
A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus
2.3. Pathogenicity Testing
2.4. Sequencing
2.5. Collection of Sequences
2.6. Evolutionary and Phylogenetic Analyses
2.7. Chickens
2.8. Experimental Infection
2.9. Necropsy, Sample Collection and Histopathological Examination
3. Results
3.1. Pathogenicity Testing
3.2. Molecular Characterization and Classification
3.3. Pathogenicity of BD-P01 for Chickens
3.3.1. Clinical Signs and Mortality
3.3.2. Gross Lesions
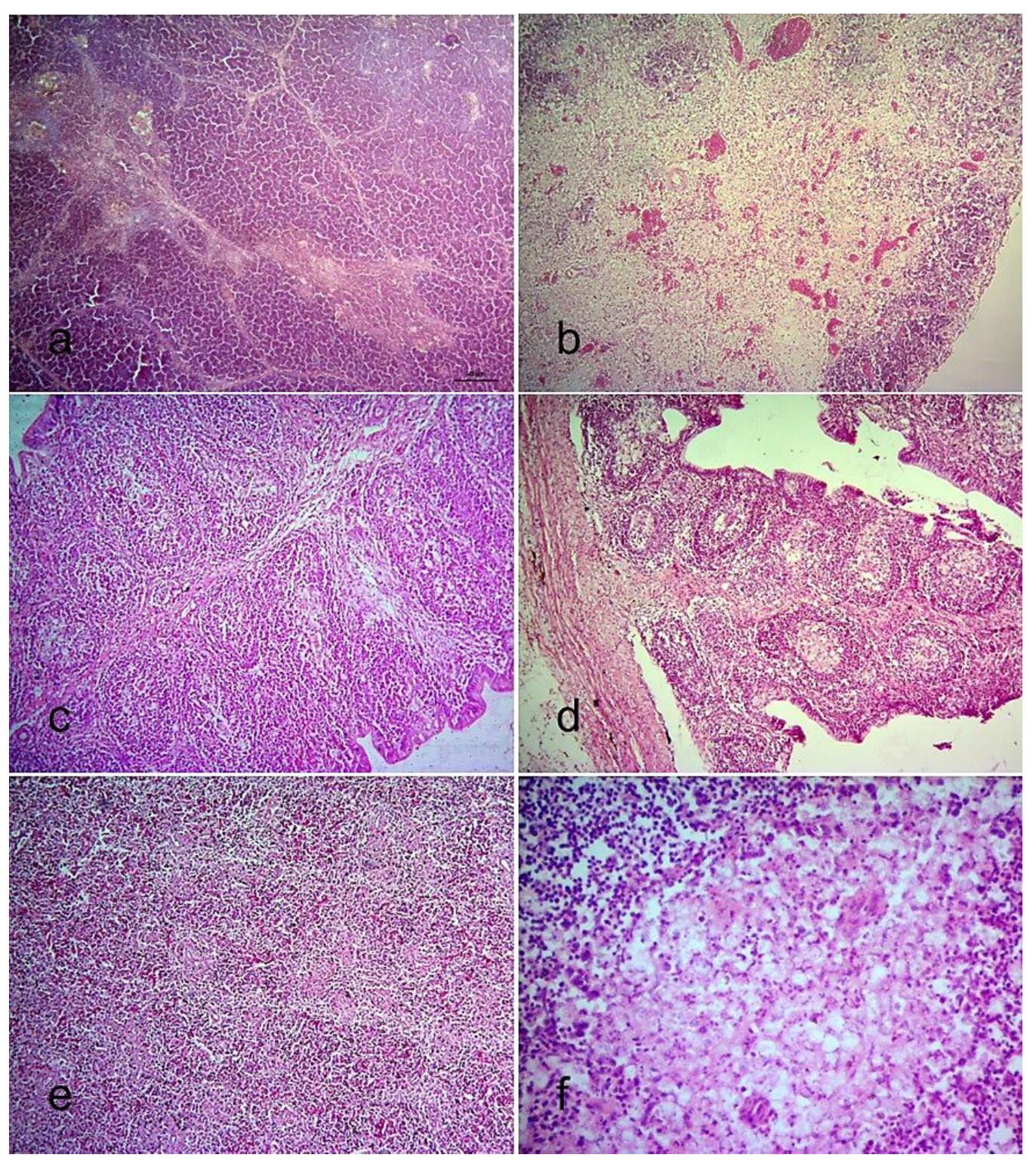
3.3.3. Histopathological Changes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amarasinghe, G.K.; Ayllón, M.A.; Bào, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo, M.A. A summary of taxonomic changes recently approved by ICTV. Arch. Virol. 2002, 147, 1655–1663. [Google Scholar] [CrossRef]
- Hanson, R.P.; Brandly, C.A. Identification of vaccine strains of Newcastle disease virus. Science 1955, 122, 156–157. [Google Scholar]
- Miller, P.J.; Koch, G. Newcastle disease. In Diseases of Poultry, 14th ed.; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 112–129. [Google Scholar]
- Alexander, D.J. Newcastle disease virus and other avian paramyxoviruses. In A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 4th ed.; Swayne, D.E., Glisson, J.R., Jackwood, M.W., Pearson, J.E., Reed, W.M., Eds.; The American Association of Avian Pathologists: Kennett Square, PA, USA, 1998; pp. 156–163. [Google Scholar]
- Panda, A.; Huang, Z.; Elankumaran, S.; Rockemann, D.D.; Samal, S.K. Role of fusion protein cleavage site in the virulence of Newcastle disease virus. Microb. Pathog. 2004, 36, 1–10. [Google Scholar] [CrossRef]
- Huang, Z.; Panda, A.; Elankumaran, S.; Govindarajan, D.; Rockemann, D.D.; Samal, S.K. The hemagglutinin-neuraminidase protein of Newcastle disease virus determines tropism and virulence. J. Virol. 2004, 78, 4176–4184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, P.; Millar, N.S.; Emmerson, P.T. Nucleotide sequence of the gene encoding the fusion glycoprotein of Newcastle disease virus. J. Gen. Virol. 1986, 67, 2685–2694. [Google Scholar] [CrossRef]
- Glickman, R.L.; Syddall, R.J.; Iorio, R.M.; Sheehan, J.P.; Bratt, M.A. Quantitative basic residue requirements in the cleavage-activation site of the fusion glycoprotein as a determinant of virulence for Newcastle disease virus. J. Virol. 1988, 62, 354–356. [Google Scholar] [CrossRef] [Green Version]
- OIE. Chapter 3.3.14. Newcastle Disease (Infection with Newcastle Disease Virus). In OIE Terrestrial Mannual 2018; World Organization for Animal Health: Paris, France, 2018; pp. 964–983. [Google Scholar]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.X.; Brown, I.H.; Choi, K.S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Diel, D.G.; da Silva, L.H.; Liu, H.; Wang, Z.; Miller, P.J.; Afonso, C.L. Genetic diversity of avian paramyxovirus type 1: Proposal for a unified nomenclature and classification system of Newcastle disease virus genotypes. Infect. Genet. Evol. 2012, 12, 1770–1779. [Google Scholar] [CrossRef]
- Kim, L.M.; King, D.J.; Curry, P.E.; Suarez, D.L.; Swayne, D.E.; Stallknecht, D.E.; Slemons, R.D.; Pedersen, J.C.; Senne, D.A.; Winker, K.; et al. Phylogenetic diversity among low-virulence Newcastle disease viruses from waterfowl and shorebirds and comparison of genotype distributions to those of poultry-origin isolates. J. Virol. 2007, 81, 12641–12653. [Google Scholar] [CrossRef] [Green Version]
- Kim, L.M.; King, D.J.; Suarez, D.L.; Wong, C.W.; Afonso, C.L. Characterization of class I Newcastle disease virus isolates from Hong Kong live bird markets and detection using real-time reverse transcription-PCR. J. Clin. Microbiol. 2007, 45, 1310–1314. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.J.; Haddas, R.; Simanov, L.; Lublin, A.; Rehmani, S.F.; Wajid, A.; Bibi, T.; Khan, T.A.; Yaqub, T.; Setiyaningsih, S.; et al. Identification of new sub-genotypes of virulent Newcastle disease virus with potential panzootic features. Infect. Genet. Evol. 2015, 29, 216–229. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lu, B.; Dimitrov, K.M.; Liang, J.; Chen, Z.; Zhao, W.; Qin, Y.; Duan, Q.; Zhou, Y.; Liu, L.; et al. Complete genome sequencing, molecular epidemiological, and pathogenicity analysis of pigeon paramyxoviruses type 1 isolated in Guangxi, China during 2012–2018. Viruses 2020, 12, 366. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.J.; Decanini, E.L.; Afonso, C.L. Newcastle disease: Evolution of genotypes and the related diagnostic challenges. Infect. Genet. Evol. 2010, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.S.; Alexander, D.J.; Brockman, S.; Kemp, P.A.; Manvell, R.J. Evaluation of mouse monoclonal antibodies raised against an isolate of the variant avian paramyxovirus type 1 responsible for the current panzootic in pigeons. Arch. Virol. 1989, 104, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Kim, L.M.; Afonso, C.L.; Suarez, D.L. Effect of probe-site mismatches on detection of virulent Newcastle disease viruses using a fusion-gene real-time reverse transcription polymerase chain reaction test. J. Vet. Diagn. Investig. 2006, 18, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ren, S.; Wang, X.; Wang, C.Y.; Fan, M.; Jia, Y.; Gao, X.; Liu, H.; Xiao, S.; Yang, Z. Genomic characterization of a wild-bird-origin pigeon paramyxovirus type 1 (PPMV-1) first isolated in the northwest region of China. Arch. Virol. 2017, 162, 749–761. [Google Scholar] [CrossRef]
- Ganar, K.; Das, M.; Raut, A.A.; Mishra, A.; Kumar, S. Emergence of a deviating genotype VI pigeon paramyxovirus type-1 isolated from India. Arch. Virol. 2017, 162, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Hu, S.; Chen, Y.; Lu, X.; Liao, T.; Guo, L.; Gao, X.; Xu, X.; Hu, Z.; Liu, X.; et al. Comparative pathogenicity of two closely related Newcastle disease virus isolates from chicken and pigeon respectively. Virus Res. 2020, 286, 198091. [Google Scholar] [CrossRef]
- Ren, S.; Wang, C.; Zhang, X.; Zhao, L.; Wang, X.; Yao, W.; Han, Q.; Wang, Y.; Fan, M.; Gao, X.; et al. Phylogenetic and pathogenic characterization of a pigeon paramyxovirus type 1 isolate reveals cross-species transmission and potential outbreak risks in the northwest region of China. Arch. Virol. 2017, 162, 2755–2767. [Google Scholar] [CrossRef]
- Mayahi, M.; Seyfi Abad Shapouri, M.R.; Jafari, R.A.; Khosravi Farsani, M. Characterization of isolated pigeon paramyxovirus-1 (PMV-1) and its pathogenicity in broiler chickens. Vet. Res. Forum 2017, 8, 15–21. [Google Scholar]
- Guo, H.; Liu, X.; Xu, Y.; Han, Z.; Shao, Y.; Kong, X.; Liu, S. A comparative study of pigeons and chickens experimentally infected with PPMV-1 to determine antigenic relationships between PPMV-1 and NDV strains. Vet. Microbiol. 2014, 168, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Hüppi, L.; Ruggli, N.; Python, S.; Hoop, R.; Albini, S.; Grund, C.; Vögtlin, A. Experimental pigeon paramyxovirus-1 infection in chicken: Evaluation of infectivity, clinical and pathological manifestations and diagnostic methods. J. Gen. Virol. 2020, 101, 156–167. [Google Scholar] [CrossRef]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Carmichael, K.P.; Brown, C.C. Pathogenesis of six pigeon-origin isolates of Newcastle disease virus for domestic chickens. Vet. Pathol. 2002, 39, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, H.L.; Taylor, T.L.; Dimitrov, K.M.; Sabra, M.; Afonso, C.L.; Suarez, D.L. Virulent Newcastle disease viruses from chicken origin are more pathogenic and transmissible to chickens than viruses normally maintained in wild birds. Vet. Microbiol. 2019, 235, 25–34. [Google Scholar] [CrossRef]
- Dortmans, J.C.; Fuller, C.M.; Aldous, E.W.; Rottier, P.J.; Peeters, B.P. Two genetically closely related pigeon paramyxovirus type 1 (PPMV-1) variants with identical velogenic fusion protein cleavage sites but with strongly contrasting virulence. Vet. Microbiol. 2010, 143, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Meulemans, G.; van den Berg, T.P.; Decaesstecker, M.; Boschmans, M. Evolution of pigeon Newcastle disease virus strains. Avian Pathol. 2002, 31, 515–519. [Google Scholar] [CrossRef] [PubMed]
- Dortmans, J.C.F.M.; Rottier, P.J.M.; Koch, G.; Peeters, B.P.H. Passaging of a Newcastle disease virus pigeon variant in chickens results in selection of viruses with mutations in the polymerase complex enhancing virus replication and virulence. J. Gen. Virol. 2011, 92, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Kommers, G.D.; King, D.J.; Seal, B.S.; Brown, C.C. Virulence of pigeon-origin Newcastle disease virus isolates for domestic chickens. Avian Dis. 2001, 45, 906–921. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.S.; Strong, I.; Alexander, D.J. Evaluation of the molecular basis of pathogenicity of the variant Newcastle disease viruses termed “pigeon PMV-1 viruses”. Arch. Virol. 1994, 134, 403–411. [Google Scholar] [CrossRef]
- Fuller, C.M.; Collins, M.S.; Easton, A.J.; Alexander, D.J. Partial characterisation of five cloned viruses differing in pathogenicity, obtained from a single isolate of pigeon paramyxovirus type 1 (PPMV-1) following passage in fowls’ eggs. Arch. Virol. 2007, 152, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Alexander, D.J.; Parsons, G.; Marshall, R. Infection of fowls with Newcastle disease virus by food contaminated with pigeon faeces. Vet. Rec. 1984, 115, 601–602. [Google Scholar] [CrossRef]
- Abolnik, C.; Gerdes, G.H.; Kitching, J.; Swanepoel, S.; Romito, M.; Bisschop, S.P. Characterization of pigeon paramyxoviruses (Newcastle disease virus) isolated in South Africa from 2001 to 2006. Onderstepoort J. Vet. Res. 2008, 75, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Biswas, P.K.; Barua, H.; Uddin, G.M.; Biswas, D.; Ahad, A.; Debnath, N.C. Serosurvey of five viruses in chickens on smallholdings in Bangladesh. Prev. Vet. Med. 2009, 88, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Xue, C.; Islam, M.R.; Islam, M.T.; Cao, Y. A longitudinal study on the incidence of mortality of infectious diseases of commercial layer birds in Bangladesh. Prev. Vet. Med. 2013, 109, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Nooruzzaman, M.; Mazumder, A.C.; Khatun, S.; Chowdhury, E.H.; Das, P.M.; Islam, M.R. Pathotypic and genotypic characterization of two Bangladeshi isolates of Newcastle disease virus of chicken and pigeon origin. Transbound. Emerg. Dis. 2015, 62, 102–107. [Google Scholar] [CrossRef]
- Barman, L.R.; Nooruzzaman, M.; Sarker, R.D.; Rahman, M.T.; Saife, M.R.B.; Giasuddin, M.; Das, B.C.; Das, P.M.; Chowdhury, E.H.; Islam, M.R. Phylogenetic analysis of Newcastle disease viruses from Bangladesh suggests continuing evolution of genotype XIII. Arch. Virol. 2017, 162, 3177–3182. [Google Scholar] [CrossRef]
- Kabiraj, C.K.; Mumu, T.T.; Chowdhury, E.H. Sequential pathology of a genotype XIII Newcastle disease virus from Bangladesh in chickens on experimental infection. Pathogens 2020, 9, 539. [Google Scholar] [CrossRef]
- Mazumder, A.C.; Khatun, S.; Nooruzzaman, M.; Chowdhury, E.H.; Das, B.C.; Islam, M.R. Isolation and identification of Newcastle disease viruses from field outbreaks in chickens and pigeons. Bangladesh Vet. 2012, 29, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.R.; Zierenberg, K.; Eterradossi, N.; Toquin, D.; Rivallan, G.; Müller, H. Molecular and antigenic characterization of Bangladeshi isolates of infectious bursal disease virus demonstrate their similarities with recent European, Asian and African very virulent strains. J. Vet. Med. B Infect. Dis. Vet. Public Health 2001, 48, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Fouchier, R.A.; Bestebroer, T.M.; Herfst, S.; Van Der Kemp, L.; Rimmelzwaan, G.F.; Osterhaus, A.D. Detection of influenza A viruses from different species by PCR amplification of conserved sequences in the matrix gene. J. Clin. Microbiol. 2000, 38, 4096–4101. [Google Scholar] [CrossRef] [Green Version]
- Gelb, J., Jr.; Weisman, Y.; Ladman, B.S.; Meir, R. S1 gene characteristics and efficacy of vaccination against infectious bronchitis virus field isolates from the United States and Israel (1996 to 2000). Avian Pathol. 2005, 34, 194–203. [Google Scholar] [CrossRef]
- Nooruzzaman, M.; Mumu, T.T.; Kabiraj, C.K.; Hasnat, A.; Rahman, M.M.; Chowdhury, E.H.; Dimitrov, K.M.; Islam, M.R. Genetic and biological characterization of Newcastle disease viruses circulating in Bangladesh during 2010-2017: Further genetic diversification of class II genotype XIII in Southcentral Asia. J. Gen. Virol. 2021, 102, 001554. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Tavaré, S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Stamatakis, A.; Aberer, A.J.; Goll, C.; Smith, S.A.; Berger, S.A.; Izquierdo-Carrasco, F. RAxML-Light: A tool for computing terabyte phylogenies. Bioinformatics 2012, 28, 2064–2066. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Suchard, M.A.; Lemey, P. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ho, S.Y.; Phillips, M.J.; Rambaut, A. Relaxed phylogenetics and dating with confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Ayres, D.L.; Darling, A.; Zwickl, D.J.; Beerli, P.; Holder, M.T.; Lewis, P.O.; Huelsenbeck, J.P.; Ronquist, F.; Swofford, D.L.; Cummings, M.P.; et al. BEAGLE: An application programming interface and high-performance computing library for statistical phylogenetics. Syst. Biol. 2011, 61, 170–173. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Luna, L.G. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology, 3rd ed.; McGrow Hill Book Co.: New York, NY, USA, 1988. [Google Scholar]
- Chong, Y.L.; Lam, T.T.-Y.; Kim, O.; Lu, H.; Dunn, P.; Poss, M. Successful establishment and global dispersal of genotype VI avian paramyxovirus serotype 1 after cross species transmission. Infect. Genet. Evol. 2013, 17, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.T.; Dimitrov, K.M.; Afonso, C.L.; Ramey, A.M.; Bahl, J. Global phylodynamic analysis of avian paramyxovirus-1 provides evidence of inter-host transmission and intercontinental spatial diffusion. BMC Evol. Biol. 2019, 19, 108. [Google Scholar] [CrossRef]
- Diel, D.G.; Susta, L.; Cardenas Garcia, S.; Killian, M.L.; Brown, C.C.; Miller, P.J.; Afonso, C.L. Complete genome and clinicopathological characterization of a virulent Newcastle disease virus isolate from South America. J. Clin. Microbiol. 2012, 50, 378–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vindevogel, H.; Duchatel, J.P. Panzootic Newcastle disease virus in pigeons. In Newcastle Disease, Development of Veterinary Virology; Alexander, D.J., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 184–196. [Google Scholar]
- Alexander, D.J. Newcastle disease in the European Union 2000 to 2009. Avian Pathol. 2011, 40, 547–558. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.J.; Parsons, G. Pathogenicity for chickens of avian paramyxovirus type 1 isolates obtained from pigeons in Great Britain during 1983–85. Avian Pathol. 1986, 15, 487–493. [Google Scholar] [CrossRef]
- Naguib, M.M.; Höper, D.; Elkady, M.F.; Afifi, M.A.; Erfan, A.; Abozeid, H.H.; Hasan, W.M.; Arafa, A.-S.; Shahein, M.; Beer, M.; et al. Comparison of genomic and antigenic properties of Newcastle disease virus genotypes II, XXI and VII from Egypt do not point to antigenic drift as selection marker. Transbound. Emerg. Dis. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Afonso, C.L. Virulence during Newcastle disease viruses cross species adaptation. Viruses 2021, 13, 110. [Google Scholar] [CrossRef]
- Soñora, M.; Moreno, P.; Echeverría, N.; Fischer, S.; Comas, V.; Fajardo, A.; Cristina, J. An evolutionary insight into Newcastle disease viruses isolated in Antarctica. Arch. Virol. 2015, 160, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Terregino, C.; Dalla Pozza, M.; Marangon, S.; Mutinelli, F. Newcastle disease outbreaks in Italy during 2000. Vet. Rec. 2002, 150, 565–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehmani, S.F.; Wajid, A.; Bibi, T.; Nazir, B.; Mukhtar, N.; Hussain, A.; Lone, N.A.; Yaqub, T.; Afonso, C.L. Presence of virulent Newcastle disease virus in vaccinated chickens in farms in Pakistan. J. Clin. Microbiol. 2015, 53, 1715–1718. [Google Scholar] [CrossRef] [Green Version]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]
- Parede, L.; Young, P.L. The pathogenesis of velogenic Newcastle disease virus infection of chickens of different ages and different levels of immunity. Avian Dis. 1990, 34, 803–808. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub/Genotype | No. of Base Substitutions per Site a | |||||
|---|---|---|---|---|---|---|
| BD-P01 | XXI.1.1 | XXI.1.2 | XXI.2 | XXI | XX | |
| BD-P01 | ||||||
| XXI.1.1 | 0.092 | |||||
| XXI.1.2 | 0.076 | 0.093 | ||||
| XXI.2 | 0.120 | 0.105 | 0.124 | |||
| XXI | 0.147 | 0.125 | 0.139 | 0.148 | ||
| XX | 0.110 | 0.096 | 0.111 | 0.118 | 0.127 | |
| VI | 0.124 | 0.108 | 0.125 | 0.131 | 0.148 | 0.102 |
| Days Post Infection | No. of Birds under Observation | No. of Birds Sick | No. of Birds Died | No. of Birds Sacrificed (Sick/Normal) |
|---|---|---|---|---|
| 1 | 31 | 0 | 0 | 3 |
| 2 | 28 | 0 | 0 | 3 |
| 3 | 25 | 0 | 0 | 3 |
| 5 | 22 | 0 | 0 | 3 |
| 7 | 19 | 4 | 3 | - |
| 8 | 16 | 5 | 3 | - |
| 9 | 13 | 3 | 10 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nooruzzaman, M.; Barman, L.R.; Mumu, T.T.; Chowdhury, E.H.; Dimitrov, K.M.; Islam, M.R. A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens. Viruses 2021, 13, 1520. https://doi.org/10.3390/v13081520
Nooruzzaman M, Barman LR, Mumu TT, Chowdhury EH, Dimitrov KM, Islam MR. A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens. Viruses. 2021; 13(8):1520. https://doi.org/10.3390/v13081520
Chicago/Turabian StyleNooruzzaman, Mohammed, Lalita Rani Barman, Tanjin Tamanna Mumu, Emdadul Haque Chowdhury, Kiril M. Dimitrov, and Mohammad Rafiqul Islam. 2021. "A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens" Viruses 13, no. 8: 1520. https://doi.org/10.3390/v13081520
APA StyleNooruzzaman, M., Barman, L. R., Mumu, T. T., Chowdhury, E. H., Dimitrov, K. M., & Islam, M. R. (2021). A Pigeon-Derived Sub-Genotype XXI.1.2 Newcastle Disease Virus from Bangladesh Induces High Mortality in Chickens. Viruses, 13(8), 1520. https://doi.org/10.3390/v13081520