
Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Setup
2.2. Blood Collection, Processing, and Analyses
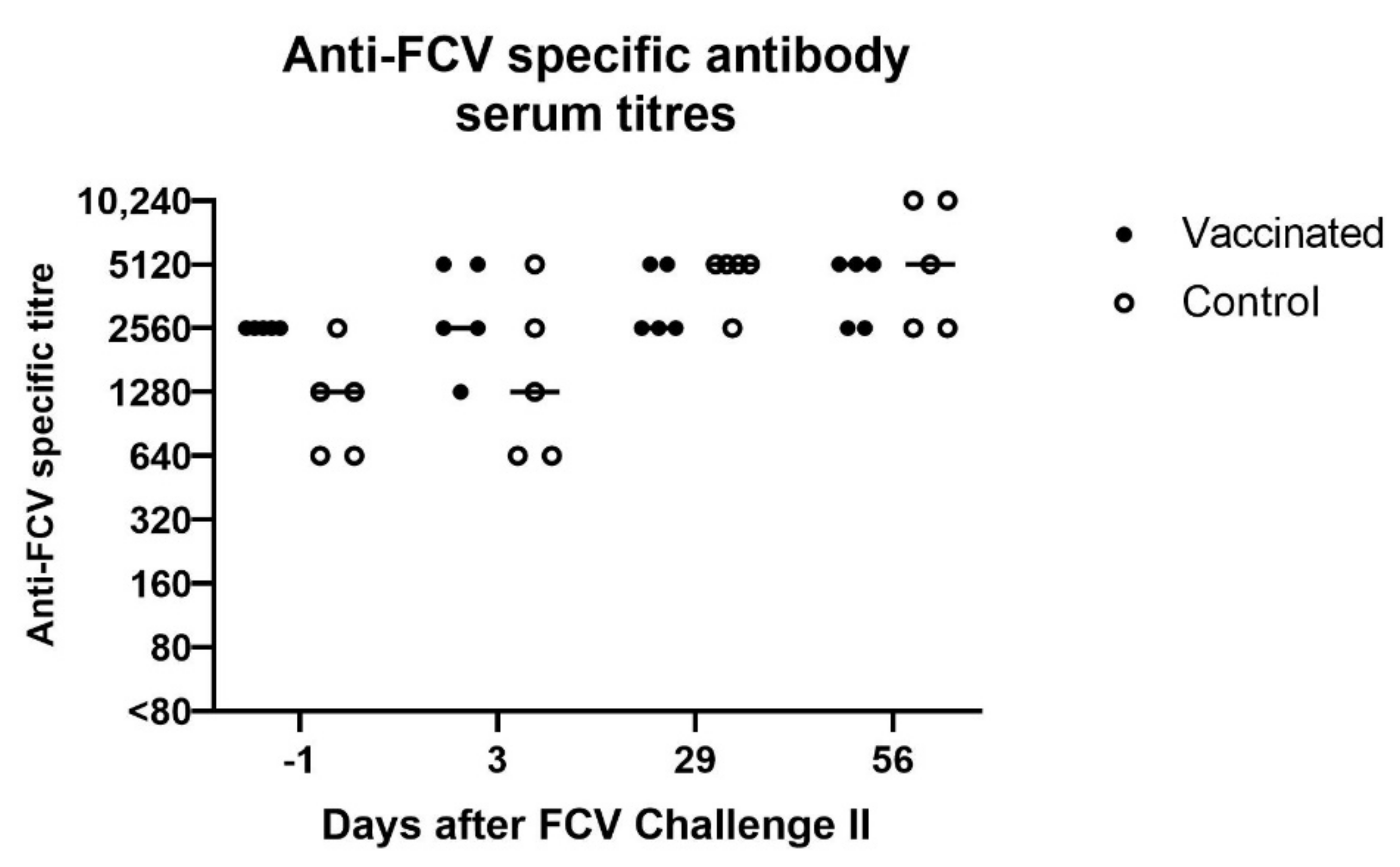
2.3. FCV Seroreactive Antibodies
2.4. FCV-Neutralising Antibodies
2.5. Cytokines
2.6. Lymphocyte Subsets
2.7. Feline IFN-γ ELISpot
2.8. Statistics
3. Results
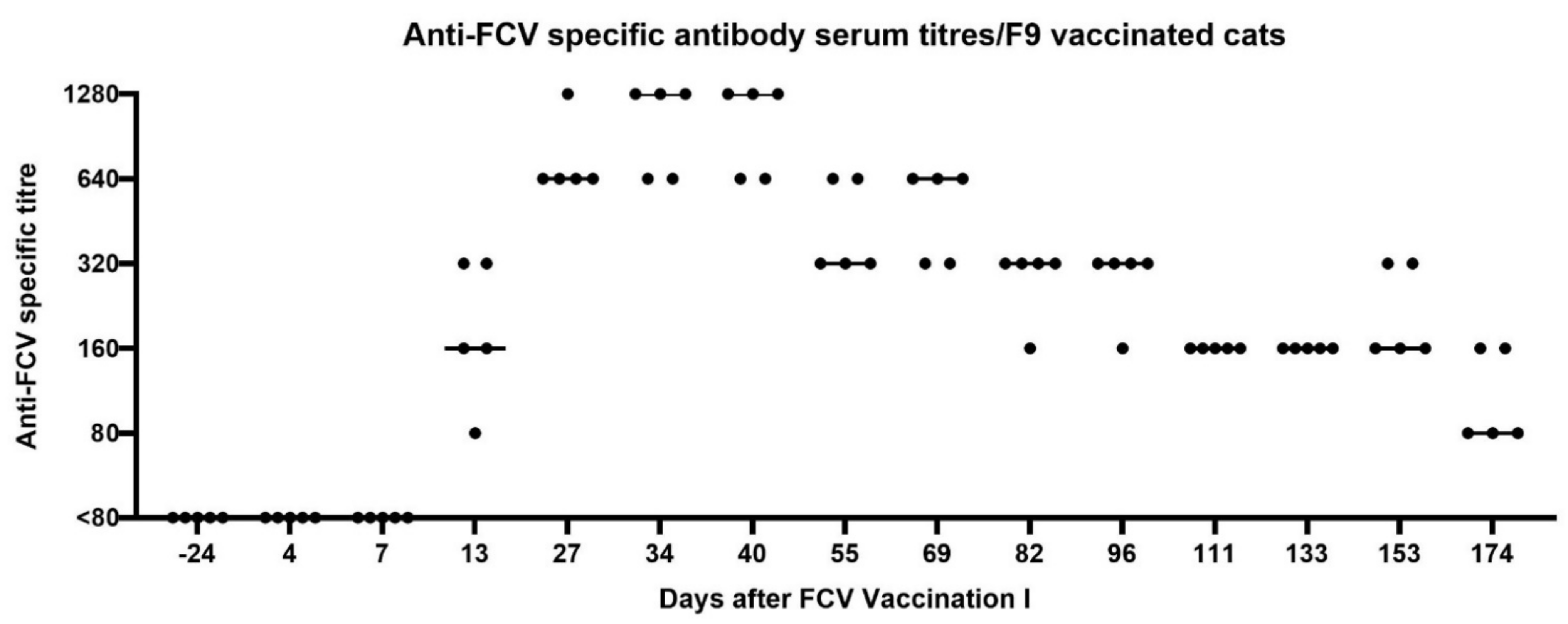
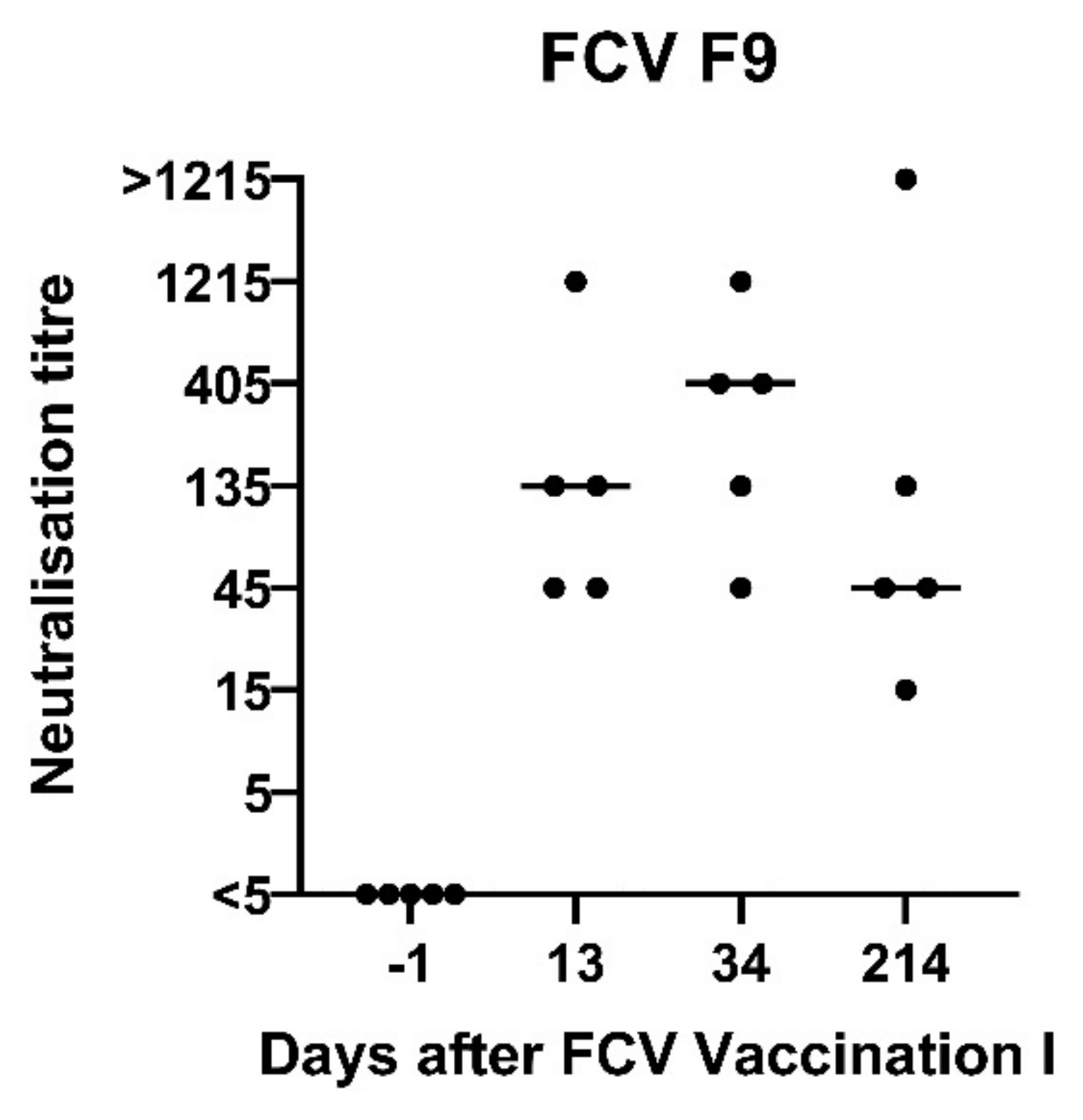
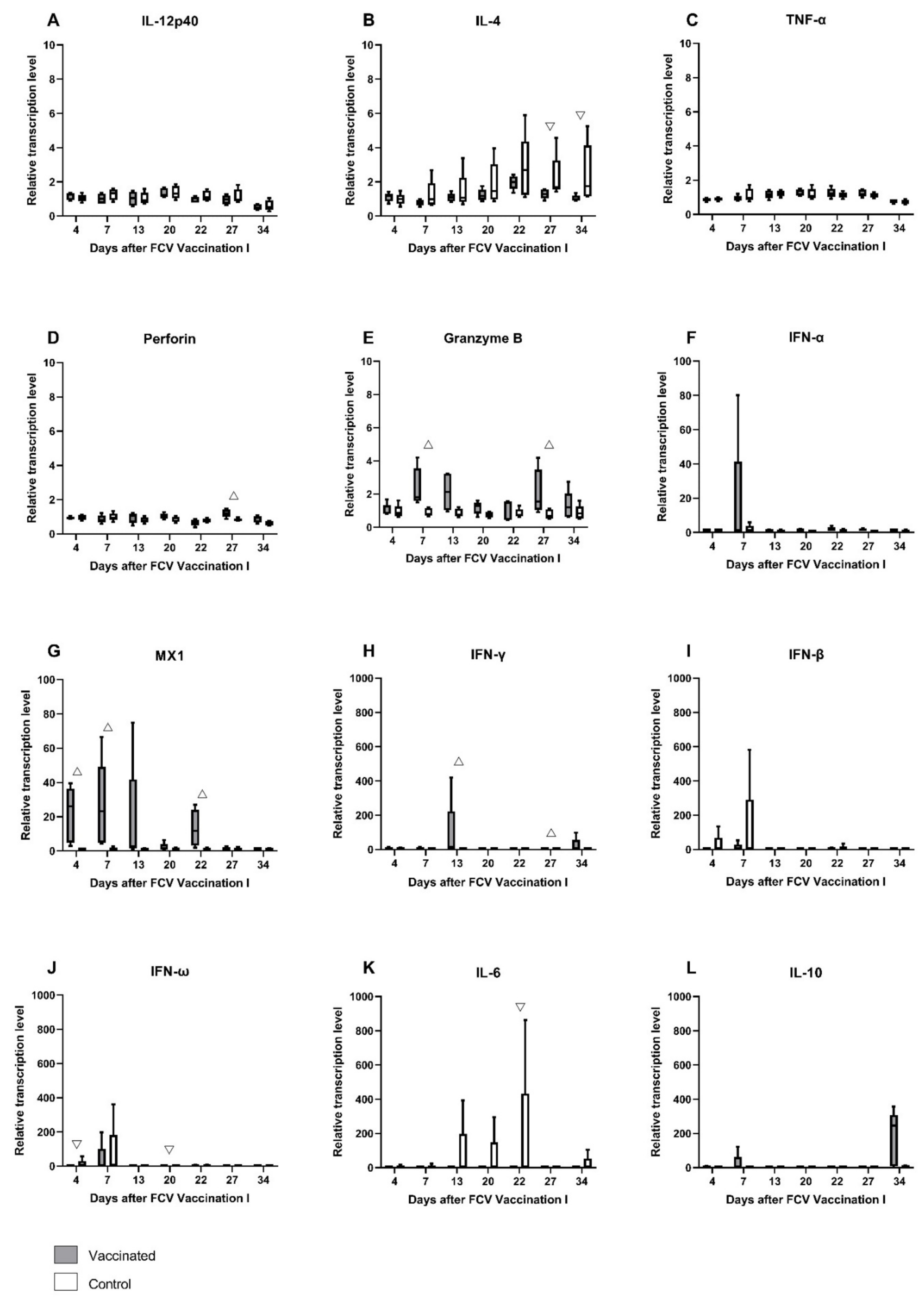
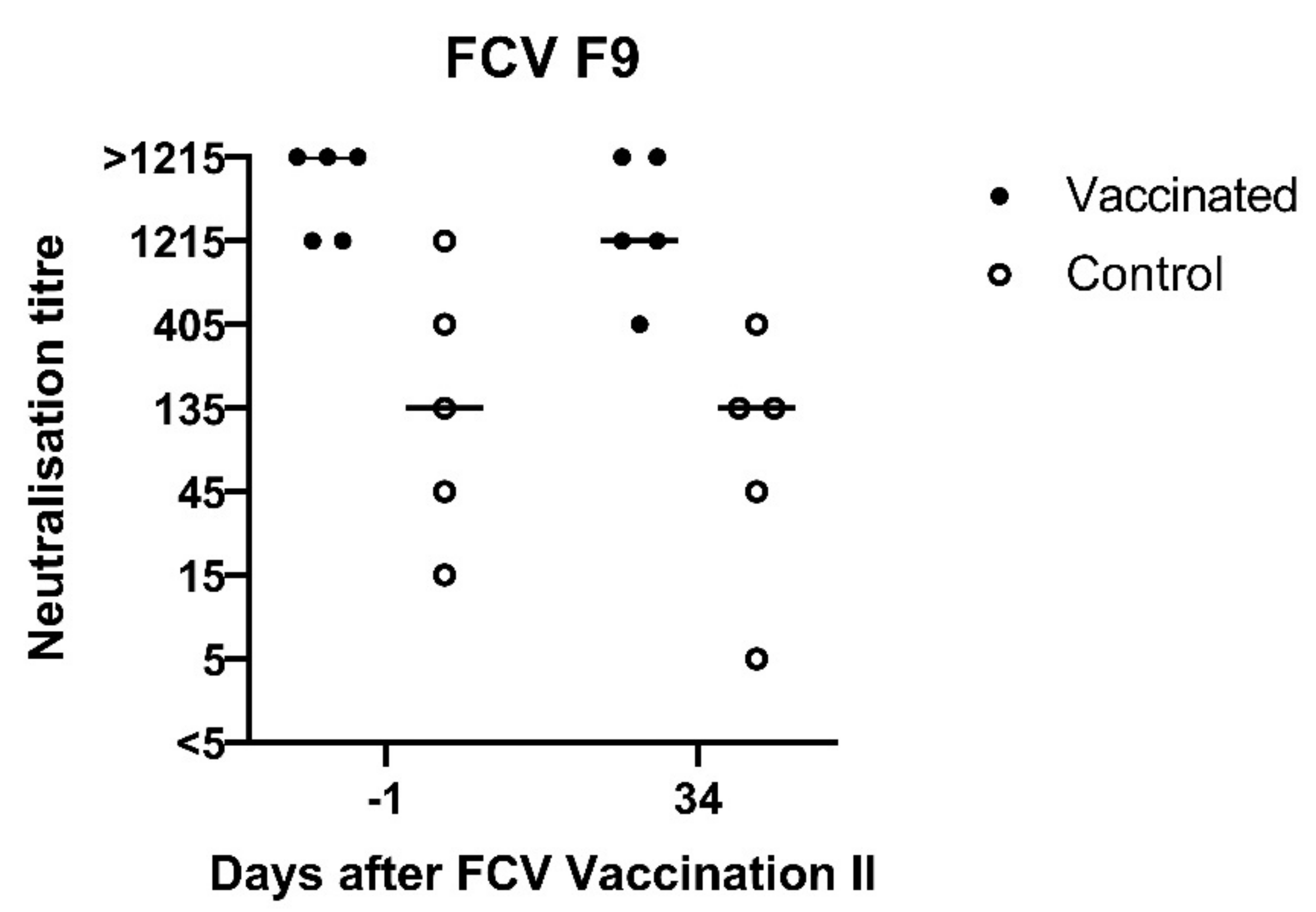
3.1. FCV Vaccination I
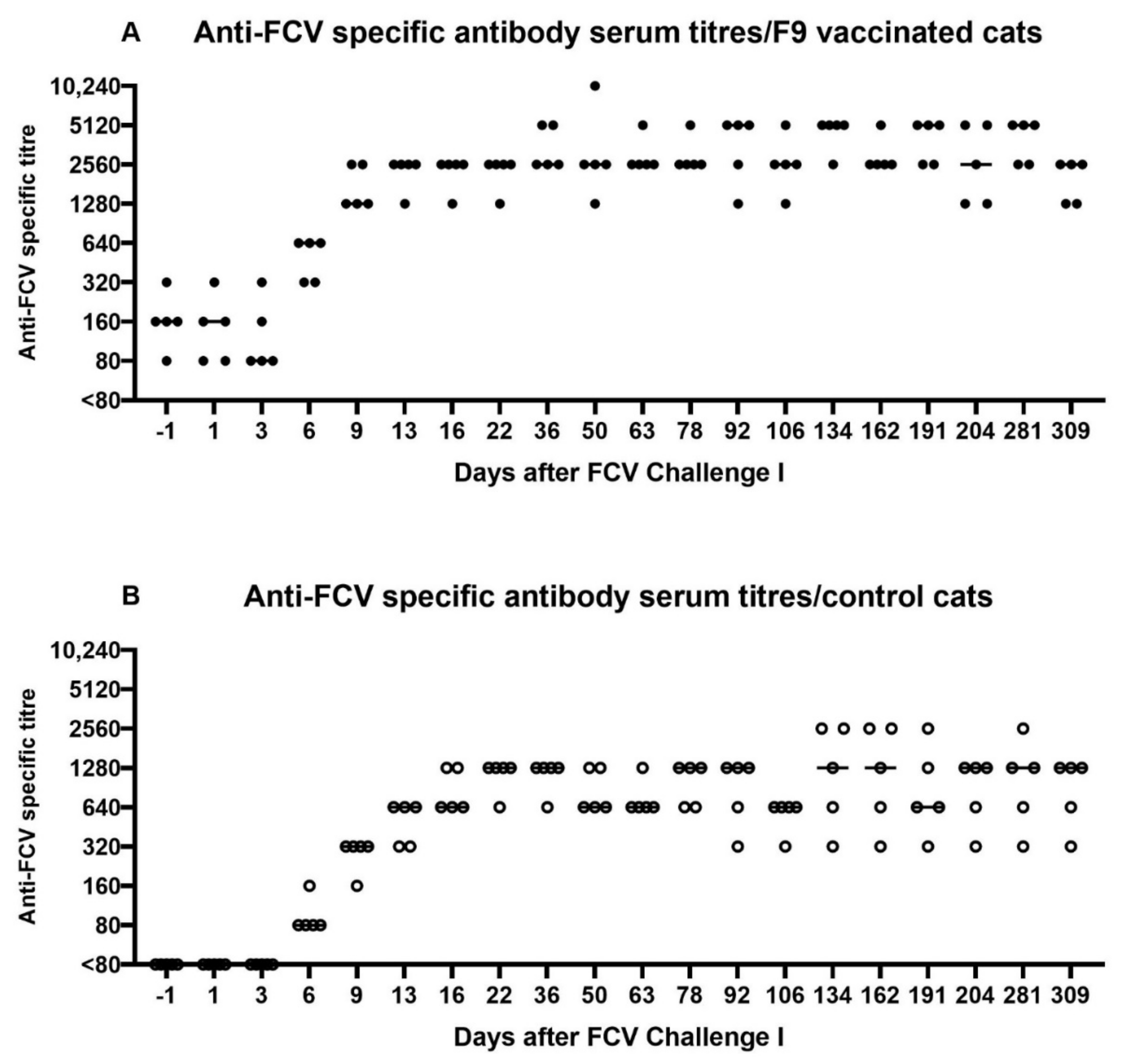
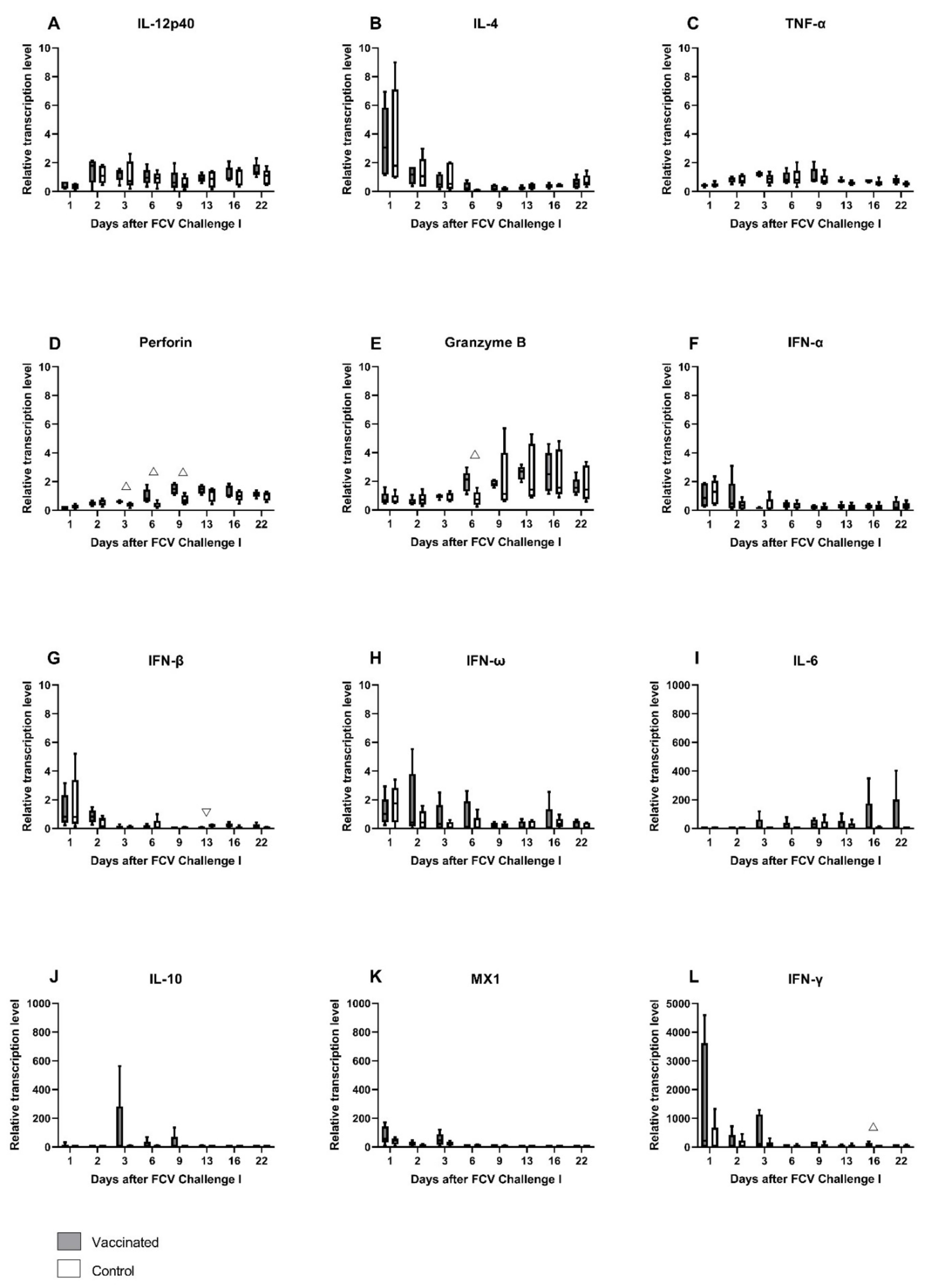
3.2. FCV Challenge I
3.3. FCV Vaccination II
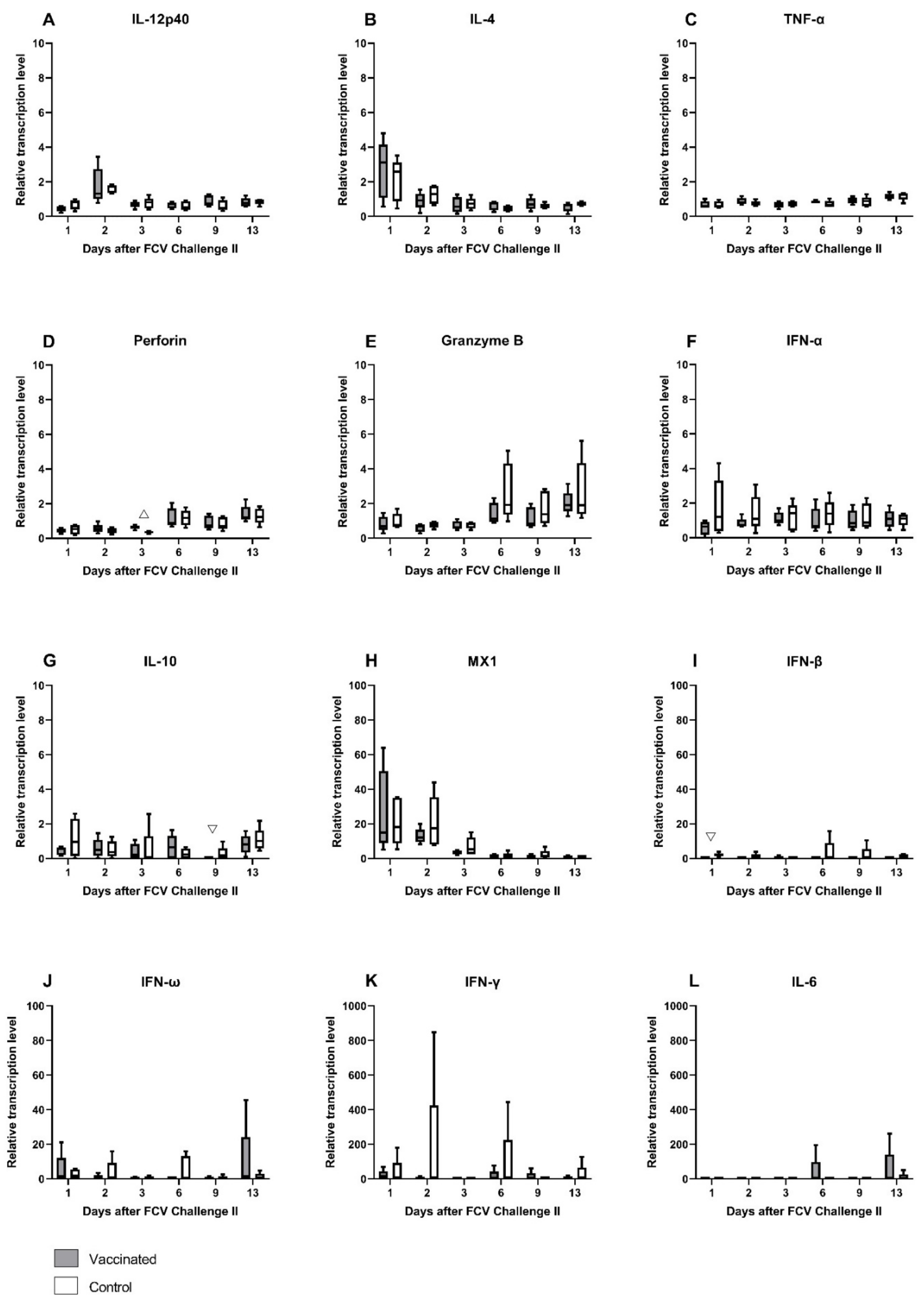
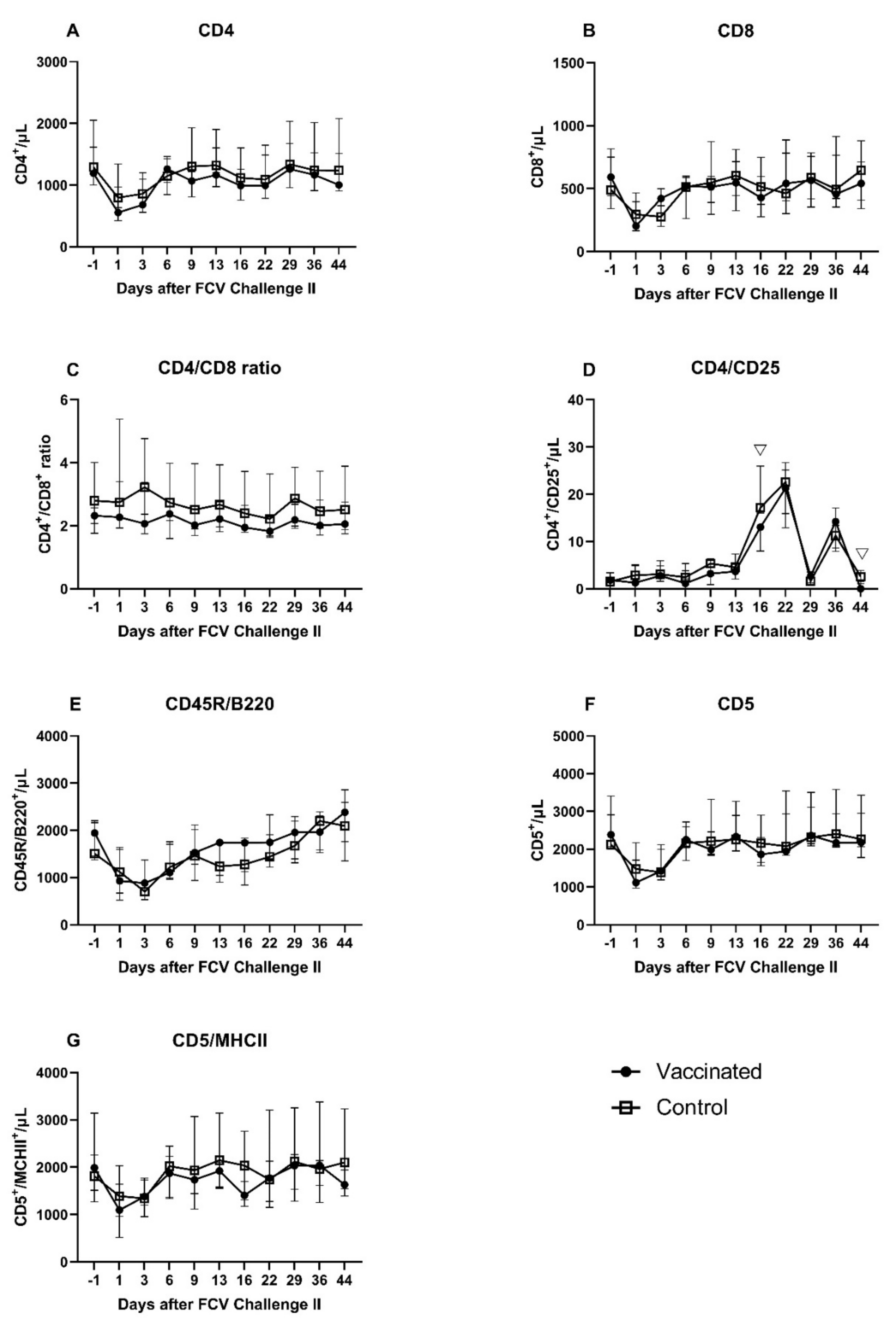
3.4. FCV Challenge II
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokine | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| IL-10 | PF = 0.0255 Day 20/Day 34 ↑ (PD = 0.0093) | PF = 0.032 Day 20/Day 34 ↑ (PD = 0.016) |
| MX1 | PF = 0.004 Day 4 ↑/Day 27 (PD = 0.0443) Day 4 ↑/Day 34 (PD = 0.0443) | Not significant |
| IFN-γ | PF = 0.0155 Day 13 ↑/Day 22 (PD = 0.0093) | Not significant |
| IL-12p40 | PF = 0.0077 Day 20 ↑/Day 34 (PD = 0.0016) | PF = 0.0037 Day 20 ↑/Day 34 (PD = 0.0009) |
| IL-4 | PF = 0.0067 Day 7/Day 22 ↑ (PD = 0.0016) | PF = 0.0003 Day 4/Day 22 ↑ (PD = 0.0093) Day 4/Day 27 ↑ (PD = 0.0443) Day 4/Day 34 ↑ (PD = 0.016) Day 7/Day 22 ↑ (PD = 0.0269) Day 7/Day 34 ↑ (PD = 0.0443) |
| IL-6 | - | - |
| TNF-α | PF = 0.0082 | PF = 0.0172 Day 13 ↑/Day 34 (PD = 0.0269) |
| IFN-ω | - | - |
| Perforin | PF = 0.044 Day 22/Day 27 ↑ (PD = 0.016) | PF = 0.029 Day 4 ↑/Day 34 (PD = 0.016) |
| Granzyme B | PF = 0.0272 | - |
| Lymphocyte Subset | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| CD5 | PF < 0.0001 Day 4↓/Day 27 (PD = 0.023) Day 7↓/Day 27 (PD = 0.0463) Day 13↓/Day 27 (PD = 0.0367) Day 200↓/Day 27 (PD = 0.0291) | PF = 0.0368 Day 48/Day 90↑ (PD = 0.0463) |
| CD45 | PF = 0.0001 Day −1↑/Day 22 (PD = 0.023) Day 20↑/Day 22 (PD = 0.0367) Day 34↑/Day 22 (PD = 0.0463) Day 4↓/Day 34 (PD = 0.0023) Day 200↓/Day 34 (PD = 0.0111) | PF = 0.0007 |
| CD4 | PF = 0.0001 Day 40↑/Day 200 (PD = 0.0142) Day 90↑/Day 200 (PD = 0.0463) | PF = 0.0038 Day 48/Day 90↑ (PD = 0.0367) |
| CD8 | PF < 0.0001 Day 4↓/Day 40 (PD = 0.0111) Day 7↓/Day 40 (PD = 0.0181) Day 13↓/Day 40 (PD = 0.0463) Day 22↓/Day 40 (PD = 0.0142) Day 200↓/Day 40 (PD = 0.023) | Not significant |
| CD4/CD8 ratio | PF = 0.0003 Day 4↑/Day 40 (PD = 0.0367) Day 7↑/Day 40 (PD = 0.0086) Day 13↑/Day 40 (PD = 0.0291) Day 22↑/Day 40 (PD = 0.023) | Not significant |
| IFN-γ-Releasing PBMCs (ELISpot) | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| PF = 0.0293 PD > 0.05 significance lost in the post test | PF > 0.05 |
| Cytokine | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| IFN-γ | PF = 0.036 Day 1 ↑/Day 13 (PD = 0.0221) | PF = 0.0022 Day 1↑/Day 6 (PD = 0.0221) Day1↑/Day 16 (PD = 0.0137) |
| IL-12p40 | PF = 0.0222 Day 1 ↓/Day 22 (PD = 0.0221) | PF = 0.0123 Day 1 ↓/Day 2 (PD = 0.0221) |
| IL-4 | PF = 0.0067 Day 1↑/Day 13 (PD = 0.0084) | PF = 0.0003 Day 1↑/Day 6 (PD = 0.0006) Day 1↑/Day 9 (PD = 0.035) Day 2↑/Day 6 (PD = 0.0137) |
| IL-10 | PF = 0.027 | PF = 0.0335 |
| TNF-α | PF = 0.0036 Day 1↓/Day 3 (PD = 0.0006) Day 1↓/Day 9 (PD = 0.035) | PF = 0.0126 |
| IFN-α | PF = 0.0387 Day 1↑/Day 3 (PD = 0.035) | PF = 0.0162 Day 1↑/Day 9 (PD = 0.035) Day 1↑/Day 16 (PD = 0.0084) |
| IFN-β | PF = 0.0003 Day 1↑/Day 13 (PD = 0.0137) Day 2↑/Day 3 (PD = 0.035) Day 2↑/Day 9 (PD = 0.035) Day 2↑/Day 13 (PD = 0.0084) | PF = 0.0063 Day 1/Day 9 (PD = 0.0221) |
| MX1 | PF < 0.0001 Day 1↑/Day 13 (PD = 0.0084) Day 1↑/Day 16 (PD = 0.0137) Day 1↑/Day 22 (PD = 0.0006) Day 3↑/Day 22 (PD = 0.0084) | PF < 0.0001 Day 1↑/Day 13 (PD = 0.0137) Day 1↑/Day 16 (PD = 0.0084) Day 1↑/Day 22 (PD = 0.001) Day 3↑/Dy 16 (PD = 0.035) Day 3↑/Day 22 (PD = 0.0051) |
| Perforin | PF < 0.0001 Day 1↓/Day 9 (PD = 0.0018) Day 1↓/Day 13 (PD = 0.0051) Day 2↓/Day 9 (PD = 0.0221) | PF = 0.0002 Day 1↓/Day 13 (PD = 0.0221) Day 1↓/Day 16 (PD = 0.0221) Day 1↓/Day 22 (PD = 0.0221) |
| Granzyme B | PF = 0.0005 Day 1↓/Day 13 (PD = 0.035) Day 2↓/Day 13 (PD = 0.0051) Day 2↓/Day16 (PD = 0.035) | PF = 0.0005 Day 1↓/Day 13 (PD = 0.0221) Day 6↓/Day 13 (PD = 0.035) |
| Lymphocyte Subset | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| CD5 | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0017) Day −1↑/Day 3 (PD = 0.008) Day −1↑/Day 6 (PD = 0.0336) Day −1↑/Day 162 (PD = 0.0409) Day 1↓/Day 78 (PD = 0.0034) Day 3↓/Day 78 (PD = 0.0151) | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0496) Day 1↓/Day 22 (PD = 0.0004) Day 3↓/Day 22 (PD = 0.0027) Day 6↓/Day 22 (PD = 0.0014) Day 1↓/Day 78 (PD = 0.0409) |
| CD5/MHCII | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0017) Day −1↑/Day 3 (PD = 0.0151) Day −1↑/Day 6 (PD = 0.0409) Day −1↑/Day 162 (PD = 0.0276) Day 78↑/Day 1 (PD = 0.0226) Day 1↓/Day 50 (PD = 0.0496) Day 1↓/Day 56 (PD = 0.0336) Day 1↓/Day 106 (PD = 0.0099) | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0496) Day 1↓/Day 22 (PD = 0.0007) Day 3↓/Day 22 (PD = 0.0042) Day 6↓/Day 22 (PD = 0.0022 Day 1↓/Day 63 (PD = 0.0496) |
| CD45 | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0226) Day −1↑/Day 3 (PD = 0.0151) Day 1↓/Day 29 (PD = 0.008) Day 1↓/Day 43 (PD = 0.0122) Day 1↓/Day 50 (PD = 0.0099 Day 1↓/Day 63 (PD = 0.0409 Day 1↓/Day 78 (PD = 0.0226 Day 3↓/Day 29 (PD = 0.0052 Day 3↓/Day 43 (PD = 0.008) Day 3↓/Day 50 (PD = 0.0065) Day 3↓/Day 63 (PD = 0.0276) Day 3↓/Day 78 (PD = 0.0151) Day 6↓/Day 29 (PD = 0.0336) Day 6↓/Day 43 (PD = 0.0496) Day 6↓/Day 50 (PD = 0.0409) | PF < 0.0001 Day 1↓/Day 50 (PD = 0.0122) Day 1↓/Day 56 (PD = 0.008) Day 1↓/Day 63 (PD = 0.0151) Day 3↓/Day 50 (PD = 0.0276) Day 3↓/Day 56 (PD = 0.0185) Day 3↓/Day 63 (PD = 0.0336) Day 6↓/Day 50 (PD = 0.0099) Day 6↓/Day 56 (PD = 0.0065) Day 6↓/Day 63 (PD = 0.0122) |
| CD4 | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0011) Day −1↑/Day 3 (PD = 0.0052) Day −1↑/Day 6 (PD = 0.0276) Day −1↑/Day 162 (PD = 0.0226) Day 1↓/Day 78 (PD = 0.008) Day 1↓/Day 106 (PD = 0.0226) Day 3↓/Day 78 (PD = 0.0336) | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0226) Day 1↓/Day 22 (PD = 0.0007) Day 3↓/Day 22 (PD = 0.0042) Day 6↓/Day 22 (PD = 0.0022) Day 1↓/Day 78 (PD = 0.0226) |
| CD4/CD25 | PF < 0.0001 Day 6↓/Day 13 (PD = 0.0099) Day 6↓/Day 16 (PD = 0.0122) Day 6↓/Day 29 (PD = 0.0034) Day 6↓/Day 106 (PD = 0.0496) Day 6↓/Day 162 (PD = 0.0276) Day 13↑/Day 63 (PD = 0.0496) Day 29↑/Day 63 (PD = 0.0185) Day 29↑/Day 71 (PD = 0.0409) | PF < 0.0001 Day 1↓/Day 13 (PD = 0.0042) Day 1↓/Day 16 (PD = 0.0014) Day 1↓/Day 29 (PD = 0.0042) Day 1↓/Day 162 (PD = 0.0052) Day 6↓/Day 13 (PD = 0.0017) Day 6↓/Day 16 (PD = 0.0005) Day 6↓/Day 29 (PD = 0.0017) Day 6↓/Day 85 (PD = 0.0496) Day 6↓/Day 162 (PD = 0.0022) Day 16↑/Day 71 (PD = 0.0226) |
| CD8 | PF < 0.0001 Day −1↑/Day 1 (PD = 0.0027) Day −1↑/Day 3 (PD = 0.008) Day −1↑/Day 6 (PD = 0.0496) Day 50↑/Day 1 (PD = 0.0276) Day 78↑/Day 1(PD = 0.0042) Day 78↑/Day 3 (PD = 0.0122) | PF < 0.0001 Day 22↑/Day 1 (PD = 0.0002) Day 22↑/Day 3 (PD = 0.0009) Day 22↑/Day 6 (PD = 0.0005) Day 22↑/Day 9 (PD = 0.0151) Day 22↑/Day 162 (PD = 0.0151) |
| CD4/CD8 ratio | PF = 0.0003 Day 3↑/Day 29 (PD = 0.0042) Day 3↑/Day 43 (PD = 0.008) Day 3↑/Day 50 (PD = 0.0336) | PF < 0.0001 Day3↑/Day 22 (PD = 0.0226) Day 3↑/Day 29 (PD = 0.0122) Day 3↑/Day 36 (PD = 0.0409) Day 3↑/Day 43 (PD = 0.0276) Day 6↑/Day 22 (PD = 0.0185) Day 6↑/Day 29 (PD = 0.0099) Day 6↑/Day 36 (PD = 0.0336) Day 6↑/Day 43 (PD = 0.0226) Day 6↑/Day 134 (PD = 0.0151) |
| IFN-γ-Releasing PBMCs (ELISpot) | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| No Friedman and Dunn’s post test possible (missing value) | PF = 0.0001 Day −5↓/Day 55 (PD = 0.0051) Day −5↓/Day 204 (PD = 0.035) Day 2↓/Day 55 (PD = 0.0018) Day 2↓/Day 204 (PD = 0.0137) |
| Cytokine | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| IL-12p40 | PF = 0.0037 Day 1↓/Day2 (PD = 0.0007) | PF = 0.0234 Day 2↑/Day 9 (PD = 0.0352) |
| IL-4 | PF = 0.0134 Day 1↑/Day 13 (PD = 0.0108) | PF = 0.0076 Day 1↑/Day 6 (PD = 0.0108) Day 2↑/Day 6 (PD = 0.0198) |
| IL-6 | PF = 0.008 Day 1↓/Day 13 (PD = 0.0352) Day 2↓/Day 13 (PD = 0.0058) | PF = 0.0213 Day 2↓/Day 13 (PD = 0.0058) |
| IL-10 | PF = 0.0421 Day 9↓/Day 13 (PD = 0.0198) | Not significant |
| TNF-α | PF = 0.0088 Day 1↓/Day 13 (PD = 0.0198) Day 3↓/Day 13 (PD = 0.0198) | Not significant |
| IFN-β | Not significant | Not significant |
| MX1 | PF = 0.0009 Day 1↑/Day 9 (PD = 0.0198) Day 1↑/Day 13 (PD = 0.0352) Day 2↑/Day 9 (PD = 0.0352) | PF = 0.0005 Day 1↑/Day 6 (PD = 0.0108) Day 1↑/Day 13 (PD = 0.0108) Day 2↑/Day 6 (PD = 0.0198) Day 2↑/Day 13 (PD = 0.0198) |
| Perforin | PF = 0.0005 Day 1↓/Day 6 (PD = 0.0198) Day 1↓/Day 13 (PD = 0.0030) Day 2↓/Day 13 (PD = 0.0352) | PF = 0.0009 Day 2↓/Day 13(PD = 0.0198) Day 3↓/Day 13 (PD = 0.0030) |
| Granzyme B | PF = 0.0106 Day 2↓/Day 13 (PD = 0.0108) | PF = 0.0024 Day 3↓/Day 6 (PD = 0.0108) Day 3↓/Day 13 (PD = 0.0352) |
| Lymphocyte Subset | Change over Time (Friedman and Dunn’s Post Test) Vaccinated Group | Change over Time (Friedman and Dunn’s Post Test) Control Group |
|---|---|---|
| CD5 | PF = 0.0002 Day 1↓/Day 29 (PD = 0.0075) Day 3↓/Day 29 (PD = 0.011) | PF = 0.0006 Day 1↓/Day 29 (PD = 0.0329) Day 3↓/Day 29 (PD = 0.011) Day 3↓/Day 36 (PD = 0.0466) |
| CD5/MHCII | PF = 0.0007 Day 1↓/Day 13 (PD = 0.0329) Day 1↓/Day 29 (PD = 0.0466) | PF = 0.0046 Significance was lost in the post test |
| CD45 | PF = 0.0001 Day −1↑/Day 3 (PD = 0.0329) Day 3↓/Day 36 (PD = 0.023) Day 3↓/Day 44 (PD = 0.011) | PF < 0.0001 Day −1↑/Day 3 (PD = 0.0329) Day 1↓/Day 36 (PD = 0.0329) Day 1↓/Day 44 (PD = 0.0034) Day 3↓/Day 36 (PD = 0.0034) Day 3↓/Day 44 (PD = 0.0003) Day 6↓/Day 44 (PD = 0.0466) Day 13↓/Day 44 (PD = 0.0466) |
| CD4 | PF = 0.0002 Day −1↑/Day 1 (PD = 0.0466) Day −1↑/Day 3 (PD = 0.016) Day 1↓/Day 29 (PD = 0.023) Day 3↓/Day 13 (PD = 0.0329) Day 3↓/Day 29 (PD = 0.0075) | PF = 0.0002 Day 1↓/Day 29 (PD = 0.0329) Day 3↓/Day 29 (PD = 0.0075) Day 3↓/Day 44 (PD = 0.023) |
| CD4/CD25 | PF = 0.0004 Day 1↓/Day 22 (PD = 0.0466) Day 22↑/Day 44 (PD = 0.0034) | PF < 0.0001 Day −1↓/Day 16 (PD = 0.0051) Day −1↓/Day 22 (PD = 0.0075) Day 16↑/Day 29 (PD = 0.0329) Day 22↑/Day 29 (PD = 0.0466) |
| CD8 | PF = 0.0029 Day −1↑/Day 1 (PD = 0.023) | PF < 0.0001 Day 1↓/Day 36 (PD = 0.0466) Day 1↓/Day 44 (PD = 0.016) Day 3↓/Day 36 (PD = 0.0329) Day 3↓/Day 44 (PD = 0.011) |
| CD4/CD8 ratio | PF = 0.0314 Day 1↑/Day 22 (PD = 0.0034) | PF = 0.0027 Day 1↑/Day 22 (PD = 0.0466) Day 3↑/Day 22 (PD = 0.0023) |
| IFN-γ-Releasing PBMCs (ELISpot) | Change over Time (Mann–Whitney U) Vaccinated Group | Change over Time (Mann–Whitney U) Control Group |
|---|---|---|
| Not significant | Not significant |
References
- Radford, A.D.; Coyne, K.P.; Dawson, S.; Porter, C.J.; Gaskell, R.M. Feline calicivirus. Vet. Res. 2007, 38, 319–335. [Google Scholar] [CrossRef] [Green Version]
- Radford, A.D.; Sommerville, L.; Ryvar, R.; Cox, M.B.; Johnson, D.R.; Dawson, S.; Gaskell, R.M. Endemic infection of a cat colony with a feline calicivirus closely related to an isolate used in live attenuated vaccines. Vaccine 2001, 19, 4358–4362. [Google Scholar] [CrossRef]
- Radford, A.D.; Dawson, S.; Ryvar, R.; Coyne, K.; Johnson, D.R.; Cox, M.B.; Acke, E.F.; Addie, D.D.; Gaskell, R.M. High Genetic Diversity of the Immunodominant Region of the Feline Calicivirus Capsid Gene in Endemically Infected Cat Colonies. Virus Genes 2003, 27, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Bannasch, M.J.; Foley, J.E. Epidemiologic evaluation of multiple respiratory pathogens in cats in animal shelters. J. Feline Med. Surg. 2005, 7, 109–119. [Google Scholar] [CrossRef]
- Helps, C.R.; Lait, P.; Damhuis, A.; Björnehammar, U.; Bolta, D.; Brovida, C.; Chabanne, L.; Egberink, H.; Ferrand, G.; Fontbonne, A.; et al. Factors associated with upper respiratory tract disease caused by feline herpesvirus, feline calicivirus, Chlamydophila felis and Bordetella bronchiseptica in cats: Experience from 218 European catteries. Vet. Rec. 2005, 156, 669–673. [Google Scholar] [CrossRef] [Green Version]
- Coyne, K.P.; Dawson, S.; Radford, A.; Cripps, P.J.; Porter, C.J.; McCracken, C.M.; Gaskell, R.M. Long-term analysis of feline calicivirus prevalence and viral shedding patterns in naturally infected colonies of domestic cats. Vet. Microbiol. 2006, 118, 12–25. [Google Scholar] [CrossRef]
- Berger, A.; Willi, B.; Meli, M.L.; Boretti, F.S.; Hartnack, S.; Dreyfus, A.; Lutz, H.; Hofmann-Lehmann, R. Feline calicivirus and other respiratory pathogens in cats with Feline calicivirus-related symptoms and in clinically healthy cats in Switzerland. BMC Vet. Res. 2015, 11, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radford, A.D.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; et al. Feline calicivirus infection. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 556–564. [Google Scholar] [CrossRef] [PubMed]
- Willi, B.; Spiri, A.M.; Meli, M.L.; Samman, A.; Hoffmann, K.; Sydler, T.; Cattori, V.; Graf, F.; Diserens, K.A.; Padrutt, I.; et al. Molecular characterization and virus neutralization patterns of severe, non-epizootic forms of feline calicivirus infections resembling virulent systemic disease in cats in Switzerland and in Liechtenstein. Vet. Microbiol. 2016, 182, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, N.; Elliott, J.; Glasgow, A.; Poland, A.; Keel, K. An isolated epizootic of hemorrhagic-like fever in cats caused by a novel and highly virulent strain of feline calicivirus. Vet. Microbiol. 2000, 73, 281–300. [Google Scholar] [CrossRef]
- Hurley, K.F.; Pesavento, P.A.; Pedersen, N.C.; Poland, A.M.; Wilson, E.; Foley, J.E. An outbreak of virulent systemic feline calicivirus disease. J. Am. Vet. Med. Assoc. 2004, 224, 241–249. [Google Scholar] [CrossRef]
- Coyne, K.P.; Christley, R.; Pybus, O.; Dawson, S.; Gaskell, R.M.; Radford, A. Large-Scale Spatial and Temporal Genetic Diversity of Feline Calicivirus. J. Virol. 2012, 86, 11356–11367. [Google Scholar] [CrossRef] [Green Version]
- Radford, A.D.; Dawson, S.; Coyne, K.P.; Porter, C.J.; Gaskell, R.M. The challenge for the next generation of feline calicivirus vaccines. Vet. Microbiol. 2006, 117, 14–18. [Google Scholar] [CrossRef]
- Hurley, K.F.; Sykes, E.J. Update on feline calicivirus: New trends. Vet. Clin. N. Am. Small Anim. Pr. 2003, 33, 759–772. [Google Scholar] [CrossRef]
- Schorr-Evans, E.; Poland, A.; Johnson, W.; Pedersen, N. An epizootic of highly virulent feline calicivirus disease in a hospital setting in New England. J. Feline Med. Surg. 2003, 5, 217–226. [Google Scholar] [CrossRef]
- Povey, C.; Ingersoll, J. Cross-protection among feline caliciviruses. Infect. Immun. 1975, 11, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulet, H.; Brunet, S.; Leroy, V.; Chappuis, G. Immunisation with a combination of two complementary feline calicivirus strains induces a broad cross-protection against heterologous challenges. Vet. Microbiol. 2005, 106, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Poulet, H.; Jas, D.; Lemeter, C.; Coupier, C.; Brunet, S. Efficacy of a bivalent inactivated non-adjuvanted feline calicivirus vaccine: Relation between in vitro cross-neutralization and heterologous protection in vivo. Vaccine 2008, 26, 3647–3654. [Google Scholar] [CrossRef] [PubMed]
- Lesbros, C.; Martin, V.; Najbar, W.; Sanquer, A.; McGahie, D.; Eun, H.-M.; Gueguen, S. Protective Efficacy of the Calicivirus Valency of the Leucofeligen Vaccine against a Virulent Heterologous Challenge in Kittens. Vet. Med. Int. 2013, 2013, 232397. [Google Scholar] [CrossRef]
- Sommerville, L.M.; Radford, A.D.; Glenn, M.; Dawson, S.; Gaskell, C.J.; Kelly, D.F.; Cripps, P.J.; Porter, C.J.; Gaskell, R.M. DNA vaccination against feline calicivirus infection using a plasmid encoding the mature capsid protein. Vaccine 2002, 20, 1787–1796. [Google Scholar] [CrossRef]
- Lauritzen, A.; Jarrett, O.; Sabara, M. Serological analysis of feline calicivirus isolates from the United States and United Kingdom. Vet. Microbiol. 1997, 56, 55–63. [Google Scholar] [CrossRef]
- Addie, D.; Poulet, H.; Golder, M.C.; McDonald, M.; Brunet, S.; Thibault, J.-C.; Hosie, M.J. Ability of antibodies to two new caliciviral vaccine strains to neutralise feline calicivirus isolates from the uk. Vet. Rec. 2008, 163, 355–357. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.J.; Radford, A.D.; Gaskell, R.M.; Ryvar, R.; Coyne, K.P.; Pinchbeck, G.L.; Dawson, S. Comparison of the ability of feline calicivirus (FCV) vaccines to neutralise a panel of current UK FCV isolates. J. Feline Med. Surg. 2008, 10, 32–40. [Google Scholar] [CrossRef]
- Smith, S.L.; Afonso, M.M.; Pinchbeck, G.L.; Gaskell, R.M.; Dawson, S.; Radford, A.D. Temporally separated feline calicivirus isolates do not cluster phylogenetically and are similarly neutralised by high-titre vaccine strain FCV-F9 antisera in vitro. J. Feline Med. Surg. 2019, 22, 602–607. [Google Scholar] [CrossRef]
- Tham, K.M.; Studdert, M.J. Antibody and Cell-mediated Immune Responses to Feline Calicivirus Following Inactivated Vaccine and Challenge. J. Vet. Med. Ser. B 1987, 34, 640–654. [Google Scholar] [CrossRef]
- De Groot-Mijnes, J.D.; Van Der Most, R.G.; Van Dun, J.M.; Lintelo, E.G.T.; Schuurman, N.M.; Egberink, H.; De Groot, R.J. Three-color flow cytometry detection of virus-specific CD4+ and CD8+ T cells in the cat. J. Immunol. Methods 2004, 285, 41–54. [Google Scholar] [CrossRef]
- Spiri, A.M.; Meli, M.L.; Riond, B.; Herbert, I.; Hosie, M.J.; Hofmann-Lehmann, R. Environmental Contamination and Hygienic Measures After Feline Calicivirus Field Strain Infections of Cats in a Research Facility. Viruses 2019, 11, 958. [Google Scholar] [CrossRef] [Green Version]
- Spiri, A.; Riond, B.; Stirn, M.; Novacco, M.; Meli, M.; Boretti, F.; Herbert, I.; Hosie, M.; Hofmann-Lehmann, R. Modified-Live Feline Calicivirus Vaccination Reduces Viral RNA Loads, Duration of RNAemia, and the Severity of Clinical Signs after Heterologous Feline Calicivirus Challenge. Viruses 2021, 13, 1505. [Google Scholar] [CrossRef]
- Brunner, C.; Kanellos, T.; Meli, M.L.; Sutton, D.J.; Gisler, R.; Gomes-Keller, M.A.; Hofmann-Lehmann, R.; Lutz, H. Antibody induction after combined application of an adjuvanted recombinant FeLV vaccine and a multivalent modified live virus vaccine with a chlamydial component. Vaccine 2006, 24, 1838–1846. [Google Scholar] [CrossRef]
- Felten, S.; Klein-Richers, U.; Hofmann-Lehmann, R.; Bergmann, M.; Unterer, S.; Leutenegger, C.M.; Hartmann, K. Correlation of Feline Coronavirus Shedding in Feces with Coronavirus Antibody Titer. Pathogens 2020, 9, 598. [Google Scholar] [CrossRef] [PubMed]
- Leutenegger, C.M.; Mislin, C.N.; Sigrist, B.; Ehrengruber, M.U.; Hofmann-Lehmann, R.; Lutz, H. Quantitative real-time PCR for the measurement of feline cytokine mRNA. Vet. Immunol. Immunopathol. 1999, 71, 291–305. [Google Scholar] [CrossRef] [Green Version]
- Robert-Tissot, C.; Rüegger, V.L.; Cattori, V.; Meli, M.L.; Riond, B.; Gomes-Keller, M.A.; Vögtlin, A.; Wittig, B.; Juhls, C.; Hofmann-Lehmann, R.; et al. The innate antiviral immune system of the cat: Molecular tools for the measurement of its state of activation. Vet. Immunol. Immunopathol. 2011, 143, 269–281. [Google Scholar] [CrossRef]
- Kessler, Y.; Helfer-Hungerbuehler, A.K.; Cattori, V.; Meli, M.L.; Zellweger, B.; Ossent, P.; Riond, B.; Reusch, E.C.; Lutz, H.; Hofmann-Lehmann, R. Quantitative TaqMan® real-time PCR assays for gene expression normalisation in feline tissues. BMC Mol. Biol. 2009, 10, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [Green Version]
- Ackley, C.O.; Hoover, L.A.; Cooper, M.D. Identification of a CD4 homologue in the cat. Tissue Antigens 1990, 35, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Ackley, C.D.; Cooper, M.D. Characterization of a feline T-cell-specific monoclonal antibody reactive with a CD5-like molecule. Am. J. Vet. Res. 1992, 53, 466–471. [Google Scholar]
- Monteith, E.C.; Chelack, B.J.; Davis, W.C.; Haines, D.M. Identification of monoclonal antibodies for immunohistochemical staining of feline B lymphocytes in frozen and formalin-fixed paraffin-embedded tissues. Can. J. Vet. Res. Rev. Can. Rech. Vet. 1996, 60, 193–198. [Google Scholar]
- Vahlenkamp, T.W.; Tompkins, M.B.; Tompkins, W.A. The role of CD4+CD25+ regulatory T cells in viral infections. Vet. Immunol. Immunopathol. 2005, 108, 219–225. [Google Scholar] [CrossRef]
- Hicks, C.; Willi, B.; Riond, B.; Novacco, M.; Meli, M.L.; Stokes, C.R.; Helps, C.; Hofmann-Lehmann, R.; Tasker, S. Protective Immunity against Infection with Mycoplasma haemofelis. Clin. Vaccine Immunol. 2015, 22, 108–118. [Google Scholar] [CrossRef] [Green Version]
- Novacco, M.; Boretti, F.S.; Franchini, M.; Riond, B.; Meli, M.L.; Hofmann-Lehmann, R. Protection from reinfection in “Candidatus Mycoplasma turicensis”-infected cats and characterization of the immune response. Vet. Res. 2012, 43, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holznagel, E.; Hofmann-Lehmann, R.; Leutenegger, C.M.; Allenspach, K.; Huettner, S.; Forster, U.; Niederer, E.; Joller, H.; Willett, B.J.; Hummel, U.; et al. The role of in vitro-induced lymphocyte apoptosis in feline immunodeficiency virus infection: Correlation with different markers of disease progression. J. Virol. 1998, 72, 9025–9033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lappin, M.R.; Andrews, J.; Simpson, D.; Jensen, W.A. Use of serologic tests to predict resistance to feline herpesvirus 1, feline calicivirus, and feline parvovirus infection in cats. J. Am. Vet. Med. Assoc. 2002, 220, 38–42. [Google Scholar] [CrossRef]
- Jas, D.; Aeberlé, C.; Lacombe, V.; Guiot, A.; Poulet, H. Onset of immunity in kittens after vaccination with a non-adjuvanted vaccine against feline panleucopenia, feline calicivirus and feline herpesvirus. Vet. J. 2009, 182, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Jas, D.; Frances-Duvert, V.; Vernes, D.; Guigal, P.-M.; Poulet, H. Three-year duration of immunity for feline herpesvirus and calicivirus evaluated in a controlled vaccination-challenge laboratory trial. Vet. Microbiol. 2015, 177, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lappin, M.R.; Veir, J.; Hawley, J. Feline panleukopenia virus, feline herpesvirus-1, and feline calicivirus antibody responses in seronegative specific pathogen-free cats after a single administration of two different modified live FVRCP vaccines. J. Feline Med. Surg. 2009, 11, 159–162. [Google Scholar] [CrossRef]
- Day, M.J.; Horzinek, M.C.; Schultz, R.D.; Squires, R.A. WSAVA Guidelines for the vaccination of dogs and cats. J. Small Anim. Pr. 2016, 57, E1–E45. [Google Scholar] [CrossRef] [Green Version]
- DiGangi, B.; Levy, J.K.; Griffin, B.; Reese, M.J.; Dingman, A.P.; Tucker, S.J.; Dubovi, E.J. Effects of maternally-derived antibodies on serologic responses to vaccination in kittens. J. Feline Med. Surg. 2012, 14, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Metwally, S.; Viljoen, G.; El Idrissi, A. Veterinary Vaccines; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Afonso, M.M.; Pinchbeck, G.L.; Smith, S.L.; Daly, J.M.; Gaskell, R.M.; Dawson, S.; Radford, A.D. A multi-national European cross-sectional study of feline calicivirus epidemiology, diversity and vaccine cross-reactivity. Vaccine 2017, 35, 2753–2760. [Google Scholar] [CrossRef]
- Bergmann, M.; Speck, S.; Rieger, A.; Truyen, U.; Hartmann, K. Antibody Response to Feline Calicivirus Vaccination in Healthy Adult Cats. Viruses 2019, 11, 702. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, M.; Schwertler, S.; Reese, S.; Speck, S.; Truyen, U.; Hartmann, K. Antibody response to feline panleukopenia virus vaccination in healthy adult cats. J. Feline Med. Surg. 2018, 20, 1087–1093. [Google Scholar] [CrossRef]
- Vatti, A.; Monsalve, D.M.; Pacheco, Y.; Chang, C.; Anaya, J.-M.; Gershwin, M.E. Original antigenic sin: A comprehensive review. J. Autoimmun. 2017, 83, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Malm, M.; Vesikari, T.; Blazevic, V. Simultaneous Immunization with Multivalent Norovirus VLPs Induces Better Protective Immune Responses to Norovirus than Sequential Immunization. Viruses 2019, 11, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef]
- Veugelers, K.; Motyka, B.; Goping, I.S.; Shostak, I.; Sawchuk, T.; Bleackley, R.C. Granule-mediated Killing by Granzyme B and Perforin Requires a Mannose 6-Phosphate Receptor and Is Augmented by Cell Surface Heparan Sulfate. Mol. Biol. Cell 2006, 17, 623–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, G.N. Host defense, viruses and apoptosis. Cell Death Differ. 2001, 8, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Danthi, P. Viruses and the Diversity of Cell Death. Annu. Rev. Virol. 2016, 3, 533–553. [Google Scholar] [CrossRef]
- Haller, O.; Staeheli, P.; Schwemmle, M.; Kochs, G. Mx GTPases: Dynamin-like antiviral machines of innate immunity. Trends Microbiol. 2015, 23, 154–163. [Google Scholar] [CrossRef]
- Verhelst, J.; Parthoens, E.; Schepens, B.; Fiers, W.; Saelens, X. Interferon-Inducible Protein Mx1 Inhibits Influenza Virus by Interfering with Functional Viral Ribonucleoprotein Complex Assembly. J. Virol. 2012, 86, 13445–13455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiner, F. Antiviral Function of Mx Proteins: Role of Cellular Factors and Subcellular Localization. Ph.D. Thesis, University of Zurich, Zurich, Switzerland, 2018. [Google Scholar]
- Robert-Tissot, C.; Rüegger, V.L.; Cattori, V.; Meli, M.L.; Riond, B.; Moore, P.F.; Engels, M.; Franchini, M.; Hofmann-Lehmann, R.; Lutz, H. Stimulation with a class A CpG oligonucleotide enhances resistance to infection with feline viruses from five different families. Vet. Res. 2012, 43, 60. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Zhang, X.; Wu, H.; Liu, C.; Liu, J.; Hu, X.; Qu, L. Assessment of the IFN-β response to four feline caliciviruses: Infection in CRFK cells. Infect. Genet. Evol. 2015, 34, 352–360. [Google Scholar] [CrossRef]
- Vermeulen, B.L.; Gleich, S.E.; Dedeurwaerder, A.; Olyslaegers, D.A.; Desmarets, L.M.; Dewerchin, H.L.; Nauwynck, H.J. In vitro assessment of the feline cell-mediated immune response against feline panleukopeniavirus, calicivirus and felid herpesvirus 1 using 5-bromo-2′-deoxyuridine labeling. Vet. Immunol. Immunopathol. 2012, 146, 177–184. [Google Scholar] [CrossRef]
- Yumiketa, Y.; Narita, T.; Inoue, Y.; Sato, G.; Kamitani, W.; Oka, T.; Katayama, K.; Sakaguchi, T.; Tohya, Y. Nonstructural protein p39 of feline calicivirus suppresses host innate immune response by preventing IRF-3 activation. Vet. Microbiol. 2016, 185, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zu, S.; Sun, X.; Liu, Y.; Tian, J.; Qu, L. N-Terminal Domain of Feline Calicivirus (FCV) Proteinase-Polymerase Contributes to the Inhibition of Host Cell Transcription. Viruses 2016, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Dolieslager, S.M.J.; Lappin, D.F.; Bennett, D.; Graham, L.; Johnston, N.; Riggio, M.P. The influence of oral bacteria on tissue levels of Toll-like receptor and cytokine mRNAs in feline chronic gingivostomatitis and oral health. Vet. Immunol. Immunopathol. 2013, 151, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.; Hurley, K.; Pesavento, P.A.; Poland, A.; Pedersen, N.C. Virulent systemic feline calicivirus infection: Local cytokine modulation and contribution of viral mutants. J. Feline Med. Surg. 2006, 8, 55–61. [Google Scholar] [CrossRef]
- Campbell, D.; Rawlings, J.; Koelsch, S.; Wallace, J.; Strain, J.; Hannigan, B. Age-related differences in parameters of feline immune status. Vet. Immunol. Immunopathol. 2004, 100, 73–80. [Google Scholar] [CrossRef]
- Knowles, J.; Macardle, F.; Dawson, S.; Carter, S.; Gaskell, C.; Gaskell, R. Studies on the role of feline calicivirus in chronic stomatitis in cats. Vet. Microbiol. 1991, 27, 205–219. [Google Scholar] [CrossRef]

















| FCV Vaccination I * | FCV Challenge I | FCV Vaccination II | FCV Challenge II | |
|---|---|---|---|---|
| Flow cytometry | −1, 4, 7, 13, 20, 22, 27, 34, 40, 48, 55, 62, 69, 76, 82, 90, 96, and 200 after injection I | −1, 1, 3, 6, 9, 13, 16, 22, 29, 36, 43, 50, 56, 63, 71, 78, 85, 92, 99, 106, 134, and 162 | Not performed | −1, 1, 3, 6, 9, 13, 16, 22, 29, 36, and 44 |
| Cytokines | −1, 4, 7, 13, 20, 22, 27, and 34 after injection I | −1, 1, 2, 3, 6, 9, 13, 16, and 22 | Not performed | −1, 1, 2, 3, 6, 9, and 13 |
| IFA | −24, 4, 7, 13, 27, 34, 40, 55, 69, 82, 96, 111, 133, 153, and 174 after injection I | −1, 1, 3, 6, 9, 13, 16, 22, 36, 50, 63, 78, 92, 106, 134, 162, 191, 204, 281, and 309 | −1, 1, 2, 3, 8, and 34 | −1, 3, 29, and 56 |
| Virus neutralisation | −1, 13, 34, and 214 after injection I | −1, 6, 13, and 154 | −1 and 34 | −1 and 36 |
| PBMC collection | −29, 22, and 92 after injection I | −5, 2, 8, 24, 38, 55, 113, and 204 | Not performed | −4 and 38 |
| Day 154 after FCV Challenge I/Endpoint Titration | ||||
|---|---|---|---|---|
| Cat ID | FCV F9 | FCV 273 | FCV 27 | |
| Vaccinated | JJG4 | 1215 | 1215 | 135 |
| JJG6 | 1215 | 1215 | 135 | |
| JJH3 | 3645 | 3645 | 135 | |
| JJI1 | 10,935 | 32,805 | 3645 | |
| JJI2 | >98,415 | 3645 | 1215 | |
| Control | JJF1 | 135 | 10,935 | 1215 |
| JJG3 | 15 | 1215 | 45 | |
| JJH2 | 3645 | 32,805 | 405 | |
| JJI3 | 405 | 10,935 | 1215 | |
| JJI4 | 135 | 10,935 | 45 | |
| Day 36 after FCV Challenge II/Endpoint Titration | ||
|---|---|---|
| Cat ID | FCV 27 | |
| Vaccinated | JJG4 | 3645 |
| JJG6 | 3645 | |
| JJH3 | 1215 | |
| JJI1 | 3645 | |
| JJI2 | 1215 | |
| Control | JJF1 | 10,935 |
| JJG3 | 3645 | |
| JJH2 | 3645 | |
| JJI3 | 98,415 | |
| JJI4 | 1215 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spiri, A.M.; Novacco, M.; Meli, M.L.; Stirn, M.; Riond, B.; Fogle, J.E.; Boretti, F.S.; Herbert, I.; Hosie, M.J.; Hofmann-Lehmann, R. Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study. Viruses 2021, 13, 1736. https://doi.org/10.3390/v13091736
Spiri AM, Novacco M, Meli ML, Stirn M, Riond B, Fogle JE, Boretti FS, Herbert I, Hosie MJ, Hofmann-Lehmann R. Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study. Viruses. 2021; 13(9):1736. https://doi.org/10.3390/v13091736
Chicago/Turabian StyleSpiri, Andrea M., Marilisa Novacco, Marina L. Meli, Martina Stirn, Barbara Riond, Jonathan E. Fogle, Felicitas S. Boretti, Imogen Herbert, Margaret J. Hosie, and Regina Hofmann-Lehmann. 2021. "Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study" Viruses 13, no. 9: 1736. https://doi.org/10.3390/v13091736
APA StyleSpiri, A. M., Novacco, M., Meli, M. L., Stirn, M., Riond, B., Fogle, J. E., Boretti, F. S., Herbert, I., Hosie, M. J., & Hofmann-Lehmann, R. (2021). Modified-Live Feline Calicivirus Vaccination Elicits Cellular Immunity against a Current Feline Calicivirus Field Strain in an Experimental Feline Challenge Study. Viruses, 13(9), 1736. https://doi.org/10.3390/v13091736