
Nonstructural Protein 2 Is Critical to Infection Efficiency of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus on PAMs and Influence Virulence In Vivo


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses, Plasmids and Antibodies
2.2. Construction of Full-Length Chimeric cDNA Clones
2.3. In Vitro Transcription and Virus Recovery
2.4. Virus Infection Efficiency Test on PAMs
2.5. Animal and Experimental Design
2.6. Viral Copy Detection
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Rescue of Chimeric and Mutant Viruses
3.2. NSP2 Primarily Contributes to the Infection Efficiency in PAMs
3.3. Determination of the Key Amino Acids in NSP2 Gene
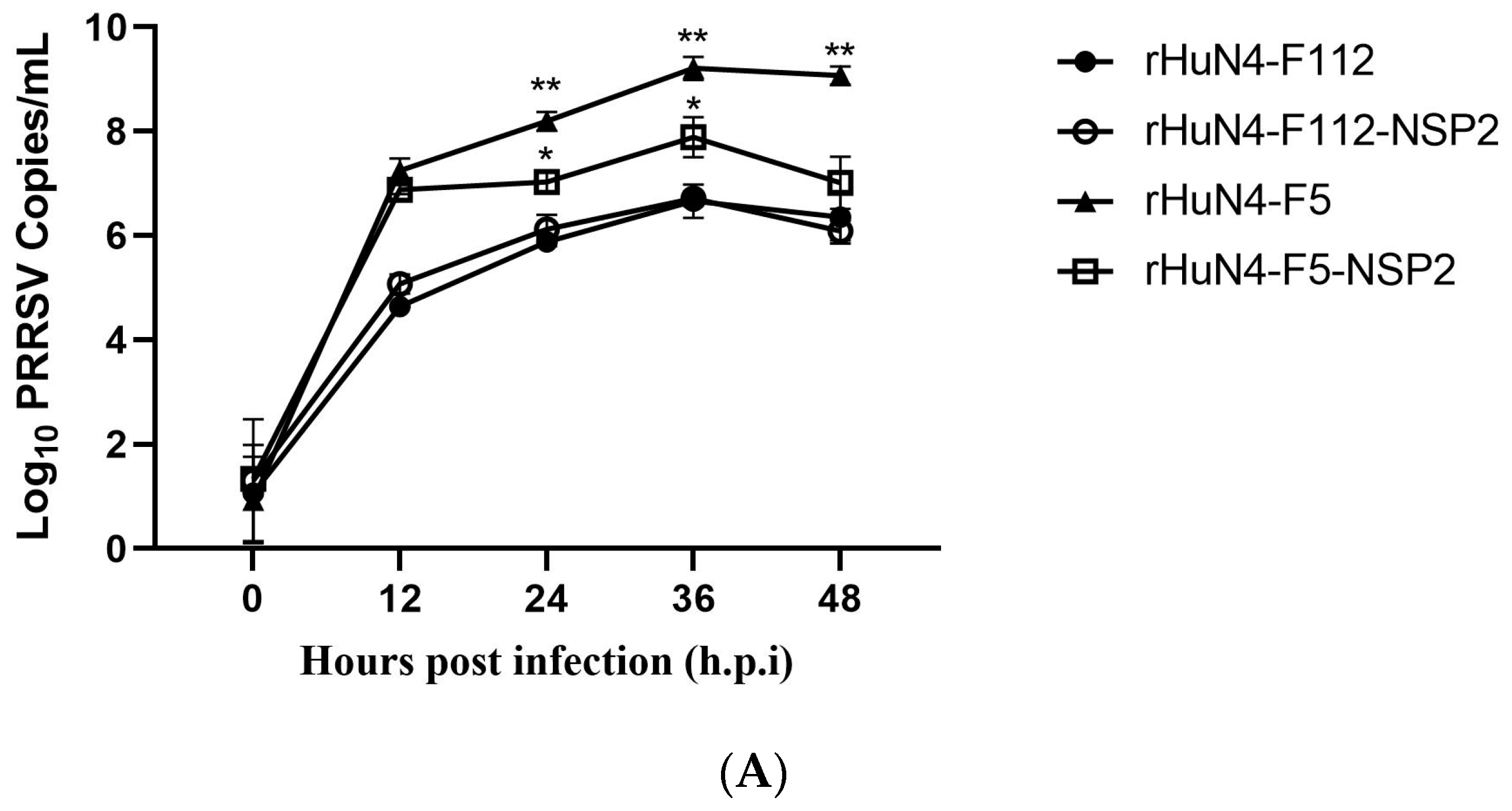
3.4. NSP2 Influence Virus Replication In Vitro
3.5. Clinical Performance after Infection
3.6. Tissue Necropsy Changes after Infection
3.7. Viremia Copies in Sera
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Wensvoort, G.; de Kluyver, E.P.; Pol, J.M.A.; Wagenaar, F.; Moormann, R.J.M.; Hulst, M.M.; Bloemraad, R.; den Besten, A.; Zetstra, T.; Terpstra, C. Lelystad virus, the cause of porcine epidemic abortion and respiratory syndrome: A review of mystery swine disease research at Lelystad. Vet. Microbiol. 1992, 33, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Wensvoort, G.; Terpstra, C.; Pol, J.M.; ter Laak, E.A.; Bloemraad, M.; de Kluyver, E.P.; Kragten, C.; van Buiten, L.; den Besten, A.; Wagenaar, F.; et al. Mystery swine disease in The Netherlands: The isolation of Lelystad virus. Vet. Q. 1991, 13, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.E.; Benfield, D.A.; Christianson, W.T.; Harris, L.; Hennings, J.C.; Shaw, D.P.; Goyal, S.M.; McCullough, S.; Morrison, R.B.; Joo, H.S.; et al. Isolation of swine infertility and respiratory syndrome virus (isolate ATCC VR-2332) in North America and experimental reproduction of the disease in gnotobiotic pigs. J. Vet. Diagn. Investig. 1992, 4, 117–126. [Google Scholar] [CrossRef]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef]
- Snijder, E.J.; Meulenberg, J.J. The molecular biology of arteriviruses. J. Gen. Virol. 1998, 79 Pt 5, 961–979. [Google Scholar] [CrossRef]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2016). Arch. Virol. 2016, 161, 2921–2949. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bào, Y.; Ng, T.F.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Firth, A.E.; Zevenhoven-Dobbe, J.C.; Wills, N.M.; Go, Y.Y.; Balasuriya, U.B.R.; Atkins, J.F.; Snijder, E.J.; Posthuma, C.C. Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production. J. Gen. Virol. 2011, 92, 1097–1106. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, H. Porcine reproductive and respiratory syndrome in China. Virus Res. 2010, 154, 31–37. [Google Scholar] [CrossRef]
- Bautista, E.M.; Meulenberg, J.J.; Choi, C.S.; Molitor, T.W. Structural polypeptides of the American (VR-2332) strain of porcine reproductive and respiratory syndrome virus. Arch. Virol. 1996, 141, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Snijder, E.J. The PRRSV replicase: Exploring the multifunctionality of an intriguing set of nonstructural proteins. Virus Res. 2010, 154, 61–76. [Google Scholar] [CrossRef] [PubMed]
- van Aken, D.; Zevenhoven-Dobbe, J.; Gorbalenya, A.E.; Snijder, E.J. Proteolytic maturation of replicase polyprotein pp1a by the nsp4 main proteinase is essential for equine arteritis virus replication and includes internal cleavage of nsp7. J. Gen. Virol. 2006, 87, 3473–3482. [Google Scholar] [CrossRef] [PubMed]
- Ziebuhr, J.; Snijder, E.J.; Gorbalenya, A.E. Virus-encoded proteinases and proteolytic processing in the Nidovirales. J. Gen. Virol. 2000, 81, 853–879. [Google Scholar] [CrossRef]
- Kroese, M.V.; Zevenhoven-Dobbe, J.C.; Bos-de Ruijter, J.N.A.; Peeters, B.P.H.; Meulenberg, J.J.M.; Cornelissen, L.A.H.M.; Snijder, E.J. The nsp1alpha and nsp1 papain-like autoproteinases are essential for porcine reproductive and respiratory syndrome virus RNA synthesis. J. Gen. Virol. 2008, 89, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.-H.; Fang, Y.; Rowland, R.R.R.; Lawson, S.R.; Christopher-Hennings, J.; Yoon, K.-J.; Nelson, E.A. The 2b protein as a minor structural component of PRRSV. Virus Res. 2005, 114, 177–181. [Google Scholar] [CrossRef]
- Shang, P.; Yuan, F.; Misra, S.; Li, Y.; Fang, Y. Hyper-phosphorylation of nsp2-related proteins of porcine reproductive and respiratory syndrome virus. Virology 2020, 543, 63–75. [Google Scholar] [CrossRef]
- Kim, H.S.; Kwang, J.; Yoon, I.J.; Joo, H.S.; Frey, M.L. Enhanced replication of porcine reproductive and respiratory syndrome (PRRS) virus in a homogeneous subpopulation of MA-104 cell line. Arch. Virol. 1993, 133, 477–483. [Google Scholar] [CrossRef]
- Duan, X.; Nauwynck, H.J.; Pensaert, M.B. Effects of origin and state of differentiation and activation of monocytes/macrophages on their susceptibility to porcine reproductive and respiratory syndrome virus (PRRSV). Arch. Virol. 1997, 142, 2483–2497. [Google Scholar] [CrossRef]
- Duan, X.; Nauwynck, H.J.; Pensaert, M.B. Virus quantification and identification of cellular targets in the lungs and lymphoid tissues of pigs at different time intervals after inoculation with porcine reproductive and respiratory syndrome virus (PRRSV). Vet. Microbiol. 1997, 56, 9–19. [Google Scholar] [CrossRef]
- Jiang, Y.; Tong, W.; Yu, L.; Li, L.; Gao, F.; Li, G.; Liu, C.; Chen, P.; Shen, Q.; Zhang, Y.; et al. Identification of Virulence Associated Region during Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus during Attenuation In Vitro: Complex Question with Different Strain Backgrounds. Viruses 2022, 14, 40. [Google Scholar] [CrossRef] [PubMed]
- Mittelholzer, C.; Moser, C.; Tratschin, J.-D.; Hofmann, M.A. Analysis of classical swine fever virus replication kinetics allows differentiation of highly virulent from avirulent strains. Vet. Microbiol. 2000, 74, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, J.; Zeng, J.; Yin, S.; Li, Y.; Zheng, L.; Guo, X.; Ge, X.; Yang, H. The 30-Amino-Acid Deletion in the Nsp2 of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Emerging in China Is Not Related to Its Virulence. J. Virol. 2009, 83, 5156–5167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allende, R.; Kutish, G.F.; Laegreid, W.; Lu, Z.; Lewis, T.L.; Rock, D.L.; Friesen, J.; Galeota, J.A.; Doster, A.R.; Osorio, F.A. Mutations in the genome of porcine reproductive and respiratory syndrome virus responsible for the attenuation phenotype. Arch. Virol. 2000, 145, 1149–1161. [Google Scholar] [CrossRef]
- Kwon, B.; Ansari, I.H.; Pattnaik, A.K.; Osorio, F.A. Identification of virulence determinants of porcine reproductive and respiratory syndrome virus through construction of chimeric clones. Virology 2008, 380, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faaberg, K.S.; Kehrli, M.E., Jr.; Lager, K.M.; Guo, B.; Han, J. In vivo growth of porcine reproductive and respiratory syndrome virus engineered nsp2 deletion mutants. Virus Res. 2010, 154, 77–85. [Google Scholar] [CrossRef]
- Morgan, S.B.; Graham, S.P.; Salguero, F.J.; Cordon, P.J.S.; Mokhtar, H.; Rebel, J.M.J.; Weesendorp, E.; Bodman-Smith, K.B.; Steinbach, F.; Frossard, J.P. Increased pathogenicity of European porcine reproductive and respiratory syndrome virus is associated with enhanced adaptive responses and viral clearance. Vet. Microbiol. 2013, 163, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-Y.; Calvert, J.G.; Chang, K.-O.; Horlen, K.; Kerrigan, M.; Rowland, R.R.R. Expression and stability of foreign tags inserted into nsp2 of porcine reproductive and respiratory syndrome virus (PRRSV). Virus Res. 2007, 128, 106–114. [Google Scholar] [CrossRef]
- Kim, D.-Y.; Kaiser, T.J.; Horlen, K.; Keith, M.L.; Taylor, L.P.; Jolie, R.; Calvert, J.G.; Rowland, R.R.R. Insertion and deletion in a non-essential region of the nonstructural protein 2 (nsp2) of porcine reproductive and respiratory syndrome (PRRS) virus: Effects on virulence and immunogenicity. Virus Genes 2009, 38, 118–128. [Google Scholar] [CrossRef]
- Wang, F.-X.; Qin, L.-T.; Liu, Y.; Liu, X.; Sun, N.; Yang, Y.; Chen, T.; Zhu, H.-W.; Ren, J.-Q.; Sun, Y.-J.; et al. Novel Nsp2 deletion based on molecular epidemiology and evolution of porcine reproductive and respiratory syndrome virus in Shandong Province from 2013 to 2014. Infect. Genet. Evol. 2015, 33, 219–226. [Google Scholar] [CrossRef]
- Xu, L.; Zhou, L.; Sun, W.; Zhang, P.; Ge, X.; Guo, X.; Han, J.; Yang, H. Nonstructural protein 9 residues 586 and 592 are critical sites in determining the replication efficiency and fatal virulence of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus. Virology 2018, 517, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Gao, J.-C.; Xiong, J.-Y.; Guo, J.-C.; Yang, Y.-B.; Jiang, C.-G.; Tang, Y.-D.; Tian, Z.-J.; Cai, X.-H.; Tong, G.-Z.; et al. Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus. J. Virol. 2018, 92, e02209-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liang, Y.; Han, J.; Burkhart, K.M.; Vaughn, E.M.; Roof, M.B.; Faaberg, K.S. Attenuation of porcine reproductive and respiratory syndrome virus strain MN184 using chimeric construction with vaccine sequence. Virology 2008, 371, 418–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Bai, J.; Wang, H.; Fan, B.; Li, Y.; Jiang, P. Effect of amino acids residues 323–433 and 628–747 in Nsp2 of representative porcine reproductive and respiratory syndrome virus strains on inflammatory response in vitro. Virus Res. 2015, 208, 13–21. [Google Scholar] [CrossRef]
- Wang, F.-X.; Wen, Y.-J.; Yang, B.-C.; Liu, Z.; Shi, X.C.; Leng, X.; Song, N.; Wu, H.; Chen, L.-Z.; Cheng, S.-P. Role of non-structural protein 2 in the regulation of the replication of the porcine reproductive and respiratory syndrome virus in MARC-145 cells: Effect of gene silencing and over expression. Vet. Microbiol. 2012, 161, 58–65. [Google Scholar] [CrossRef]
- Han, J.; Liu, G.; Wang, Y.; Faaberg, K.S. Identification of nonessential regions of the nsp2 replicase protein of porcine reproductive and respiratory syndrome virus strain VR-2332 for replication in cell culture. J. Virol. 2007, 81, 9878–9890. [Google Scholar] [CrossRef] [Green Version]
- Kappes, M.A.; Miller, C.L.; Faaberg, K.S. Highly divergent strains of porcine reproductive and respiratory syndrome virus incorporate multiple isoforms of nonstructural protein 2 into virions. J. Virol. 2013, 87, 13456–13465. [Google Scholar] [CrossRef] [Green Version]
- Kappes, M.A.; Miller, C.L.; Faaberg, K.S. Porcine reproductive and respiratory syndrome virus nonstructural protein 2 (nsp2) topology and selective isoform integration in artificial membranes. Virology 2015, 481, 51–62. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′-3′) | Information | Chimeric Viruses |
|---|---|---|---|
| Q-RT401F Q-RT401L | CTATGGTCGCTCGTCACGCTTC TGGACGAAGCGTGACGAGCGAC | Primers used for mutated NSP2 401aa from rHuN4-F5 to rHuN4-F112 | rHuN4-F5-401/477 |
| Q-RT477F Q-RT477L | GATCTTGTGAACATCATCCAAA CTGAGGATTTGGATGATGTTCA | Primers used for mutated NSP2 477aa from rHuN4-F5 to rHuN4-F112 | |
| Q-RT799F Q-RT799L | GGCTGAGCAGGTCAATTTGAAAGCT ACCCAAGCTTTCAAATTGACCTGCT | Primers used for mutated NSP2 799aa from rHuN4-F5to rHuN4-F112 | rHuN4-F5-799 |
| Q-RT893F Q-RT893L | TCGCGACGTGCCCCCAAGCTGATGA AAGGTGTCATCAGCTTGGGGGCACG | Primers used for mutated NSP2 893aa from rHuN4-F5 to rHuN4-F112 | rHuN4-F5-893 and rHuN4-F5-893/979 |
| Q-RT979F Q-RT979L | CTGTGTCATCAAGCAGCCCCTTGTC CTTAATACTTGACAAGGGGCTGCTT | Primers used for mutated NSP2 979aa from rHuN4-F5 to rHuN4-F112 | rHuN4-F5-979 and rHuN4-F5-893/979 |
| Q-RT1060F Q-RT1060L | CGTTTCGCATCTTAAGTGGCAGGTT AAACTCAAACCTGCCACTTAAGATG | Primers used for mutated NSP2 1060aa from rHuN4-F5 to rHuN4-F112 | rHuN4-F5-1060/1136 |
| Q-RT1136F Q-RT1136L | CTCTAAGGGAGAACCGGTCAGTGAC GGCAGGTTGGTCACTGACCGGTTCT | Primers used for mutated NSP2 1136aa from rHuN4-F5 to rHuN4-F112 | |
| R-QN401U R-QN401L | CTATGGTCGCTCATCACGCTTC AGCGGACGAAGCGTGATGAGCG | Primers used for mutated NSP2 401aa from rHuN4-F112 to rHuN4-F5 | rHuN4-F112-401/477 |
| R-QN477F R-QN477L | GATCTTGTGAACACCATCCAAA GAGGATTTGGATGGTGTTCACA | Primers used for mutated NSP2 477aa from rHuN4-F112 to rHuN4-F5 | |
| R-QN799F R-QN799L | TGGGCGGCTGAGCAGGTCGATTTAA CCAAGCTTTTAAATCGACCTGCTCA | Primers used for mutated NSP2 799aa from rHuN4-F112 to rHuN4-F5 | rHuN4-F112-799 |
| R-QN893F R-QN893L | GTCGCGACGTGTCCCCAAGCTGATG GTCATCAGCTTGGGGACACGTCGCG | Primers used for mutated NSP2 893aa from rHuN4-F112 to rHuN4-F5 | rHuN4-F112-893 and rHuN4-F112-893/979 |
| R-QN979F R-QN979L | GTGTCATCAAGCAGCCCCTTGTCAA ATCTTAATACTTGACAAGGGGCTGC | Primers used for mutated NSP2 979aa from rHuN4-F112 to rHuN4-F5 | rHuN4-F112-979 and rHuN4-F112-893/979 |
| R-QN1060F R-QN1060L | GTTTCGCATCTTAAATGGCAGGTTT AACTCAAACCTGCCATTTAAGATGC | Primers used for mutated NSP2 1060aa from rHuN4-F112 to rHuN4-F5 | rHuN4-F112-1060/1136 |
| R-QN1136F R-QN1136L | CAAGGGAGAACCGGTCTGTGACCAA TTGGCAGGTTGGTCACAGACCGGTT | Primers used for mutated NSP2 1136aa from rHuN4-F112 to rHuN4-F5 |
| Designation | Number | Macroscopic (Lung) | |||
|---|---|---|---|---|---|
| Mean ± SD | Pathological Changes (Score) | ||||
| ≤30 | 30 to 50 | ≥50 | |||
| rHuN4-F5 | 5 | 89 ± 9.64 d | 0 | 0 | 5 |
| rHuN4-F5-NSP2 | 5 | 77.4 ± 31.94 d | 1 | 0 | 4 |
| rHuN4-F5-893/979 | 5 | 90.5 ± 3.55 d | 0 | 0 | 5 |
| rHuN4-F112 | 5 | 23.2 ± 15.42 a | 5 | 0 | 0 |
| rHuN4-F112-NSP2 | 5 | 26.8 ± 25.47 b | 2 | 2 | 1 |
| rHuN4-F112-893/979 | 5 | 53.8 ± 21.87 b,c | 1 | 2 | 2 |
| Control | 5 | 17.6 ± 17.67 a | 4 | 1 | 0 |
| Designation | Macroscopic (Organs) | |||||
|---|---|---|---|---|---|---|
| Kidney | Spleen | Tonsil | Liver | Mesenteric Lymph Nodes | Inguinal Lymph Nodes | |
| rHuN4-F5 | 3 ± 0 c | 1.4 ± 1.14 | 1 ± 1 | 2 ± 0.7 b | 2.2 ± 1.1 | 0.4 ± 0.55 |
| rHuN4-F112 | 0 ± 0 a,c | 0.6 ± 1.34 | 0 ± 0 | 0 ± 0 a | 0 ± 0 | 0.8 ± 1.09 |
| rHuN4-F5-NSP2 | 1.2 ± 1.1 a,c | 0.2 ± 0.44 | 1.4 ± 1.34 | 1.4 ± 0.89 a,b | 0.6 ± 0.89 | 1.8 ± 1.3 |
| rHuN4-F112-NSP2 | 0.4 ± 0.89 a,b | 1 ± 1.4 | 0 ± 0 | 0.8 ± 1.09 a,b | 1 ± 1.44 | 1.2 ± 1.64 |
| rHuN4-F5-893/979 | 1.4 ± 1.14 a,c | 0.4 ± 0.55 | 1.6 ± 1.14 | 1.2 ± 0.83 a,b | 1.4 ± 1.34 | 1 ± 0.7 |
| rHuN4-F112-893/979 | 0.2 ± 0.45 a | 0 ± 0 | 0.4 ± 0.89 | 0 ± 0 a | 0.2 ± 0.45 | 0.4 ± 0.55 |
| Control | 0.4 ± 0.55 a | 0 ± 0 | 0 ± 0 | 0 ± 0 a | 0 ± 0 | 0.8 ± 1.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Yu, L.; Zhou, Y.; Yang, S.; Bai, Y.; Wang, Q.; Peng, J.; An, T.; Gao, F.; Li, L.; et al. Nonstructural Protein 2 Is Critical to Infection Efficiency of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus on PAMs and Influence Virulence In Vivo. Viruses 2022, 14, 2613. https://doi.org/10.3390/v14122613
Chen J, Yu L, Zhou Y, Yang S, Bai Y, Wang Q, Peng J, An T, Gao F, Li L, et al. Nonstructural Protein 2 Is Critical to Infection Efficiency of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus on PAMs and Influence Virulence In Vivo. Viruses. 2022; 14(12):2613. https://doi.org/10.3390/v14122613
Chicago/Turabian StyleChen, Jiazeng, Lingxue Yu, Yanjun Zhou, Shen Yang, Yun Bai, Qian Wang, Jinmei Peng, Tongqing An, Fei Gao, Liwei Li, and et al. 2022. "Nonstructural Protein 2 Is Critical to Infection Efficiency of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus on PAMs and Influence Virulence In Vivo" Viruses 14, no. 12: 2613. https://doi.org/10.3390/v14122613
APA StyleChen, J., Yu, L., Zhou, Y., Yang, S., Bai, Y., Wang, Q., Peng, J., An, T., Gao, F., Li, L., Ye, C., Liu, C., Tong, G., Cai, X., Tian, Z., & Jiang, Y. (2022). Nonstructural Protein 2 Is Critical to Infection Efficiency of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus on PAMs and Influence Virulence In Vivo. Viruses, 14(12), 2613. https://doi.org/10.3390/v14122613