
Challenges of Rabies Serology: Defining Context of Interpretation


{kind=link}
{kind=link}
Abstract
:1. Introduction
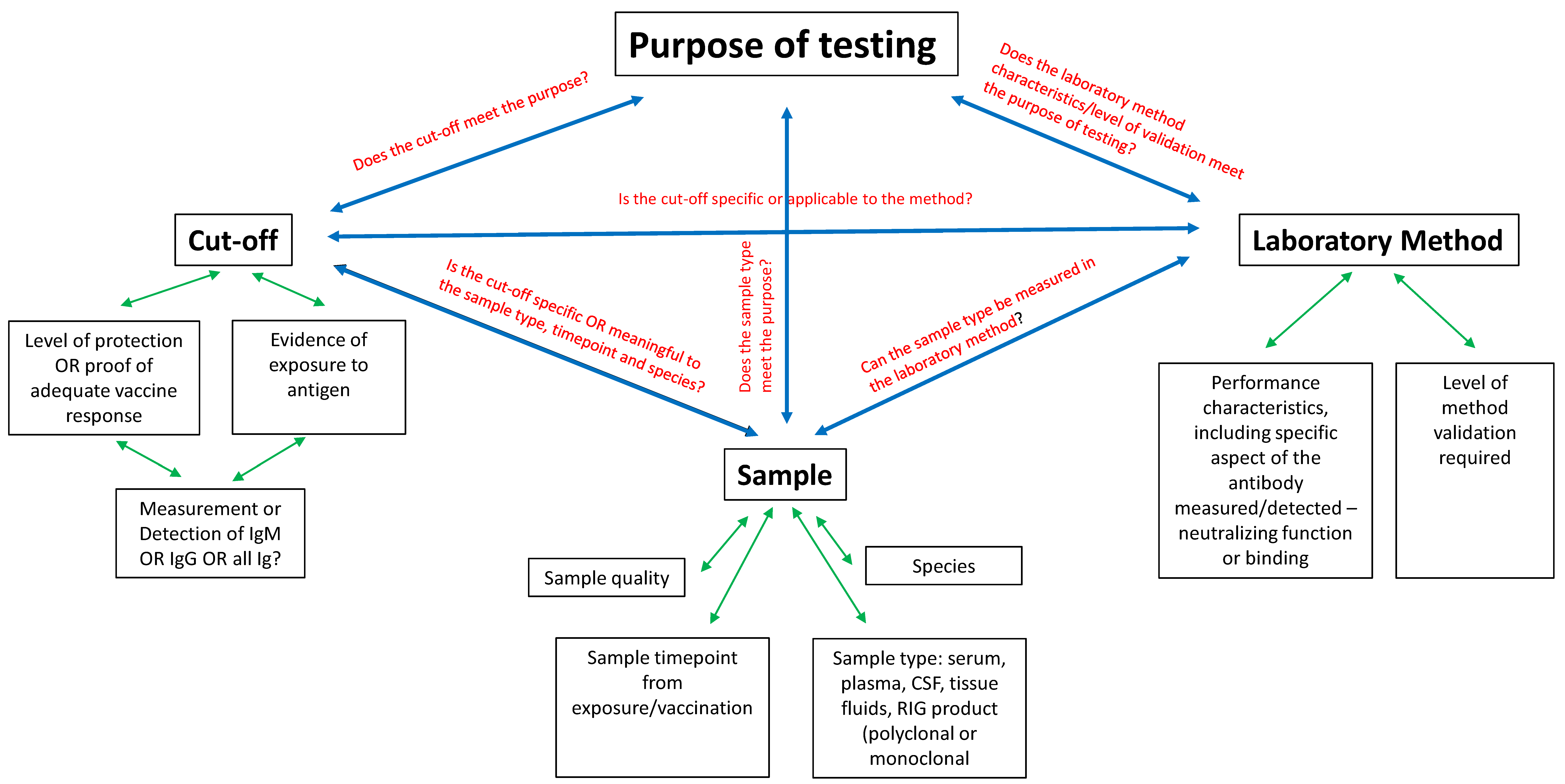
2. Defining the Purpose of Testing/Context of Interpretation
2.1. Cut-Off Level
2.2. Laboratory Method
2.3. Sample Considerations
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Finke, S.; Conzelmann, K.-K. Replication strategies of rabies virus. Virus Res. 2005, 111, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Dietzschold, B. Antibody-mediated clearance of viruses from the mammalian central nervous system. Trends Microbiol. 1993, 1, 63–66. [Google Scholar] [CrossRef]
- Dietzschold, B.; Gore, M.; Marchadier, D.; Niu, H.S.; Bunschoten, H.M.; Otvos, L., Jr.; Wunner, W.H.; Ertl, H.C.; Osterhaus, A.D.; Koprowski, H. Structural and immunological characterization of a linear virus-neutralizing epitope of the rabies virus glycoprotein and its possible use in a synthetic vaccine. J. Virol. 1990, 64, 3804–3809. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.C.; Morimoto, K.; Bette, M.; Weihe, E.; Koprowski, H.; Dietzschold, B. Collaboration of Antibody and Inflammation in Clearance of Rabies Virus from the Central Nervous System. J. Virol. 1998, 72, 3711–3719. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.C.; Phares, T.W.; Fabis, M.J.; Roy, A. The Production of Antibody by Invading B Cells Is Required for the Clearance of Rabies Virus from the Central Nervous System. PLoS Negl. Trop. Dis. 2009, 3, e535. [Google Scholar] [CrossRef]
- Hooper, D.C.; Roy, A.; Kean, R.B.; Phares, T.W.; Barkhouse, D.A. Therapeutic immune clearance of rabies virus from the CNS. Futur. Virol. 2011, 6, 387–397. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Expert Consultation on Rabies, Third Report; World Health Organization: Geneva, Switzerland, 2018; pp. 1–195. [Google Scholar]
- Rupprecht, C.E.; Briggs, D.; Brown, C.M.; Franka, R.; Katz, S.L.; Kerr, H.D.; Lett, S.M.; Levis, R.; Meltzer, M.I.; Schaffner, W.; et al. Use of a reduced (4-dose) vaccine schedule for postexposure prophylaxis to prevent human rabies: Recommendations of the advisory committee on immunization practices. MMWR Recomm. Rep. 2010, 59, 1–9. [Google Scholar]
- Tantawichien, T.; Rupprecht, C.E. Modern biologics for rabies prophylaxis and the elimination of human cases mediated by dogs. Expert Opin. Biol. Ther. 2020, 20, 1347–1359. [Google Scholar] [CrossRef]
- Potratz, M.; Zaeck, L.M.; Weigel, C.; Klein, A.; Freuling, C.M.; Müller, T.; Finke, S. Neuroglia infection by rabies virus after anterograde virus spread in peripheral neurons. Acta Neuropathol. Commun. 2020, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.C. Current and future approaches to the therapy of human rabies. Antivir. Res. 2013, 99, 61–67. [Google Scholar] [CrossRef]
- Webster, L.T. Rabies; The Macmillan Company: New York, NY, USA, 1942; p. 168. [Google Scholar]
- Steele, J. History of Rabies. In The Natural History of Rabies; Baer, G.M., Ed.; Academic Press: New York, NY, USA, 1975; Volume 1, pp. 1–28. [Google Scholar]
- Webster, L.T. Diagnostic and Immunological Tests of Rabies in Mice. Am. J. Public Health Nations Health 1936, 26, 1207–1210. [Google Scholar] [CrossRef]
- Smith, J.S.; Yager, P.A.; Baer, G.M. A rapid tissue culture test for determining rabies neutralizing antibody. Monogr. Ser. World Health Organ. 1973, 1973, 354–357. [Google Scholar]
- Bogel, K. Proposed International Reference Rabies Vaccine (HDC-Origin) and the Potency Tests Used to Test These Products. In Joint WHO/IABS Symposium on the Standardization of Rabies Vaccines for Human Use Produced in Tissue Culture [Rabies III]; S. Karger: Basel, Switzerland, 1978; pp. 267–271. [Google Scholar]
- Johnson, N.; Cunningham, A.F.; Fooks, A.R. The immune response to rabies virus infection and vaccination. Vaccine 2010, 28, 3896–3901. [Google Scholar] [CrossRef]
- Kostense, S.; Hendriks, J. Challenges of immunogenicity assays for vaccines. Bioanalysis 2012, 4, 397–406. [Google Scholar] [CrossRef]
- Bunn, T.O.; Ridpath, H.D. The relationship between rabies antibody titers in dogs and cats and protection from challenge. U. S. Department of Health, Education and Welfare. Public Health 1984, 11, 43–45. [Google Scholar]
- Aubert, M.F.A. Practical significance of rabies antibodies in cats and dogs. Rev. Sci. Tech. 1992, 11, 735–760. [Google Scholar] [CrossRef]
- Moore, S.M.; Gilbert, A.; Vos, A.; Freuling, C.M.; Ellis, C.; Kliemt, J.; Müller, T. Rabies Virus Antibodies from Oral Vaccination as a Correlate of Protection against Lethal Infection in Wildlife. Trop. Med. Infect. Dis. 2017, 2, 31. [Google Scholar] [CrossRef]
- Fooks, A. Rabies (Infection with RABV and Other Lyssaviruses). In Manual of Diagnotics Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health (OIE), Office International Des Epizooties: Paris, France, 2018; pp. 578–612. [Google Scholar]
- Kennedy, L.J.; Lunt, M.; Barnes, A.; McElhinney, L.; Fooks, A.R.; Baxter, D.N.; Ollier, W.E. Factors influencing the antibody response of dogs vaccinated against rabies. Vaccine 2007, 25, 8500–8507. [Google Scholar] [CrossRef]
- Wandeler, A.; (Canadian Food Inspection Agency, Ottawa, MB, Canada); Moore, S.M.; (Kansas State Univerisity Rabies Laboratory, Manhattan, KS, USA). WHO Level of Adequate Response to Rabies Vaccination (0.5 IU/mL). Personal communication, 2014. [Google Scholar]
- Smith, J.S.; (Centers for Disease Control and Prevention, Atlanta, GA, USA); Moore, S.M.; (Kansas State Univerisity Rabies Laboratory, Manhattan, KS, USA). ACIP Level of Adequate Response (Complete Neutralization at a 1:5 Serum Dilution) and Non Specific Inhibition. Personal communication, 2000. [Google Scholar]
- Sturgeon, C.M.; Viljoen, A. Analytical error and interference in immunoassay: Minimizing risk. Ann. Clin. Biochem. Int. J. Lab. Med. 2011, 48, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.M. Rabies Serology: Relationship between Assay Type, Interpretation, and Application of Results. In Diagnostic Medicine/Pathobiology/College of Veterinary Medicine; Kansas State University: Manhattan, KS, USA, 2015; p. 151. [Google Scholar]
- Briggs, D.J.; Dreesen, D.W.; Morgan, P.; Chin, J.E.; Seedle, C.D.; Cryz, L.; Glück, R.; Cryz, S.J. Safety and immunogenicity of Lyssavac Berna human diploid cell rabies vaccine in healthy adults. Vaccine 1996, 14, 1361–1365. [Google Scholar] [CrossRef]
- Sabchareon, A.; Chantavanich, P.; Pasuralertsakul, S.; Pojjaroen-Anant, C.; Prarinyanupharb, V.; Attanath, P.; Singhasivanon, V.; Buppodom, W.; Lang, J. Persistence of antibodies in children after intradermal or intramuscular administration of preexposure primary and booster immunizations with purified Vero cell rabies vaccine. Pediatr. Infect. Dis. J. 1998, 17, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Dreesen, D.W.; Fishbein, D.B.; Kemp, D.T.; Brown, J. Two-year comparative trial on the immunogenicity and adverse effects of purified chick embryo cell rabies vaccine for pre-exposure immunization. Vaccine 1989, 7, 397–400. [Google Scholar] [CrossRef]
- Gold, S.; Donnelly, C.A.; Nouvellet, P.; Woodroffe, R. Rabies virus-neutralising antibodies in healthy, unvaccinated individuals: What do they mean for rabies epidemiology? PLoS Negl. Trop. Dis. 2020, 14, e0007933. [Google Scholar] [CrossRef] [PubMed]
- Beran, J.; Honegr, K.; Banzhoff, A.; Malerczyk, C. Potency requirements of rabies vaccines administered intradermally using the Thai Red Cross regimen: Investigation of the immunogenicity of serially diluted purified chick embryo cell rabies vaccine. Vaccine 2005, 23, 3902–3907. [Google Scholar] [CrossRef]
- ICH. Guidelines for Validation of Analytical Procedures: Methodlolgy Q(2b), in Federal Register Vol 62, Number 96, 27464–27467; Food and Drug Administration, Health and Human Services: Rockville, MD, USA, 1997.
- Findlay, J.; Smith, W.; Lee, J.; Nordblom, G.; Das, I.; DeSilva, B.; Khan, M.; Bowsher, R. Validation of immunoassays for bioanalysis: A pharmaceutical industry perspective. J. Pharm. Biomed. Anal. 2000, 21, 1249–1273. [Google Scholar] [CrossRef]
- Moore, S.M.; Hanlon, C.A. Rabies-Specific Antibodies: Measuring Surrogates of Protection against a Fatal Disease. PLoS Negl. Trop. Dis. 2010, 4, e595. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.S.; Horton, D.; Müller, T.; Plowright, R.; Peel, A.; Bowen, R.; Wood, J.L.N.; Mills, J.; et al. Deciphering Serology to Understand the Ecology of Infectious Diseases in Wildlife. EcoHealth 2013, 10, 298–313. [Google Scholar] [CrossRef] [Green Version]
- Moore, S. Rabies prevention: The role of serology in parenteral vaccination of companion animals and livestock. Rev. Sci. Tech. 2018, 37, 461–472. [Google Scholar] [CrossRef]
- Moore, S.M.; Gordon, C.R. Measures of Rabies Immunity. In Rabies Scientific Basis of the Disease and Its Management; Jackson, A.C., Fooks, A.R., Eds.; Academic Press Elsevier: London, UK, 2020; pp. 445–480. [Google Scholar]
- Wasniewski, M.; Guiot, A.L.; Schereffer, J.L.; Tribout, L.; Mahar, K.; Cliquet, F. Evaluation of an ELISA to detect rabies antibodies in orally vaccinated foxes and raccoon dogs sampled in the field. J. Virol. Methods 2013, 187, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Feyssaguet, M.; Dacheux, L.; Audry, L.; Compoint, A.; Morize, J.; Blanchard, I.; Bourhy, H. Multicenter comparative study of a new ELISA, PLATELIA RABIES II, for the detection and titration of anti-rabies glycoprotein antibodies and comparison with the rapid fluorescent focus inhibition test (RFFIT) on human samples from vaccinated and non-vaccinated people. Vaccine 2007, 25, 2244–2251. [Google Scholar] [CrossRef]
- Dietzschold, B.; Tollis, M.; Lafon, M.; Wunner, W.H.; Koprowski, H. Mechanisms of rabies virus neutralization by glycoprotein-specific monoclonal antibodies. Virology 1987, 161, 29–36. [Google Scholar] [CrossRef]
- Irie, T.; Kawai, A. Further Studies on the Mechanism of Rabies Virus Neutralization by a Viral Glycoprotein-Specific Monoclonal Antibody, #1-46-12. Microbiol. Immunol. 2005, 49, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Lafon, M. Immunobiology of Lyssaviruses: The Basis for Immunoprotection. Curr. Top. Microbiol. Immunol. 1994, 187, 145–160. [Google Scholar] [CrossRef]
- Bioanalytical Method Validation Guidance for Industry 2018; U.S. Department of Health and Human Services, Food and Drug Administration: Rockville, MD, USA, 2018; p. 44.
- Moore, S.M.; Pralle, S.; Engelman, L.; Hartschuh, H.; Smith, M. Rabies vaccine response measurement is assay dependent. Biologicals 2016, 44, 481–486. [Google Scholar] [CrossRef]
- Moore, S.M.; Ricke, T.A.; Davis, R.D.; Briggs, D.J. The influence of homologous vs. heterologous challenge virus strains on the serological test results of rabies virus neutralizing assays. Biologicals 2005, 33, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, J.; Nie, J.; Li, J.; Cao, S.; Wang, L.; Yu, C.; Huang, W.; Li, Y.; Yu, Y.; et al. Antigenic variations of recent street rabies virus. Emerg. Microbes Infect. 2019, 8, 1584–1592. [Google Scholar] [CrossRef] [PubMed]
- Rathore, A.P.S.; St. John, A.L. Cross-Reactive Immunity Among Flaviviruses. Front. Immunol. 2020, 11, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.H.; McGowan, E.; Jadi, R.; Young, E.; Lopez, C.; Baric, R.S.; LaZear, H.M.; De Silva, A.M. Lack of Durable Cross-Neutralizing Antibodies Against Zika Virus from Dengue Virus Infection. Emerg. Infect. Dis. 2017, 23, 773–781. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Pan, H.; Chiu, C.Y.; Stremlau, M. Human Tibroviruses: Commensals or Lethal Pathogens? Viruses 2020, 12, 252. [Google Scholar] [CrossRef] [Green Version]
- Pearlman, E.S.; Ballas, S.K. False-positive human immunodeficiency virus screening test related to rabies vaccination. Arch. Pathol. Lab. Med. 1994, 118, 805–806. [Google Scholar]
- Rudd, R.J.; Appler, K.A.; Wong, S.J. Presence of Cross-Reactions with Other Viral Encephalitides in the Indirect Fluorescent-Antibody Test for Diagnosis of Rabies. J. Clin. Microbiol. 2013, 51, 4079–4082. [Google Scholar] [CrossRef] [Green Version]
- Zajac, M. Development and Evaluation of a Rabies Enzyme-Linked Immunosorbent Assay (ELISA) Targeting IgM and IgG in Human Sera. In Department of Diagnostic Medicine/Pathobiology; Kansas State University: Manhattan, KS, USA, 2019; p. 72. [Google Scholar]
- Ovsyannikova, I.G.; Schaid, D.J.; Larrabee, B.R.; Haralambieva, I.H.; Kennedy, R.B.; Poland, G.A. A large population-based association study between HLA and KIR genotypes and measles vaccine antibody responses. PLoS ONE 2017, 12, e0171261. [Google Scholar] [CrossRef]
- Schaid, D.J.; Haralambieva, I.H.; Larrabee, B.; Ovsyannikova, I.G.; Kennedy, R.B.; Poland, G.A. Heritability of vaccine-induced measles neutralizing antibody titers. Vaccine 2017, 35, 1390–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juanes-Velasco, P.; Landeira-Viñuela, A.; Acebes-Fernandez, V.; Hernández, Á.P.; Garcia-Vaquero, M.L.; Arias-Hidalgo, C.; Bareke, H.; Montalvillo, E.; Gongora, R.; Fuentes, M. Deciphering Human Leukocyte Antigen Susceptibility Maps From Immunopeptidomics Characterization in Oncology and Infections. Front. Cell. Infect. Microbiol. 2021, 11, 642583. [Google Scholar] [CrossRef] [PubMed]
- Kuwert, E.K.; Barsenbach, C.; Werner, J.; Marcus, I.; Scheiermann, N.; Grosse-Wilde, H.; Krumbacher, K. Early/High and Late/Low Responders among HDCS Vaccinees, in Cell Culture Rabies Vaccines and Their Protective Effect in Man; Wiktor, T.J., Koprowski, H., Eds.; International Green Cross: Geneva, Switzerland, 1981; pp. 160–168. [Google Scholar]
- CDC. Human Rabies Prevention—United States, 2008 Recommendatoins of the Advisory Committee On Immunization Practices, in MMWR; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2008; pp. 1–28.
- Strady, A.; Lang, J.; Lienard, M.; Blondeau, C.; Jaussaud, R.; Plotkin, S.A. Antibody persistence following preexposure regimens of cell-culture rabies vaccines: 10-year follow-up and proposal for a new booster policy. J. Infect. Dis. 1998, 177, 1290–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moore, S.M. Challenges of Rabies Serology: Defining Context of Interpretation. Viruses 2021, 13, 1516. https://doi.org/10.3390/v13081516
Moore SM. Challenges of Rabies Serology: Defining Context of Interpretation. Viruses. 2021; 13(8):1516. https://doi.org/10.3390/v13081516
Chicago/Turabian StyleMoore, Susan M. 2021. "Challenges of Rabies Serology: Defining Context of Interpretation" Viruses 13, no. 8: 1516. https://doi.org/10.3390/v13081516
APA StyleMoore, S. M. (2021). Challenges of Rabies Serology: Defining Context of Interpretation. Viruses, 13(8), 1516. https://doi.org/10.3390/v13081516