
A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target


Abstract
:1. Introduction
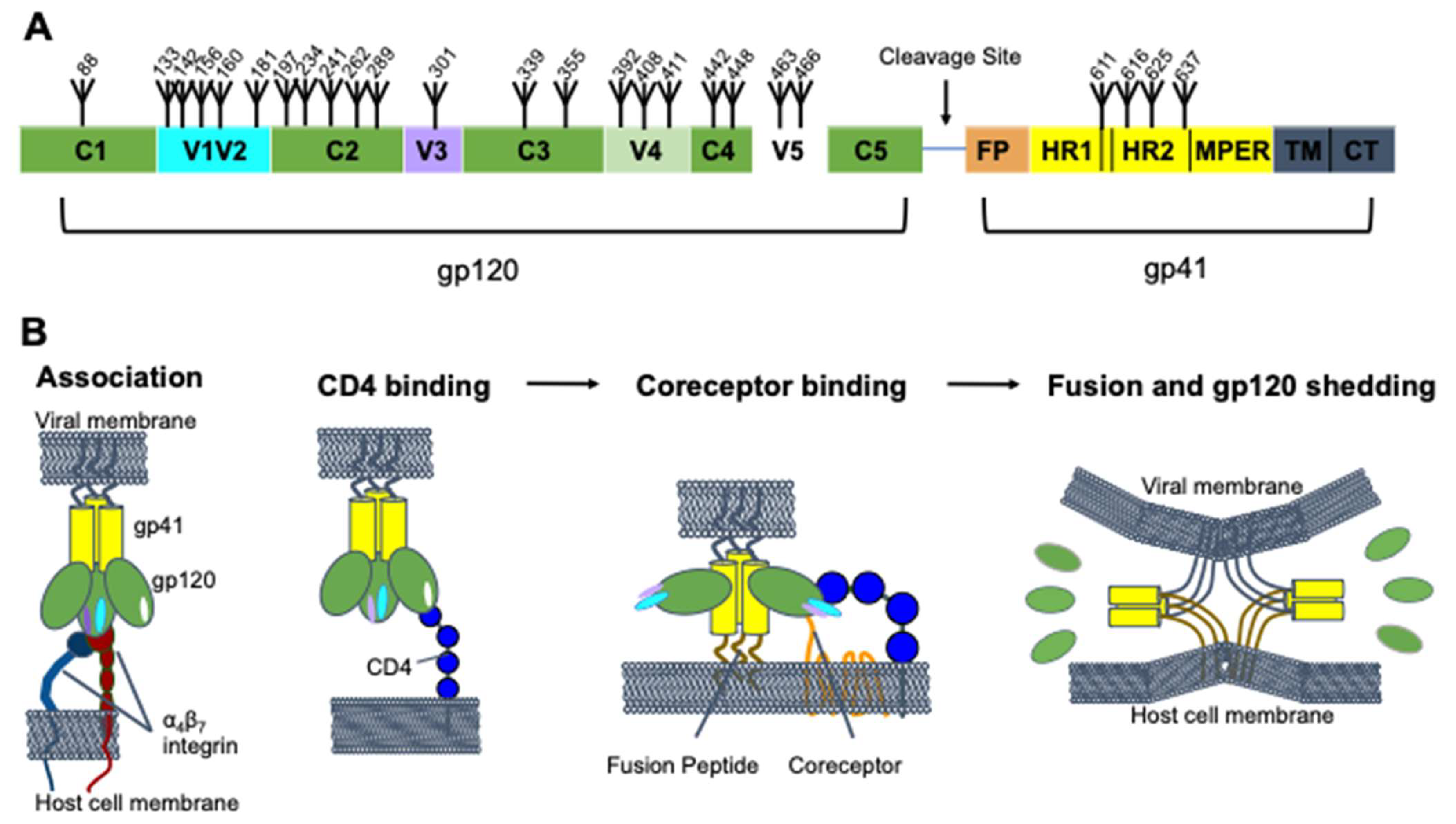
2. HIV-1 Env Structure and Function
3. HIV-1 Mechanisms for Evading Antibody Neutralization Responses
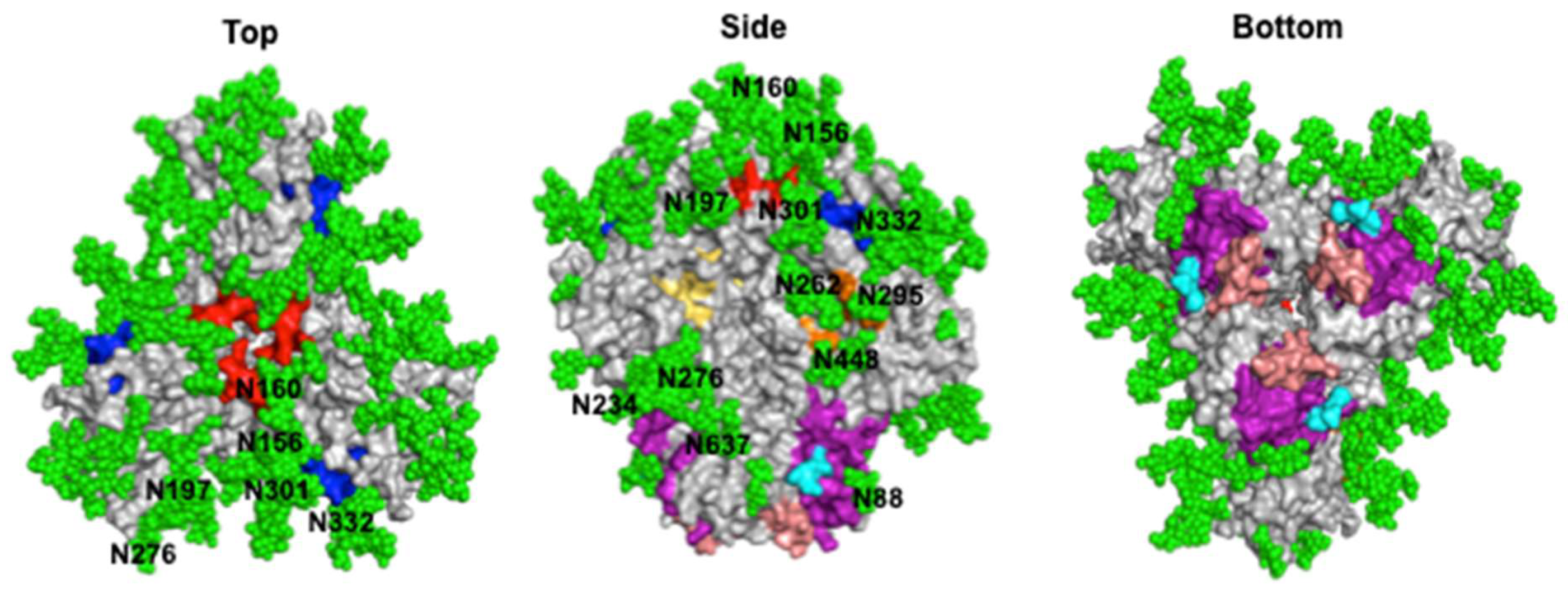
4. HIV-1 Neutralizing Epitopes
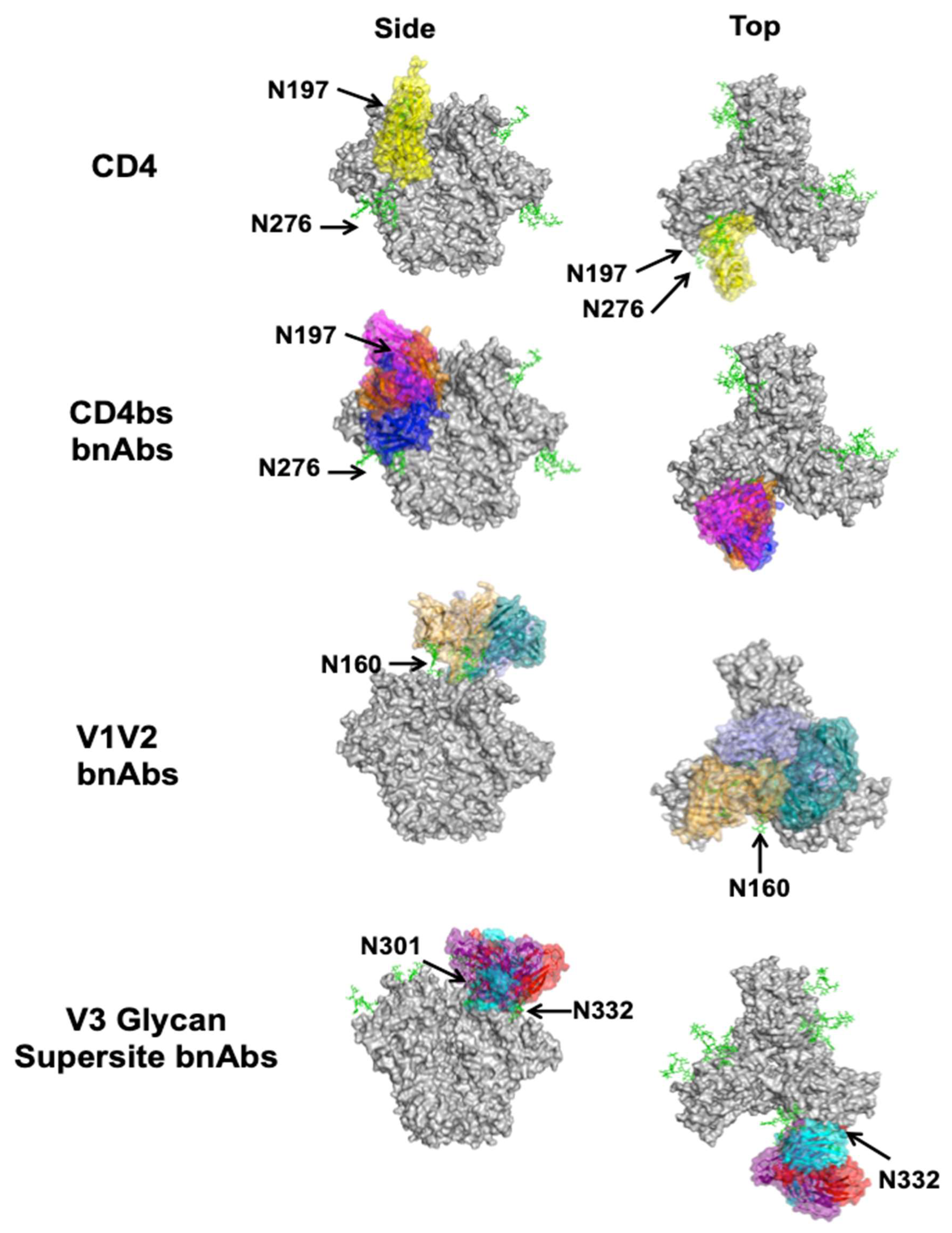
4.1. CD4 Binding Site
4.2. V1V2
4.3. V3 Glycan Supersite
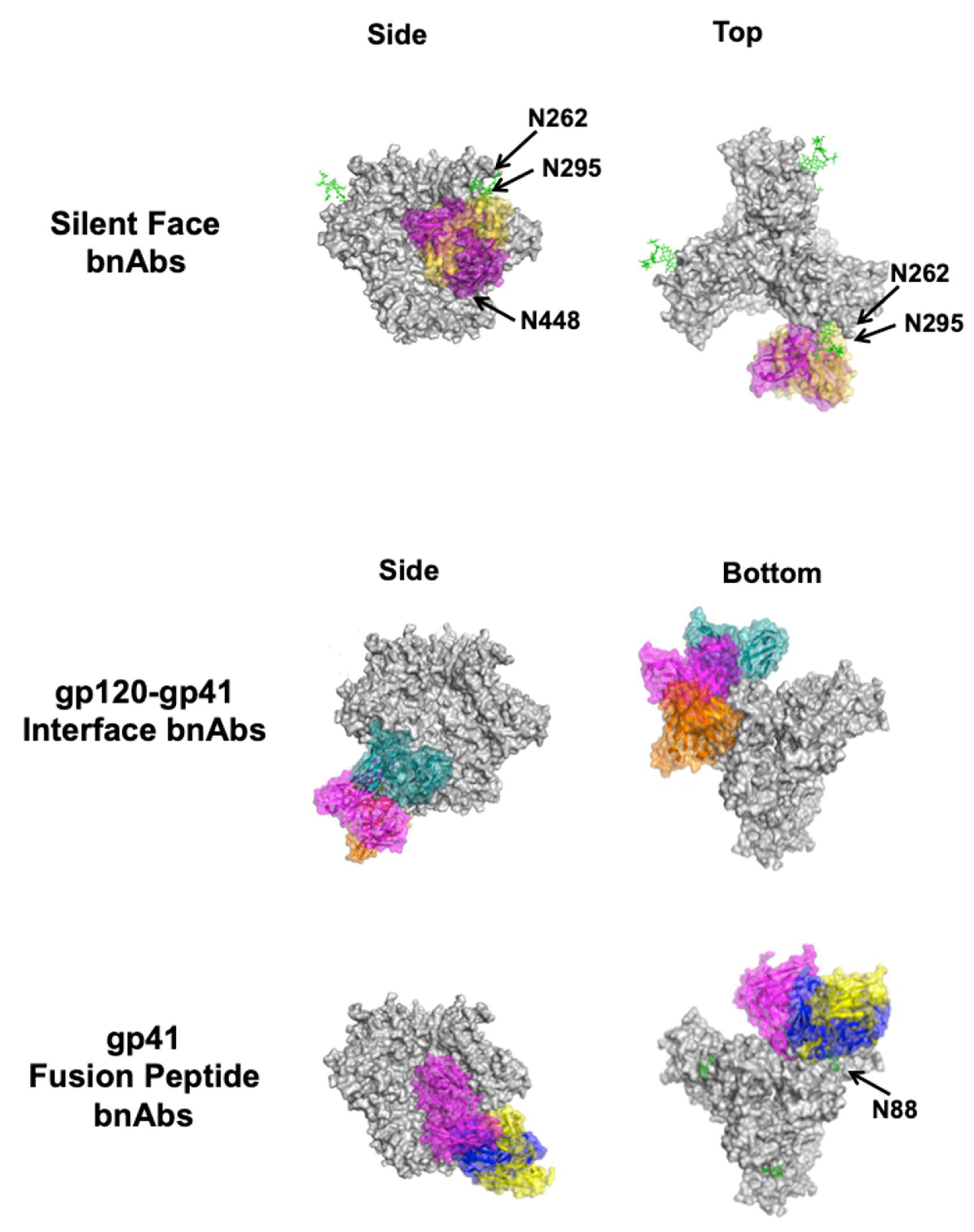
4.4. Silent Face of Gp120
4.5. Gp120–Gp41 Interface
4.6. Gp41
5. BnAb Elicitation in Mice and Other Animals
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Margolis, D.M.; Koup, R.A.; Ferrari, G. HIV Antibodies for Treatment of HIV Infection. Immunol. Rev. 2017, 275, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheid, J.F.; Horwitz, J.A.; Bar-On, Y.; Kreider, E.F.; Lu, C.-L.; Lorenzi, J.C.C.; Feldmann, A.; Braunschweig, M.; Nogueira, L.; Oliveira, T.; et al. HIV-1 Antibody 3BNC117 Suppresses Viral Rebound in Humans during Treatment Interruption. Nature 2016, 535, 556–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, R.M.; Boritz, E.; Coates, E.E.; DeZure, A.; Madden, P.; Costner, P.; Enama, M.E.; Plummer, S.; Holman, L.; Hendel, C.S.; et al. Virologic Effects of Broadly Neutralizing Antibody VRC01 Administration during Chronic HIV-1 Infection. Sci. Transl. Med. 2015, 7, 319ra206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caskey, M.; Klein, F.; Lorenzi, J.C.C.; Seaman, M.S.; West, A.P.; Buckley, N.; Kremer, G.; Nogueira, L.; Braunschweig, M.; Scheid, J.F.; et al. Viraemia Suppressed in HIV-1-Infected Humans by Broadly Neutralizing Antibody 3BNC117. Nature 2015, 522, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bar, K.J.; Sneller, M.C.; Harrison, L.J.; Justement, J.S.; Overton, E.T.; Petrone, M.E.; Salantes, D.B.; Seamon, C.A.; Scheinfeld, B.; Kwan, R.W.; et al. Effect of HIV Antibody VRC01 on Viral Rebound after Treatment Interruption. N. Engl. J. Med. 2016, 375, 2037–2050. [Google Scholar] [CrossRef] [PubMed]
- Gruell, H.; Klein, F. Antibody-Mediated Prevention and Treatment of HIV-1 Infection. Retrovirology 2018, 15, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, L.M.; Su, B.; Moog, C. Non-Neutralizing Antibodies Directed against HIV and Their Functions. Front. Immunol. 2017, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Mayr, L.M.; Decoville, T.; Schmidt, S.; Laumond, G.; Klingler, J.; Ducloy, C.; Bahram, S.; Zolla-Pazner, S.; Moog, C. Non-Neutralizing Antibodies Targeting the V1V2 Domain of HIV Exhibit Strong Antibody-Dependent Cell-Mediated Cytotoxic Activity. Sci. Rep. 2017, 7, 12655. [Google Scholar] [CrossRef] [Green Version]
- Burton, D.R.; Pyati, J.; Koduri, R.; Sharp, S.J.; Thornton, G.B.; Parren, P.W.H.I.; Sawyer, L.S.W.; Hendry, R.M.; Dunlop, N.; Nara, P.L.; et al. Efficient Neutralization of Primary Isolates of HIV-1 by a Recombinant Human Monoclonal Antibody. Science 1994, 266, 1024–1027. [Google Scholar] [CrossRef]
- Zwick, M.B.; Parren, P.W.H.I.; Saphire, E.O.; Church, S.; Wang, M.; Scott, J.K.; Dawson, P.E.; Wilson, I.A.; Burton, D.R. Molecular Features of the Broadly Neutralizing Immunoglobulin G1 B12 Required for Recognition of Human Immunodeficiency Virus Type 1 Gp120. J. Virol. 2003, 77, 5863–5876. [Google Scholar] [CrossRef] [Green Version]
- Shcherbakov, D.N.; Bakulina, A.Y.; Karpenko, L.I.; Ilyichev, A.A. Broadly Neutralizing Antibodies against HIV-1 As a Novel Aspect of the Immune Response. Acta Nat. 2015, 7, 11–21. [Google Scholar] [CrossRef]
- Kwong, P.D.; Mascola, J.R. HIV-1 Vaccines Based on Antibody Identification, B Cell Ontogeny, and Epitope Structure. Immunity 2018, 48, 855–871. [Google Scholar] [CrossRef] [Green Version]
- McCoy, L.E.; Burton, D.R. Identification and Specificity of Broadly Neutralizing Antibodies against HIV. Immunol. Rev. 2017, 275, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Burton, D.R.; Hangartner, L. Broadly Neutralizing Antibodies to HIV and Their Role in Vaccine Design. Annu. Rev. Immunol. 2016, 34, 635–659. [Google Scholar] [CrossRef] [PubMed]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.D.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C.; et al. Deep Mutational Scanning of SARS-CoV-2 Receptor Binding Domain Reveals Constraints on Folding and ACE2 Binding. Cell 2020, 182, 1295–1310.e20. [Google Scholar] [CrossRef]
- Caillat, C.; Guilligay, D.; Torralba, J.; Friedrich, N.; Nieva, J.L.; Trkola, A.; Chipot, C.J.; Dehez, F.L.; Weissenhorn, W. Structure of HIV-1 Gp41 with Its Membrane Anchors Targeted by Neutralizing Antibodies. eLife 2021, 10, e65005. [Google Scholar] [CrossRef] [PubMed]
- Leonard, C.K.; Spellman, M.W.; Riddle, L.; Harris, R.J.; Thomas, J.N.; Gregory, T.J. Assignment of Intrachain Disulfide Bonds and Characterization of Potential Glycosylation Sites of the Type 1 Recombinant Human Immunodeficiency Virus Envelope Glycoprotein (Gp120) Expressed in Chinese Hamster Ovary Cells. J. Biol. Chem. 1990, 265, 10373–10382. [Google Scholar] [CrossRef]
- Zhu, P.; Chertova, E.; Bess, J.; Lifson, J.D.; Arthur, L.O.; Liu, J.; Taylor, K.A.; Roux, K.H. Electron Tomography Analysis of Envelope Glycoprotein Trimers on HIV and Simian Immunodeficiency Virus Virions. Proc. Natl. Acad. Sci. USA 2003, 100, 15812–15817. [Google Scholar] [CrossRef] [Green Version]
- McCune, J.M.; Rabin, L.B.; Feinberg, M.B.; Lieberman, M.; Kosek, J.C.; Reyes, G.R.; Weissman, I.L. Endoproteolytic Cleavage of Gp160 Is Required for the Activation of Human Immunodeficiency Virus. Cell 1988, 53, 55–67. [Google Scholar] [CrossRef]
- Robey, W.G.; Safai, B.; Oroszlan, S.; Arthur, L.O.; Gonda, M.A.; Gallo, R.C.; Fischinger, P.J. Characterization of Envelope and Core Structural Gene Products of HTLV-III with Sera from AIDS Patients. Science 1985, 228, 593–595. [Google Scholar] [CrossRef]
- Kowalski, M.; Potz, J.; Basiripour, L.; Dorfman, T.; Goh, W.; Terwilliger, E.; Dayton, A.; Rosen, C.; Haseltine, W.; Sodroski, J. Functional Regions of the Envelope Glycoprotein of Human Immunodeficiency Virus Type 1. Science 1987, 237, 1351–1355. [Google Scholar] [CrossRef]
- Kwong, P.D.; Wyatt, R.; Robinson, J.; Sweet, R.W.; Sodroski, J.; Hendrickson, W.A. Structure of an HIV Gp120 Envelope Glycoprotein in Complex with the CD4 Receptor and a Neutralizing Human Antibody. Nature 1998, 393, 648–659. [Google Scholar] [CrossRef] [Green Version]
- Da, L.-T.; Quan, J.-M.; Wu, Y.-D. Understanding of the Bridging Sheet Formation of HIV-1 Glycoprotein Gp120. J. Phys. Chem. B 2009, 113, 14536–14543. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Vogan, E.M.; Gong, H.; Skehel, J.J.; Wiley, D.C.; Harrison, S.C. Structure of an Unliganded Simian Immunodeficiency Virus Gp120 Core. Nature 2005, 433, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Bartesaghi, A.; Merk, A.; Borgnia, M.J.; Milne, J.L.S.; Subramaniam, S. Prefusion Structure of Trimeric HIV-1 Envelope Glycoprotein Determined by Cryo-Electron Microscopy. Nat. Struct. Mol. Biol. 2013, 20, 1352–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamm, L.K.; Lee, J.; Liang, B. Capturing Glimpses of an Elusive HIV Gp41 Prehairpin Fusion Intermediate. Structure 2014, 22, 1225–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cicala, C.; Arthos, J.; Fauci, A.S. HIV-1 Envelope, Integrins and Co-Receptor Use in Mucosal Transmission of HIV. J. Transl. Med. 2011, 9, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.; Peachman, K.K.; Kim, J.; Gao, G.; Alving, C.R.; Michael, N.L.; Rao, V.B. HIV-1 Variable Loop 2 and Its Importance in HIV-1 Infection and Vaccine Development. Curr. HIV Res. 2013, 11, 427–438. [Google Scholar] [CrossRef] [Green Version]
- Kwong, P.D.; Doyle, M.L.; Casper, D.J.; Cicala, C.; Leavitt, S.A.; Majeed, S.; Steenbeke, T.D.; Venturi, M.; Chaiken, I.; Fung, M.; et al. HIV-1 Evades Antibody-Mediated Neutralization through Conformational Masking of Receptor-Binding Sites. Nature 2002, 420, 678–682. [Google Scholar] [CrossRef]
- Moore, P.L.; Crooks, E.T.; Porter, L.; Zhu, P.; Cayanan, C.S.; Grise, H.; Corcoran, P.; Zwick, M.B.; Franti, M.; Morris, L.; et al. Nature of Nonfunctional Envelope Proteins on the Surface of Human Immunodeficiency Virus Type 1. J. Virol. 2006, 80, 2515–2528. [Google Scholar] [CrossRef] [Green Version]
- Richman, D.D.; Wrin, T.; Little, S.J.; Petropoulos, C.J. Rapid Evolution of the Neutralizing Antibody Response to HIV Type 1 Infection. Proc. Natl. Acad. Sci. USA 2003, 100, 4144–4149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, D.L.; Sharp, P.M.; McCutchan, F.E.; Hahn, B.H. Recombination in HIV-1. Nature 1995, 374, 124–126. [Google Scholar] [CrossRef] [PubMed]
- Gaschen, B.; Taylor, J.; Yusim, K.; Foley, B.; Gao, F.; Lang, D.; Novitsky, V.; Haynes, B.; Hahn, B.H.; Bhattacharya, T.; et al. Diversity Considerations in HIV-1 Vaccine Selection. Science 2002, 296, 2354–2360. [Google Scholar] [CrossRef] [PubMed]
- McBurney, S.P.; Ross, T.M. Viral Sequence Diversity: Challenges for AIDS Vaccine Designs. Expert Rev. Vaccines 2008, 7, 1405–1417. [Google Scholar] [CrossRef]
- Cao, L.; Diedrich, J.K.; Kulp, D.W.; Pauthner, M.; He, L.; Park, S.-K.R.; Sok, D.; Su, C.Y.; Delahunty, C.M.; Menis, S.; et al. Global Site-Specific N-Glycosylation Analysis of HIV Envelope Glycoprotein. Nat. Commun. 2017, 8, 14954. [Google Scholar] [CrossRef]
- Zhou, T.; Zheng, A.; Baxa, U.; Chuang, G.-Y.; Georgiev, I.S.; Kong, R.; O’Dell, S.; Shahzad-Ul-Hussan, S.; Shen, C.-H.; Tsybovsky, Y.; et al. A Neutralizing Antibody Recognizing Primarily N-Linked Glycan Targets the Silent Face of the HIV Envelope. Immunity 2018, 48, 500–513.e6. [Google Scholar] [CrossRef]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody Neutralization and Escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar] [CrossRef]
- Sanders, R.W.; Vesanen, M.; Schuelke, N.; Master, A.; Schiffner, L.; Kalyanaraman, R.; Paluch, M.; Berkhout, B.; Maddon, P.J.; Olson, W.C.; et al. Stabilization of the Soluble, Cleaved, Trimeric Form of the Envelope Glycoprotein Complex of Human Immunodeficiency Virus Type 1. J. Virol. 2002, 76, 8875–8889. [Google Scholar] [CrossRef] [Green Version]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.-P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Peña, A.T.; Korzun, J.; et al. A Next-Generation Cleaved, Soluble HIV-1 Env Trimer, BG505 SOSIP.664 Gp140, Expresses Multiple Epitopes for Broadly Neutralizing but Not Non-Neutralizing Antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Bale, S.; Behrens, A.-J.; Kumar, S.; Sharma, S.K.; de Val, N.; Pallesen, J.; Irimia, A.; Diwanji, D.C.; Stanfield, R.L.; et al. Structure of a Cleavage-Independent HIV Env Recapitulates the Glycoprotein Architecture of the Native Cleaved Trimer. Nat. Commun. 2018, 9, 1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schommers, P.; Gruell, H.; Abernathy, M.E.; Tran, M.-K.; Dingens, A.S.; Gristick, H.B.; Barnes, C.O.; Schoofs, T.; Schlotz, M.; Vanshylla, K.; et al. Restriction of HIV-1 Escape by a Highly Broad and Potent Neutralizing Antibody. Cell 2020, 180, 471–489.e22. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Liberatore, R.A.; Guo, Y.; Chan, K.-W.; Pan, R.; Lu, H.; Waltari, E.; Mittler, E.; Chandran, K.; Finzi, A.; et al. VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA. Cell Host Microbe 2020, 27, 963–975.e5. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhou, T.; Schmidt, S.D.; Duan, H.; Cheng, C.; Chuang, G.-Y.; Gu, Y.; Louder, M.K.; Lin, B.C.; Shen, C.-H.; et al. Vaccination Induces Maturation in a Mouse Model of Diverse Unmutated VRC01-Class Precursors to HIV-Neutralizing Antibodies with >50% Breadth. Immunity 2021, 54, 324–339.e8. [Google Scholar] [CrossRef]
- Gorman, J.; Chuang, G.-Y.; Lai, Y.-T.; Shen, C.-H.; Boyington, J.C.; Druz, A.; Geng, H.; Louder, M.K.; McKee, K.; Rawi, R.; et al. Structure of Super-Potent Antibody CAP256-VRC26.25 in Complex with HIV-1 Envelope Reveals a Combined Mode of Trimer-Apex Recognition. Cell Rep. 2020, 31, 107488. [Google Scholar] [CrossRef] [PubMed]
- Roark, R.S.; Li, H.; Williams, W.B.; Chug, H.; Mason, R.D.; Gorman, J.; Wang, S.; Lee, F.-H.; Rando, J.; Bonsignori, M.; et al. Recapitulation of HIV-1 Env-Antibody Coevolution in Macaques Leading to Neutralization Breadth. Science 2021, 371, eabd2638. [Google Scholar] [CrossRef]
- Saunders, K.O.; Wiehe, K.; Tian, M.; Acharya, P.; Bradley, T.; Alam, S.M.; Go, E.P.; Scearce, R.; Sutherland, L.; Henderson, R.; et al. Targeted Selection of HIV-Specific Antibody Mutations by Engineering B Cell Maturation. Science 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ju, B.; Shapero, B.; Lin, X.; Ren, L.; Zhang, L.; Li, D.; Zhou, Z.; Feng, Y.; Sou, C.; et al. A V(H)1-69 Antibody Lineage from an Infected Chinese Donor Potently Neutralizes HIV-1 by Targeting the V3 Glycan Supersite. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Barnes, C.O.; Gautam, R.; Cetrulo Lorenzi, J.C.; Mayer, C.T.; Oliveira, T.Y.; Ramos, V.; Cipolla, M.; Gordon, K.M.; Gristick, H.B.; et al. A Broadly Neutralizing Macaque Monoclonal Antibody against the HIV-1 V3-Glycan Patch. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, T.; Barnes, C.O.; Suh-Toma, N.; Golijanin, J.; Schommers, P.; Gruell, H.; West, A.P.J.; Bach, F.; Lee, Y.E.; Nogueira, L.; et al. Broad and Potent Neutralizing Antibodies Recognize the Silent Face of the HIV Envelope. Immunity 2019, 50, 1513–1529.e9. [Google Scholar] [CrossRef] [Green Version]
- Dubrovskaya, V.; Tran, K.; Ozorowski, G.; Guenaga, J.; Wilson, R.; Bale, S.; Cottrell, C.A.; Turner, H.L.; Seabright, G.; O’Dell, S.; et al. Vaccination with Glycan-Modified HIV NFL Envelope Trimer-Liposomes Elicits Broadly Neutralizing Antibodies to Multiple Sites of Vulnerability. Immunity 2019, 51, 915–929.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, T.; Xu, L.; Dey, B.; Hessell, A.J.; Van Ryk, D.; Xiang, S.-H.; Yang, X.; Zhang, M.-Y.; Zwick, M.B.; Arthos, J.; et al. Structural Definition of a Conserved Neutralization Epitope on HIV-1 Gp120. Nature 2007, 445, 732–737. [Google Scholar] [CrossRef] [Green Version]
- Stewart-Jones, G.B.E.; Soto, C.; Lemmin, T.; Chuang, G.-Y.; Druz, A.; Kong, R.; Thomas, P.V.; Wagh, K.; Zhou, T.; Behrens, A.-J.; et al. Trimeric HIV-1-Env Structures Define Glycan Shields from Clades A, B and G. Cell 2016, 165, 813–826. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Lynch, R.M.; Chen, L.; Acharya, P.; Wu, X.; Doria-Rose, N.A.; Joyce, M.G.; Lingwood, D.; Soto, C.; Bailer, R.T.; et al. Structural Repertoire of HIV-1-Neutralizing Antibodies Targeting the CD4 Supersite in 14 Donors. Cell 2015, 161, 1280–1292. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Zhu, J.; Wu, X.; Moquin, S.; Zhang, B.; Acharya, P.; Georgiev, I.S.; Altae-Tran, H.R.; Chuang, G.-Y.; Joyce, M.G.; et al. Multi-Donor Analysis Reveals Structural Elements, Genetic Determinants, and Maturation Pathway for Effective HIV-1 Neutralization by VRCO1-Class Antibodies. Immunity 2013, 39, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Pan, R.; Gorny, M.K.; Zolla-Pazner, S.; Kong, X.-P. The V1V2 Region of HIV-1 Gp120 Forms a Five-Stranded Beta Barrel. J. Virol. 2015, 89, 8003–8010. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.-X.; Lynch, R.; Zhou, T.; Gao, F.; Alam, S.M.; Boyd, S.D.; Fire, A.Z.; Roskin, K.M.; Schramm, C.A.; Zhang, Z.; et al. Co-Evolution of a Broadly Neutralizing HIV-1 Antibody and Founder Virus. Nature 2013, 496, 469–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, J.; Soto, C.; Yang, M.M.; Davenport, T.M.; Guttman, M.; Bailer, R.T.; Chambers, M.; Chuang, G.-Y.; DeKosky, B.J.; Doria-Rose, N.A.; et al. Structures of HIV-1 Env V1V2 with Broadly Neutralizing Antibodies Reveal Commonalities That Enable Vaccine Design. Nat. Struct. Mol. Biol. 2016, 23, 81–90. [Google Scholar] [CrossRef] [PubMed]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.-P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 Gp120 V1/V2 Domain with Broadly Neutralizing Antibody PG9. Nature 2011, 480, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Löving, R.; Sjöberg, M.; Wu, S.-R.; Binley, J.M.; Garoff, H. Inhibition of the HIV-1 Spike by Single-PG9/16-Antibody Binding Suggests a Coordinated-Activation Model for Its Three Protomeric Units. J. Virol. 2013, 87, 7000–7007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Andrabi, R.; Su, C.-Y.; Yasmeen, A.; Julien, J.-P.; Kong, L.; Wu, N.C.; McBride, R.; Sok, D.; Pauthner, M.; et al. A Broadly Neutralizing Antibody Targets the Dynamic HIV Envelope Trimer Apex via a Long, Rigidified, and Anionic β-Hairpin Structure. Immunity 2017, 46, 690–702. [Google Scholar] [CrossRef] [Green Version]
- Cale, E.M.; Gorman, J.; Radakovich, N.A.; Crooks, E.T.; Osawa, K.; Tong, T.; Li, J.; Nagarajan, R.; Ozorowski, G.; Ambrozak, D.R.; et al. Virus-like Particles Identify an HIV V1V2 Apex-Binding Neutralizing Antibody That Lacks a Protruding Loop. Immunity 2017, 46, 777–791.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonsignori, M.; Kreider, E.F.; Fera, D.; Meyerhoff, R.R.; Bradley, T.; Wiehe, K.; Alam, S.M.; Aussedat, B.; Walkowicz, W.E.; Hwang, K.-K.; et al. Staged Induction of HIV-1 Glycan–Dependent Broadly Neutralizing Antibodies. Sci. Transl. Med. 2017, 9, eaai7514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonich, C.A.; Williams, K.L.; Verkerke, H.P.; Williams, J.A.; Nduati, R.; Lee, K.K.; Overbaugh, J. HIV-1 Neutralizing Antibodies with Limited Hypermutation from an Infant. Cell 2016, 166, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Sok, D.; Pauthner, M.; Briney, B.; Lee, J.H.; Saye-Francisco, K.L.; Hsueh, J.; Ramos, A.; Le, K.M.; Jones, M.; Jardine, J.G.; et al. A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity 2016, 45, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.-P.; Wang, S.-K.; Ramos, A.; Chan-Hui, P.-Y.; Moyle, M.; et al. Broad Neutralization Coverage of HIV by Multiple Highly Potent Antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Shaik, M.M.; Peng, H.; Lu, J.; Rits-Volloch, S.; Xu, C.; Liao, M.; Chen, B. Structural Basis of Coreceptor Recognition by HIV-1 Envelope Spike. Nature 2019, 565, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Wilen, C.B.; Tilton, J.C.; Doms, R.W. HIV: Cell Binding and Entry. Cold Spring Harb. Perspect. Med. 2012, 2, a006866. [Google Scholar] [CrossRef] [PubMed]
- Daniels, C.N.; Saunders, K.O. Antibody Responses to the HIV-1 Envelope High Mannose Patch. Adv. Immunol. 2019, 143, 11–73. [Google Scholar] [CrossRef]
- Moyo, T.; Kitchin, D.; Moore, P.L. Targeting the N332-Supersite of the HIV-1 Envelope for Vaccine Design. Expert Opin. Ther. Targets 2020, 24, 499–509. [Google Scholar] [CrossRef]
- Sok, D.; Burton, D.R. Recent Progress in Broadly Neutralizing Antibodies to HIV. Nat. Immunol. 2018, 19, 1179–1188. [Google Scholar] [CrossRef]
- Fera, D.; Lee, M.S.; Wiehe, K.; Meyerhoff, R.R.; Piai, A.; Bonsignori, M.; Aussedat, B.; Walkowicz, W.E.; Ton, T.; Zhou, J.O.; et al. HIV Envelope V3 Region Mimic Embodies Key Features of a Broadly Neutralizing Antibody Lineage Epitope. Nat. Commun. 2018, 9, 1111. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.P.; Willey, R.L.; Lewis, G.K.; Robinson, J.; Sodroski, J. Immunological Evidence for Interactions between the First, Second, and Fifth Conserved Domains of the Gp120 Surface Glycoprotein of Human Immunodeficiency Virus Type 1. J. Virol. 1994, 68, 6836–6847. [Google Scholar] [CrossRef] [Green Version]
- Seabright, G.E.; Cottrell, C.A.; van Gils, M.J.; D’addabbo, A.; Harvey, D.J.; Behrens, A.-J.; Allen, J.D.; Watanabe, Y.; Scaringi, N.; Polveroni, T.M.; et al. Networks of HIV-1 Envelope Glycans Maintain Antibody Epitopes in the Face of Glycan Additions and Deletions. Structure 2020, 28, 897–909.e6. [Google Scholar] [CrossRef]
- Van Gils, M.J.; van den Kerkhof, T.L.G.M.; Ozorowski, G.; Cottrell, C.A.; Sok, D.; Pauthner, M.; Pallesen, J.; de Val, N.; Yasmeen, A.; de Taeye, S.W.; et al. An HIV-1 Antibody from an Elite Neutralizer Implicates the Fusion Peptide as a Site of Vulnerability. Nat. Microbiol. 2016, 2, 16199. [Google Scholar] [CrossRef]
- Blattner, C.; Lee, J.H.; Sliepen, K.; Derking, R.; Falkowska, E.; de la Peña, A.T.; Cupo, A.; Julien, J.-P.; van Gils, M.; Lee, P.S.; et al. Structural Delineation of a Quaternary, Cleavage-Dependent Epitope at the Gp41–Gp120 Interface on Intact HIV-1 Env Trimers. Immunity 2014, 40, 669–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.H.; Leaman, D.P.; Kim, A.S.; Torrents de la Peña, A.; Sliepen, K.; Yasmeen, A.; Derking, R.; Ramos, A.; de Taeye, S.W.; Ozorowski, G.; et al. Antibodies to a Conformational Epitope on Gp41 Neutralize HIV-1 by Destabilizing the Env Spike. Nat. Commun. 2015, 6, 8167. [Google Scholar] [CrossRef] [PubMed]
- Scharf, L.; Scheid, J.F.; Lee, J.H.; West, A.P.; Chen, C.; Gao, H.; Gnanapragasam, P.N.P.; Mares, R.; Seaman, M.S.; Ward, A.B.; et al. Antibody 8ANC195 Reveals a Site of Broad Vulnerability on the HIV-1 Envelope Spike. Cell Rep. 2014, 7, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Pancera, M.; Zhou, T.; Druz, A.; Georgiev, I.S.; Soto, C.; Gorman, J.; Huang, J.; Acharya, P.; Chuang, G.-Y.; Ofek, G.; et al. Structure and Immune Recognition of Trimeric Prefusion HIV-1 Env. Nature 2014, 514, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Chuang, G.-Y.; Zhou, J.; Acharya, P.; Rawi, R.; Shen, C.-H.; Sheng, Z.; Zhang, B.; Zhou, T.; Bailer, R.T.; Dandey, V.P.; et al. Structural Survey of Broadly Neutralizing Antibodies Targeting the HIV-1 Env Trimer Delineates Epitope Categories and Characteristics of Recognition. Structure 2019, 27, 196–206. [Google Scholar] [CrossRef] [Green Version]
- Eckert, D.M.; Kim, P.S. Mechanisms of Viral Membrane Fusion and Its Inhibition. Annu. Rev. Biochem. 2001, 70, 777–810. [Google Scholar] [CrossRef] [Green Version]
- Kong, R.; Xu, K.; Zhou, T.; Acharya, P.; Lemmin, T.; Liu, K.; Ozorowski, G.; Soto, C.; Taft, J.D.; Bailer, R.T.; et al. Fusion Peptide of HIV-1 as a Site of Vulnerability to Neutralizing Antibody. Science 2016, 352, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.C. Viral Membrane Fusion. Nat. Struct. Mol. Biol. 2008, 15, 690. [Google Scholar] [CrossRef]
- Colman, P.M.; Lawrence, M.C. The Structural Biology of Type I Viral Membrane Fusion. Nat. Rev. Mol. Cell Biol. 2003, 4, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Ananthaswamy, N.; Fang, Q.; AlSalmi, W.; Jain, S.; Chen, Z.; Klose, T.; Sun, Y.; Liu, Y.; Mahalingam, M.; Chand, S.; et al. A Sequestered Fusion Peptide in the Structure of an HIV-1 Transmitted Founder Envelope Trimer. Nat. Commun. 2019, 10, 873. [Google Scholar] [CrossRef] [Green Version]
- Kong, R.; Louder, M.K.; Wagh, K.; Bailer, R.T.; de Camp, A.; Greene, K.; Gao, H.; Taft, J.D.; Gazumyan, A.; Liu, C.; et al. Improving Neutralization Potency and Breadth by Combining Broadly Reactive HIV-1 Antibodies Targeting Major Neutralization Epitopes. J. Virol. 2015, 89, 2659–2671. [Google Scholar] [CrossRef] [Green Version]
- De la Peña, A.T.; Rantalainen, K.; Cottrell, C.A.; Allen, J.D.; van Gils, M.J.; Torres, J.L.; Crispin, M.; Sanders, R.W.; Ward, A.B. Similarities and Differences between Native HIV-1 Envelope Glycoprotein Trimers and Stabilized Soluble Trimer Mimetics. PLoS Pathog. 2019, 15, e1007920. [Google Scholar] [CrossRef] [Green Version]
- Irimia, A.; Sarkar, A.; Stanfield, R.L.; Wilson, I.A. Crystallographic Identification of Lipid as an Integral Component of the Epitope of HIV Broadly Neutralizing Antibody 4E10. Immunity 2016, 44, 21–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, L.D.; Ofek, G.; Schätzle, S.; McDaniel, J.R.; Lu, X.; Nicely, N.I.; Wu, L.; Lougheed, C.S.; Bradley, T.; Louder, M.K.; et al. Potent and Broad HIV-Neutralizing Antibodies in Memory B Cells and Plasma. Sci. Immunol. 2017, 2, eaal2200. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, R.M.F.; Zwick, M.B.; Stanfield, R.L.; Kunert, R.; Binley, J.M.; Katinger, H.; Burton, D.R.; Wilson, I.A. Broadly Neutralizing Anti-HIV Antibody 4E10 Recognizes a Helical Conformation of a Highly Conserved Fusion-Associated Motif in Gp41. Immunity 2005, 22, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Ofek, G.; Laub, L.; Louder, M.K.; Doria-Rose, N.A.; Longo, N.S.; Imamichi, H.; Bailer, R.T.; Chakrabarti, B.; Sharma, S.K.; et al. Broad and Potent Neutralization of HIV-1 by a Gp41-Specific Human Antibody. Nature 2012, 491, 406–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ofek, G.; Tang, M.; Sambor, A.; Katinger, H.; Mascola, J.R.; Wyatt, R.; Kwong, P.D. Structure and Mechanistic Analysis of the Anti-Human Immunodeficiency Virus Type 1 Antibody 2F5 in Complex with Its Gp41 Epitope. J. Virol. 2004, 78, 10724–10737. [Google Scholar] [CrossRef] [Green Version]
- Krebs, S.J.; Kwon, Y.D.; Schramm, C.A.; Law, W.H.; Donofrio, G.; Zhou, K.H.; Gift, S.; Dussupt, V.; Georgiev, I.S.; Schätzle, S.; et al. Longitudinal Analysis Reveals Early Development of Three MPER-Directed Neutralizing Antibody Lineages from an HIV-1-Infected Individual. Immunity 2019, 50, 677–691.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, D.; Fenwick, C.; Caillat, C.; Silacci, C.; Guseva, S.; Dehez, F.; Chipot, C.; Barbieri, S.; Minola, A.; Jarrossay, D.; et al. Structural Basis for Broad HIV-1 Neutralization by the MPER-Specific Human Broadly Neutralizing Antibody LN01. Cell Host Microbe 2019, 26, 623–637.e8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Irimia, A.; He, L.; Landais, E.; Rantalainen, K.; Leaman, D.P.; Vollbrecht, T.; Stano, A.; Sands, D.I.; Kim, A.S.; et al. An MPER Antibody Neutralizes HIV-1 Using Germline Features Shared among Donors. Nat. Commun. 2019, 10, 5389. [Google Scholar] [CrossRef]
- Pejchal, R.; Gach, J.S.; Brunel, F.M.; Cardoso, R.M.; Stanfield, R.L.; Dawson, P.E.; Burton, D.R.; Zwick, M.B.; Wilson, I.A. A Conformational Switch in Human Immunodeficiency Virus Gp41 Revealed by the Structures of Overlapping Epitopes Recognized by Neutralizing Antibodies. J. Virol. 2009, 83, 8451–8462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morris, L.; Chen, X.; Alam, M.; Tomaras, G.; Zhang, R.; Marshall, D.J.; Chen, B.; Parks, R.; Foulger, A.; Jaeger, F.; et al. Isolation of a Human Anti-HIV Gp41 Membrane Proximal Region Neutralizing Antibody by Antigen-Specific Single B Cell Sorting. PLoS ONE 2011, 6, e23532. [Google Scholar] [CrossRef]
- Caillat, C.; Guilligay, D.; Sulbaran, G.; Weissenhorn, W. Neutralizing Antibodies Targeting HIV-1 Gp41. Viruses 2020, 12, 1210. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.L.; Andrabi, R.; Cottrell, C.A.; Richey, S.T.; Song, G.; Callaghan, S.; Anzanello, F.; Moyer, T.J.; Abraham, W.; Melo, M.; et al. Disassembly of HIV Envelope Glycoprotein Trimer Immunogens Is Driven by Antibodies Elicited via Immunization. Sci. Adv. 2021, 7, eabh2791. [Google Scholar] [CrossRef]
- Xu, K.; Acharya, P.; Kong, R.; Cheng, C.; Chuang, G.-Y.; Liu, K.; Louder, M.K.; O’Dell, S.; Rawi, R.; Sastry, M.; et al. Epitope-Based Vaccine Design Yields Fusion Peptide-Directed Antibodies That Neutralize Diverse Strains of HIV-1. Nat. Med. 2018, 24, 857–867. [Google Scholar] [CrossRef]
- Dingens, A.S.; Acharya, P.; Haddox, H.K.; Rawi, R.; Xu, K.; Chuang, G.-Y.; Wei, H.; Zhang, B.; Mascola, J.R.; Carragher, B.; et al. Complete Functional Mapping of Infection- and Vaccine-Elicited Antibodies against the Fusion Peptide of HIV. PLoS Pathog. 2018, 14, e1007159. [Google Scholar] [CrossRef] [Green Version]
- Doria-Rose, N.A.; Schramm, C.A.; Gorman, J.; Moore, P.L.; Bhiman, J.N.; DeKosky, B.J.; Ernandes, M.J.; Georgiev, I.S.; Kim, H.J.; Pancera, M.; et al. Developmental Pathway for Potent V1V2-Directed HIV-Neutralizing Antibodies. Nature 2014, 509, 55–62. [Google Scholar] [CrossRef]
- Mendoza, P.; Gruell, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.Y.; Lorenzi, J.C.C.; et al. Combination Therapy with Anti-HIV-1 Antibodies Maintains Viral Suppression. Nature 2018, 561, 479–484. [Google Scholar] [CrossRef]
- Bar-On, Y.; Gruell, H.; Schoofs, T.; Pai, J.A.; Nogueira, L.; Butler, A.L.; Millard, K.; Lehmann, C.; Suárez, I.; Oliveira, T.Y.; et al. Safety and Antiviral Activity of Combination HIV-1 Broadly Neutralizing Antibodies in Viremic Individuals. Nat. Med. 2018, 24, 1701–1707. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Antanasijevic, A.; de Gast, M.; Allen, J.D.; Bijl, T.P.L.; Yasmeen, A.; Ravichandran, R.; Burger, J.A.; Ozorowski, G.; Torres, J.L.; et al. Immunofocusing and Enhancing Autologous Tier-2 HIV-1 Neutralization by Displaying Env Trimers on Two-Component Protein Nanoparticles. NPJ Vaccines 2021, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Karch, C.P.; Bai, H.; Torres, O.B.; Tucker, C.A.; Michael, N.L.; Matyas, G.R.; Rolland, M.; Burkhard, P.; Beck, Z. Design and Characterization of a Self-Assembling Protein Nanoparticle Displaying HIV-1 Env V1V2 Loop in a Native-like Trimeric Conformation as Vaccine Antigen. Nanomed. Nanotechnol. Biol. Med. 2019, 16, 206–216. [Google Scholar] [CrossRef]
- Sok, D.; Briney, B.; Jardine, J.G.; Kulp, D.W.; Menis, S.; Pauthner, M.; Wood, A.; Lee, E.-C.; Le, K.M.; Jones, M.; et al. Priming HIV-1 Broadly Neutralizing Antibody Precursors in Human Ig Loci Transgenic Mice. Science 2016, 353, 1557–1560. [Google Scholar] [CrossRef] [Green Version]
- He, L.; de Val, N.; Morris, C.D.; Vora, N.; Thinnes, T.C.; Kong, L.; Azadnia, P.; Sok, D.; Zhou, B.; Burton, D.R.; et al. Presenting Native-like Trimeric HIV-1 Antigens with Self-Assembling Nanoparticles. Nat. Commun. 2016, 7, 12041. [Google Scholar] [CrossRef]
- Jardine, J.G.; Ota, T.; Sok, D.; Pauthner, M.; Kulp, D.W.; Kalyuzhniy, O.; Skog, P.D.; Thinnes, T.C.; Bhullar, D.; Briney, B.; et al. Priming a Broadly Neutralizing Antibody Response to HIV-1 Using a Germline-Targeting Immunogen. Science 2015, 349, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Jardine, J.; Julien, J.-P.; Menis, S.; Ota, T.; Kalyuzhniy, O.; McGuire, A.; Sok, D.; Huang, P.-S.; MacPherson, S.; Jones, M.; et al. Rational HIV Immunogen Design to Target Specific Germline B Cell Receptors. Science 2013, 340, 711–716. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epitope | bnAb | PDB ID | Neutralization Potency (IC50) | Breadth (%) | Year Published | Reference |
|---|---|---|---|---|---|---|
| CD4bs | PGV19 | 6B0N | 0.34–1.3 µg/mL | 70–75 | 2018 | [40] |
| 1–18 | 6UDJ | 0.048 µg/mL | 97 | 2020 | [41] | |
| 1–55 | 6UDK | 0.096 µg/mL | 92 | 2020 | [41] | |
| M1214_N1 | 6VY2 | 0.19 µg/mL | 65 | 2020 | [42] | |
| 2411a | 7JKS | 1.49 µg/mL | 51 | 2021 | [43] | |
| 2413a | 7JKT | 3.86 µg/mL | 34 | 2021 | [43] | |
| V1V2 | CAP256-VRC26.25 | 6VTT | 0.012 µg/mL | 59 | 2020 | [44] |
| RHA1.V2.01 | 6XRT | 0.35 µg/mL | 49 | 2021 | [45] | |
| V3 glycan supersite | DH270.6 | 6UM6 | 0.08 µg/mL | 55 | 2019 | [46] |
| 438-B11 | 6UTK | 0.18 µg/mL | 67 | 2020 | [47] | |
| Ab1485 | 7KDE | 0.055 µg/mL | 38 | 2020 | [48] | |
| Silent face | SF12 | 6OKP | 0.20 µg/mL | 62 | 2019 | [49] |
| gp120–gp41 interface | 1C2 | 6P65 | 8.03 µg/mL | 85 | 2019 | [50] |
| gp41 | N/A | N/A | N/A | N/A | N/A | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parker Miller, E.; Finkelstein, M.T.; Erdman, M.C.; Seth, P.C.; Fera, D. A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target. Viruses 2021, 13, 1774. https://doi.org/10.3390/v13091774
Parker Miller E, Finkelstein MT, Erdman MC, Seth PC, Fera D. A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target. Viruses. 2021; 13(9):1774. https://doi.org/10.3390/v13091774
Chicago/Turabian StyleParker Miller, Emma, Maxwell T. Finkelstein, Molly C. Erdman, Paul C. Seth, and Daniela Fera. 2021. "A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target" Viruses 13, no. 9: 1774. https://doi.org/10.3390/v13091774
APA StyleParker Miller, E., Finkelstein, M. T., Erdman, M. C., Seth, P. C., & Fera, D. (2021). A Structural Update of Neutralizing Epitopes on the HIV Envelope, a Moving Target. Viruses, 13(9), 1774. https://doi.org/10.3390/v13091774