
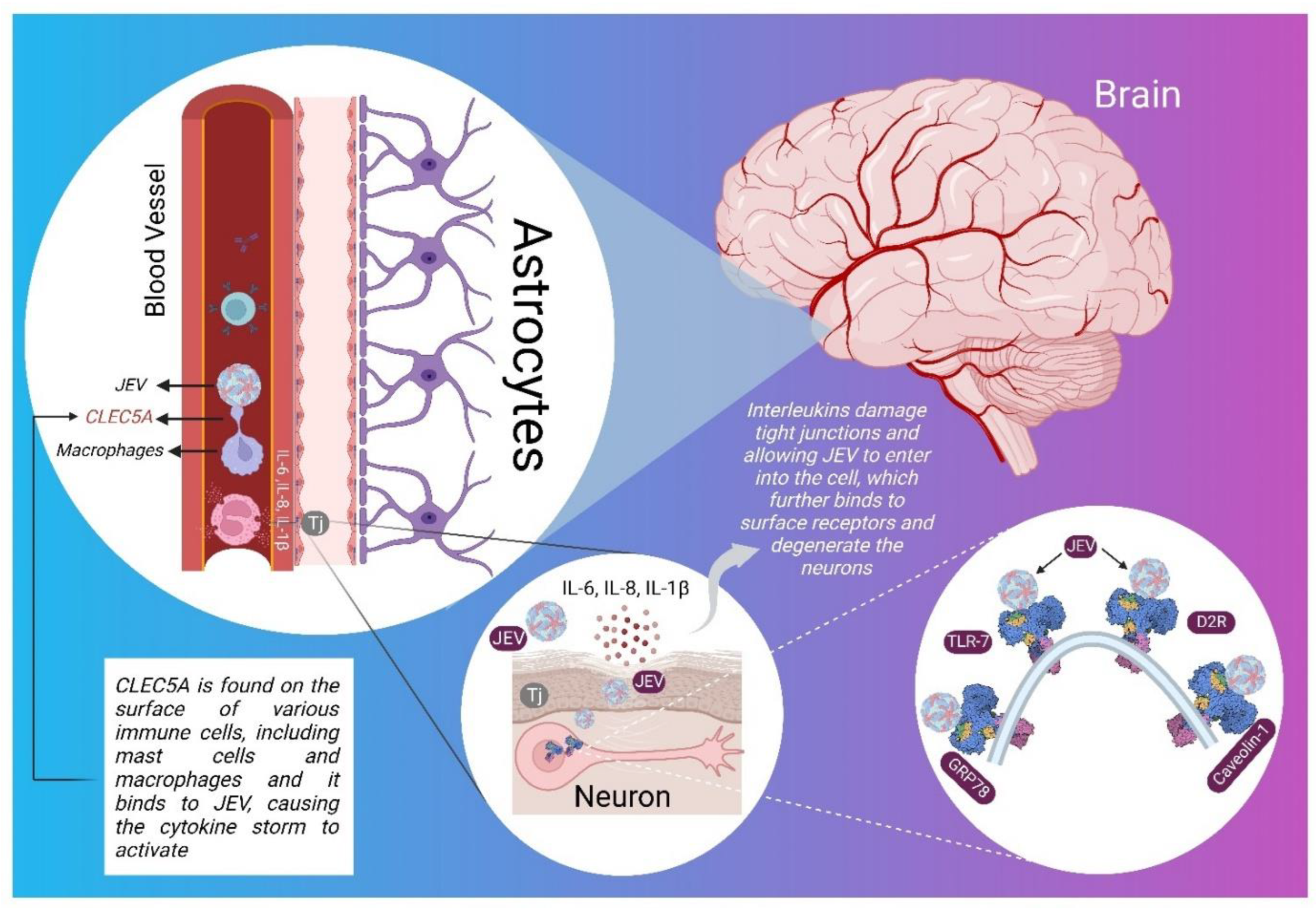
Molecular Mechanism and Role of Japanese Encephalitis Virus Infection in Central Nervous System-Mediated Diseases

 ,
,  , , and
, , and 
{kind=link}
{kind=link}
Abstract
Share and Cite
Yadav, P.; Chakraborty, P.; Jha, N.K.; Dewanjee, S.; Jha, A.K.; Panda, S.P.; Mishra, P.C.; Dey, A.; Jha, S.K. Molecular Mechanism and Role of Japanese Encephalitis Virus Infection in Central Nervous System-Mediated Diseases. Viruses 2022, 14, 2686. https://doi.org/10.3390/v14122686
Yadav P, Chakraborty P, Jha NK, Dewanjee S, Jha AK, Panda SP, Mishra PC, Dey A, Jha SK. Molecular Mechanism and Role of Japanese Encephalitis Virus Infection in Central Nervous System-Mediated Diseases. Viruses. 2022; 14(12):2686. https://doi.org/10.3390/v14122686
Chicago/Turabian StyleYadav, Pardeep, Pratik Chakraborty, Niraj Kumar Jha, Saikat Dewanjee, Abhimanyu Kumar Jha, Siva Prasad Panda, Prabhu Chandra Mishra, Abhijit Dey, and Saurabh Kumar Jha. 2022. "Molecular Mechanism and Role of Japanese Encephalitis Virus Infection in Central Nervous System-Mediated Diseases" Viruses 14, no. 12: 2686. https://doi.org/10.3390/v14122686
APA StyleYadav, P., Chakraborty, P., Jha, N. K., Dewanjee, S., Jha, A. K., Panda, S. P., Mishra, P. C., Dey, A., & Jha, S. K. (2022). Molecular Mechanism and Role of Japanese Encephalitis Virus Infection in Central Nervous System-Mediated Diseases. Viruses, 14(12), 2686. https://doi.org/10.3390/v14122686