenicity of Marek’s disease alphaherpesvirus. J. Virol. 2019, 93, e01903-18. [Google Scholar] [CrossRef] [Green Version]
- Schat, K.A.; Sellers, H.S. Cell-culture methods. In A Laboratory Manual for the Identification and Characterization of Avian Pathogens, 5th ed.; Dufour-Zavala, L., Swayne, D.E., Glisson, J.R., Pearson, J.E., Reed, W.M., Jackwood, M.W., Woolcock, P.R., Eds.; American Association of Avian Pathologists: Jacksonville, FL, USA, 2008; pp. 195–203. [Google Scholar]
- Niikura, M.; Kim, T.; Silva, R.F.; Dodgson, J.; Cheng, H.H. Virulent Marek’s disease virus generated from infectious bacterial artificial chromosome clones with complete DNA sequence and the implication of viral genetic homogeneity in pathogenesis. J. Gen. Virol. 2011, 92 Pt 3, 598–607. [Google Scholar] [CrossRef]
- Baigent, S.J.; Petherbridge, L.J.; Smith, L.P.; Zhao, Y.; Chesters, P.M.; Nair, V.K. Herpesvirus of turkey reconstituted from bacterial artificial chromosome clones induces protection against Marek’s disease. J. Gen. Virol. 2006, 87 Pt 4, 769–776. [Google Scholar] [CrossRef]
- Mundt, A.; Mundt, E.; Hogan, R.J.; Garcia, M. Glycoprotein J of infectious laryngotracheitis virus is required for efficient egress of infectious virions from cells. J. Gen. Virol. 2011, 92 Pt 11, 2586–2589. [Google Scholar] [CrossRef] [PubMed]
- Jarosinski, K.W.; Osterrieder, N.; Nair, V.K.; Schat, K.A. Attenuation of Marek’s disease virus by deletion of open reading frame RLORF4 but not RLORF5a. J. Virol. 2005, 79, 11647–11659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramoff, M.D.; Magalhaes, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Jarosinski, K.W.; Yunis, R.; O’Connell, P.H.; Markowski-Grimsrud, C.J.; Schat, K.A. Influence of genetic resistance of the chicken and virulence of Marek’s disease virus (MDV) on nitric oxide responses after MDV infection. Avian Dis. 2002, 46, 636–649. [Google Scholar] [CrossRef]
- Rux, A.H.; Moore, W.T.; Lambris, J.D.; Abrams, W.R.; Peng, C.; Friedman, H.M.; Cohen, G.H.; Eisenberg, R.J. Disulfide bond structure determination and biochemical analysis of glycoprotein C from herpes simplex virus. J. Virol. 1996, 70, 5455–5465. [Google Scholar] [CrossRef] [Green Version]
- Pagni, M.; Ioannidis, V.; Cerutti, L.; Zahn-Zabal, M.; Jongeneel, C.V.; Hau, J.; Martin, O.; Kuznetsov, D.; Falquet, L. MyHits: Improvements to an interactive resource for analyzing protein sequences. Nucleic Acids Res. 2007, 35, W433–W437. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [Green Version]
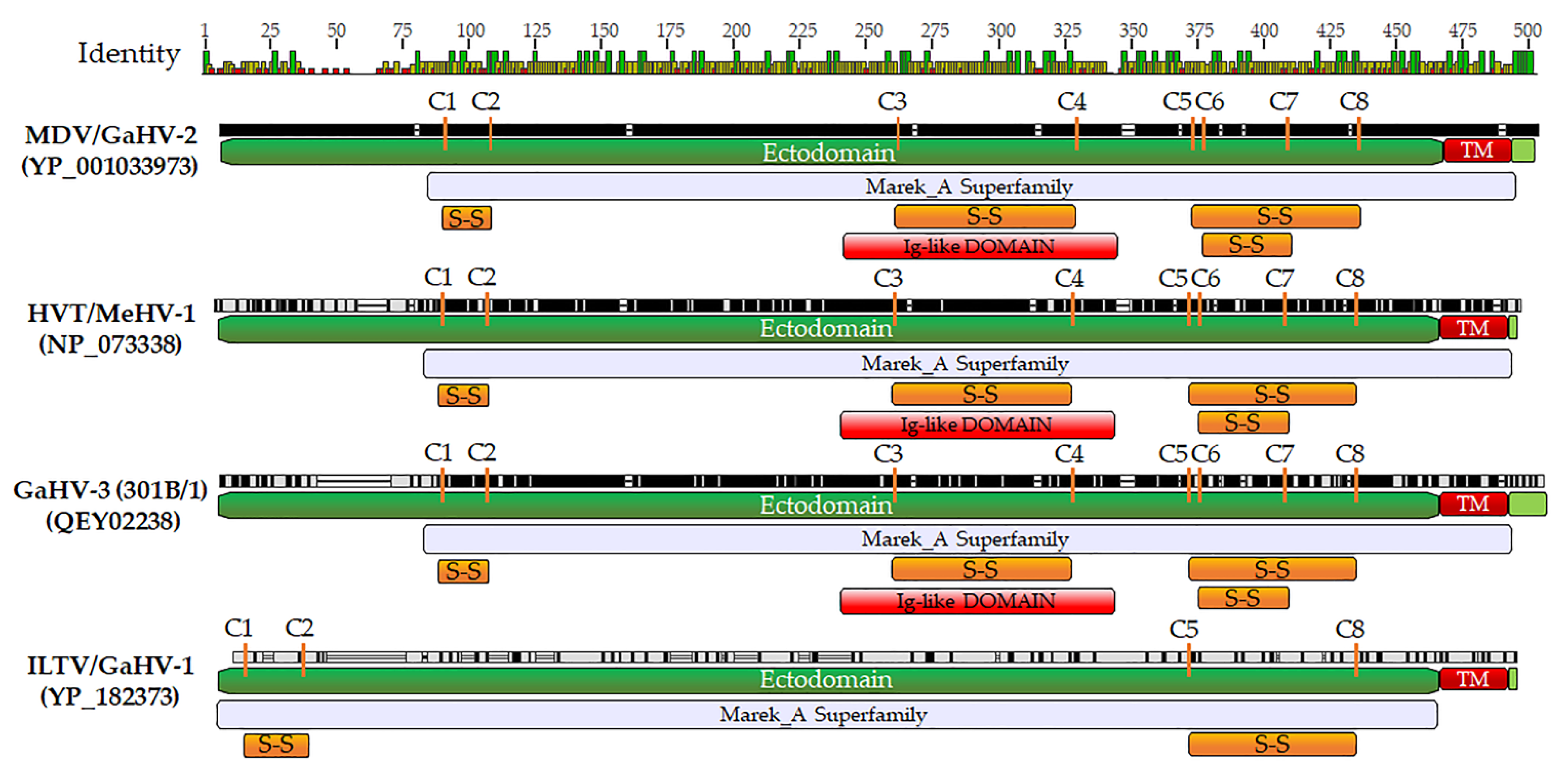

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct 1 | Direction 2 | Sequence (5′→3′) 3 |
|---|---|---|
| pcHVTgC | For | CGTAAGCTTTGTGTTTTATTGAGCGGTCG |
| Rev | CGTTCTAGATTTGGCCGCTGCGTGATACC | |
| pcHVTgC-in | For | CGACGGGATCCCCAGGGTTCTTTCTGGACTAGTCCTACACCCCGTGGAAATAGGGATAACAGGGTAATCGATTT |
| Rev | TTAACGGATCCGCCAGTGTTACAACCAATTAACC | |
| pcILTVgC-in | For | TCGCACTTAAGTGTTGAAGCGCTTGGCGCTTATCCTCCATCTGCTGCGCTGGGTATAGGGATAACAGGGTAATCGATTT |
| Rev | TTAACCTTAAGGCCAGTGTTACAACCAATTAACC | |
| pcHVTgC*-in | Vector For | TGAGTTTAAACCCGCTGATCGTTTAAACCCGCTGATCAGCCT |
| Vector Rev | TCGAAGGGCCCTCTAGACTCATTCCGCCCCGGTAGG | |
| Insert For | TTTACCTACCGGGGCGGAATGAGTCTAGAGGGCCCTTCGAACAAAA | |
| Insert Rev | GCTGATCAGCGGGTTTAAACGATCAGCGGGTTTAAACTCAATGGT |
| Modification 1 | Direction 2 | Sequence (5′→3′) 3 |
|---|---|---|
| MDV-HVTgC | For | ATACTAAACGATGGAGTTGTGTTTTATGAGCGTTGAAAACGATCCACTAGTAACGGCCGCCAG |
| Rev | TCACGTTTCTCCACTATTGCATTATTGTCTGACAAATAAAAGCTCTAGCATTTAGGTGACACT | |
| MDV-ILTVgC | For | ATACTAAACGATGGAGTTGTGTTTTATGAGCGTTGAAAACGATCCACTAGTAACGGCCGCCAG |
| Rev | TCACGTTTCTCCACTATTGCATTATTGTCTGACAAATAAAAGCTCTAGCATTTAGGTGACACT | |
| MDV-HVTgC* | For | ATACTAAACGATGGAGTTGTGTTTTATGAGCGTTGAAAACTGTGTTTTATTGAGCGGTCG |
| Rev | TCACGTTTCTCCACTATTGCATTATTGTCTGACAAATAAACTGATCAGCGGGTTTAAACG |
| Identities 1 | GaHV-1 (ILTV) | GaHV-2 (MDV) | GaHV-3 | MeHV-1 (HVT) |
|---|---|---|---|---|
| GaHV-1 (ILTV) | - | 15.2% | 18.3% | 16.4% |
| GaHV-2 (MDV) | 15.2% | - | 72.7% | 75.7% |
| GaHV-3 | 18.3% | 72.7% | - | 69.6% |
| MeHV-1 (HVT) | 16.4% | 75.7% | 69.6% | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega-Rodriguez, W.; Ponnuraj, N.; Garcia, M.; Jarosinski, K.W. The Requirement of Glycoprotein C for Interindividual Spread Is Functionally Conserved within the Alphaherpesvirus Genus (Mardivirus), but Not the Host (Gallid). Viruses 2021, 13, 1419. https://doi.org/10.3390/v13081419
Vega-Rodriguez W, Ponnuraj N, Garcia M, Jarosinski KW. The Requirement of Glycoprotein C for Interindividual Spread Is Functionally Conserved within the Alphaherpesvirus Genus (Mardivirus), but Not the Host (Gallid). Viruses. 2021; 13(8):1419. https://doi.org/10.3390/v13081419
Chicago/Turabian StyleVega-Rodriguez, Widaliz, Nagendraprabhu Ponnuraj, Maricarmen Garcia, and Keith W. Jarosinski. 2021. "The Requirement of Glycoprotein C for Interindividual Spread Is Functionally Conserved within the Alphaherpesvirus Genus (Mardivirus), but Not the Host (Gallid)" Viruses 13, no. 8: 1419. https://doi.org/10.3390/v13081419