
Tropism of Highly Pathogenic Avian Influenza H5 Viruses from the 2020/2021 Epizootic in Wild Ducks and Geese

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Virus Histochemistry
2.3. Virology, Pathology and Immunohistochemistry of Naturally Infected Wild Birds
3. Results
3.1. Pattern of Virus Attachment to Epithelia of Digestive and Respiratory Tracts
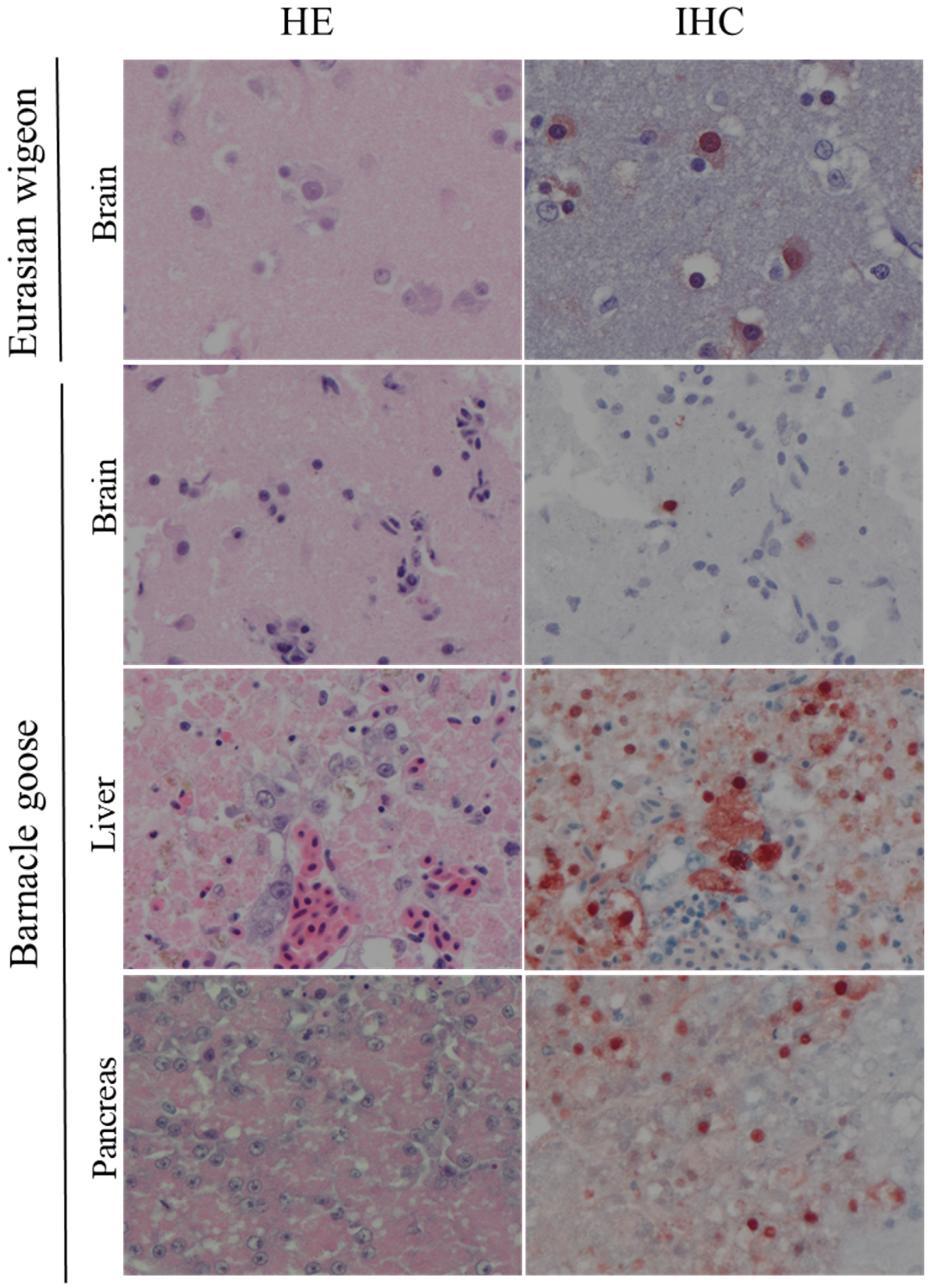
3.2. Virology, Influenza Virus Antigen Expression and Associated Lesions in Naturally Infected Wild Birds
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lycett, S.J.; Pohlmann, A.; Staubach, C.; Caliendo, V.; Woolhouse, M.; Beer, M.; Kuiken, T. Global Consortium for HN, Related Influenza V. Genesis and spread of multiple reassortants during the 2016/2017 H5 avian influenza epidemic in Eurasia. Proc. Natl. Acad. Sci. USA 2020, 117, 20814–20825. [Google Scholar] [CrossRef] [PubMed]
- Lycett, S.; Bodewes, R.; Pohlmann, A.; Bank, J.; Banyai, K.; Boni, M. Role for migratory wild birds in the global spread of avian influenza H5N8. Science 2016, 354, 213–217. [Google Scholar]
- Madslien, K.; Moldal, T.; Gjerset, B.; Gudmundsson, S.; Follestad, A.; Whittard, E.; Tronerud, O.H.; Dean, K.R.; Åkerstedt, J.; Jørgensen, H.J.; et al. First detection of highly pathogenic avian influenza virus in Norway. BMC Vet. Res. 2021, 17, 218. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lv, X.; Li, Y.; Peng, P.; Zhou, R.; Qin, S.; Ma, E.; Liu, W.; Fu, T.; Ma, P.; et al. Highly Pathogenic Avian Influenza A(H5N8) Virus in Swans, China, 2020. Emerg. Infect. Dis. 2021, 27, 1732–1734. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, E.; Smietanka, K.; Staubach, C.; Terregino, C.; Baldinelli, F. Avian influenza overview—Update on 19 November 2020, EU/EEA and the UK. EFSA J. 2020, 18, 6341. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, E.; Staubach, C.; Terregino, C.; Baldinelli, F. Scientific Report: Avian influenza overview August–December 2020. EFSA J. 2020, 18, 6379. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Lima, E.; Muñoz Guajardo, I.; et al. Scientific report: Avian influenza overview December 2020–February 2021. EFSA J. 2021, 19, 6497. [Google Scholar] [CrossRef]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Aznar, I.; Muñoz Guajardo, I.; et al. Scientific report: Avian influenza overview February–May 2021. EFSA J. 2021, 19, e06951. [Google Scholar]
- Adlhoch, C.; Fusaro, A.; Gonzales, J.L.; Kuiken, T.; Marangon, S.; Niqueux, É.; Staubach, C.; Terregino, C.; Aznar, I.; Muñoz Guajardo, I.; et al. Scientific report: Avian influenza overview May–August 2021. EFSA J. 2021, 18, e06270. [Google Scholar]
- Verhagen, J.H.; Fouchier, R.A.M.; Lewis, N. Highly Pathogenic Avian Influenza Viruses at the Wild–Domestic Bird Interface in Europe: Future Directions for Research and Surveillance. Viruses 2021, 13, 212. [Google Scholar] [CrossRef]
- Kleyheeg, E.; Slaterus, R.; Bodewes, R.; Rijks, J.M.; Spierenburg, M.A.H.; Beerens, N.; Kelder, L.; Poen, M.J.; Stegeman, J.A.; Fouchier, R.A.M.; et al. Deaths among Wild Birds during Highly Pathogenic Avian Influenza A (H5N8) Virus Outbreak, The Netherlands. Emerg. Infect Dis. 2017, 23, 2050–2054. [Google Scholar] [CrossRef] [PubMed]
- Poen, M.J.; Bestebroer, T.M.; Vuong, O.; Scheuer, R.D.; van der Jeugd, H.P.; Kleyheeg, E.; Eggink, D.; Lexmond, P.; van den Brand, J.M.A.; Begeman, L.; et al. Local amplification of highly pathogenic avian influenza H5N8 viruses in wild birds in The Netherlands, 2016 to 2017. Eurosurveillance 2018, 23, 17-00449. [Google Scholar] [CrossRef]
- Lewis, N.S.; Banyard, A.C.; Whittard, E.; Karibayev, T.; Al Kafagi, T.; Chvala, I.; Byrne, A.; Meruyert Akberovna, S.; King, J.; Harder, T.; et al. Emergence and spread of novel H5N8, H5N5 and H5N1 clade 2.3.4.4 highly pathogenic avian influenza in 2020. Emerg. Microbes Infect. 2021, 10, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Keawcharoen, J.; Van Riel, D.; Van Amerongen, G.; Bestebroer, T.; Beyer, W.; Van Lavieren, R.; Osterhaus, A.; Fouchier, R.; Kuiken, T. Wild Ducks as Long-Distance Vectors of Highly Pathogenic Avian Influenza Virus (H5N1). Emerg. Infect. Dis. 2008, 14, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Van den Brand, J.M.A.; Verhagen, J.H.; Veldhuis Kroeze, E.J.B.; van de Bildt, M.W.G.; Bodewes, R.; Herfst, S.; Richard, M.; Lexmond, P.; Bestebroer, T.M.; Fouchier, R.A.M.; et al. Wild ducks excrete highly pathogenic avian influenza virus H5N8 (2014–2015) without clinical or pathological evidence of disease. Emerg. Microbes Infect. 2018, 7, 67. [Google Scholar] [CrossRef] [Green Version]
- Caliendo, V.; Leijten, L.; Begeman, L.; Poen, M.J.; Fouchier, R.A.M.; Beerens, N.; Kuiken, T. Enterotropism of highly pathogenic avian influenza virus H5N8 from the 2016/2017 epidemic in some wild bird species. Vet. Res. 2020, 51, 117. [Google Scholar] [CrossRef]
- Munster, V.J.; Baas, C.; Lexmond, P.; Bestebroer, T.M.; Guldemeester, J.; Beyer, W.E.; de Wit, E.; Schutten, M.; Rimmelzwaan, G.F.; Osterhaus, A.D.M.E.; et al. Practical considerations for high-throughput influenza A virus surveillance studies of wild birds by use of molecular diagnostic tests. J. Clin. Microbiol. 2009, 47, 666–673. [Google Scholar] [CrossRef] [Green Version]
- Kuiken, T. Is low pathogenic avian influenza virus virulent for wild waterbirds? Proc. Biol. Sci. 2013, 280, 1763. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Kleijn, D.; Müskens, G.J.D.M.; Fouchier, R.A.M.; Verhagen, J.H.; Glazov, P.M.; Si, Y.; Prins, H.H.T.; de Boer, W.F. No evidence that migratory geese disperse avian influenza viruses from breeding to wintering ground. PLoS ONE 2017, 12, e0177790. [Google Scholar] [CrossRef] [Green Version]
- Guillemain, M.; Martin, G.R.; Fritz, H. Feeding methods, visual fields and vigilance in dabbling ducks (Anatidae). Functional. Ecol. 2002, 16, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Dessborn, L.; Brochet, A.L.; Elmberg, J.; Legagneux, P.; Gauthier-Clerc, M.; Guillemain, M. Geographical and temporal patterns in the diet of pintail Anas acuta, wigeon Anas penelope, mallard Anas platyrhynchos and teal Anas crecca in the Western Palearctic. Eur. J. Wildl. Res. 2011, 57, 1119–1129. [Google Scholar] [CrossRef]
- Percival, S.M.; Percival, T. Feeding ecology of barnacle geese on their spring staging grounds in northern Iceland. Ecography 1997, 20, 461–465. [Google Scholar] [CrossRef]
- Black, J.M.; Carbone, C.; Wells, R.L.; Owen, M. Foraging dynamics in goose flocks: The cost of living on the edge. Anim. Behav. 1992, 44, 41–50. [Google Scholar] [CrossRef]
- Swayne, D.E. Understanding the Complex Pathobiology of High Pathogenicity Avian Influenza Viruses in Birds. Avian Dis. 2007, 51, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Bröjer, C.; Ågren, E.O.; Uhlhorn, H.; Bernodt, K.; Jansson, D.S.; Gavier-Widén, D. Characterization of Encephalitis in Wild Birds Naturally Infected by Highly Pathogenic Avian Influenza H5N1. Avian Dis. 2012, 56, 144–152. [Google Scholar] [CrossRef] [PubMed]
- van den Brand, J.M.; Krone, O.; Wolf, P.U.; van de Bildt, M.W.; van Amerongen, G.; Osterhaus, A.D.; Kuiken, T. Host-specific exposure and fatal neurologic disease in wild raptors from highly pathogenic avian influenza virus H5N1 during the 2006 outbreak in Germany. Vet. Res. 2015, 46, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grund, C.; Hoffmann, D.; Ulrich, R.; Naguib, M.; Schinköthe, J.; Hoffmann, B.; Harder, T.; Saenger, S.; Zscheppang, K.; Tönnies, M.; et al. A novel European H5N8 influenza A virus has increased virulence in ducks but low zoonotic potential. Emerg. Microbes Infect. 2018, 7, 132. [Google Scholar] [CrossRef]
- Genovart, M.; Negre, N.; Tavecchia, G.; Bistuer, A.; Parpal, L.; Oro, D. The young, the weak and the sick: Evidence of natural selection by predation. PLoS ONE 2010, 5, e9774. [Google Scholar] [CrossRef] [Green Version]
- Lebarbenchon, C.; Feare, C.J.; Renaud, F.; Thomas, F.; Gauthier-Clerc, M. Persistence of highly pathogenic avian influenza viruses in natural ecosystems. Emerg. Infect Dis. 2010, 16, 1057–1062. [Google Scholar] [CrossRef]
- Geoghegan, J.L.; Holmes, E.C. The phylogenomics of evolving virus virulence. Nat. Rev. Genet. 2018, 19, 756–769. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; George, D.; Pepin, K.M.; Pitzer, V.E.; Pulliam, J.R.C.; Dobson, A.P.; Hudson, P.J.; Grenfell, B.T. Epidemic dynamics at the human-animal interface. Science 2009, 326, 1362–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Avian Influenza Viruses | |||||
|---|---|---|---|---|---|
| Species | Tissues | 2005 H5N1 | 2014 H5N8 | 2016 H5N8 | 2020 H5N8 |
| Mallard | Duodenum | ++ | + | ++ | ++ |
| Jejunum | ++ | ++ | ++ | ++ | |
| Ileum | + | + | ++ | ++ | |
| Colon | + | + | ++ | ++ | |
| Eurasian wigeon | Duodenum | ± | ± | ± | ± |
| Jejunum | − | ± | ± | ± | |
| Ileum | − | ± | − | ± | |
| Colon | − | ± | − | ± | |
| Tufted duck | Duodenum | ± | ± | ± | ± |
| Jejunum | − | ± | ± | − | |
| Ileum | − | ± | − | − | |
| Colon | ± | ± | − | − | |
| Graylag goose | Duodenum | ± | ± | ± | + |
| Jejunum | nd | nd | nd | nd | |
| Ileum | nd | nd | nd | nd | |
| Colon | ± | ± | ± | ± | |
| Chicken | Duodenum | ++ | ++ | ++ | ++ |
| Jejunum | ++ | ++ | ++ | ++ | |
| Ileum | ++ | ++ | ++ | ++ | |
| Colon | + | + | + | + | |
| Avian Influenza Viruses | |||||
|---|---|---|---|---|---|
| Species | Tissues | 2005 H5N1 | 2014 H5N8 | 2016 H5N8 | 2020 H5N8 |
| Mallard | Trachea | ++ | ++ | ++ | ++ |
| Primary bronchus | ++ | ++ | ++ | ++ | |
| Secondary bronchus | ++ | ++ | ++ | ++ | |
| Parabronchus atria | ± | ± | ± | ± | |
| Air capillaries | + | ± | ± | ± | |
| Air sac | ++ | ++ | ++ | ++ | |
| Eurasian wigeon | Trachea | + | ++ | ++ | ++ |
| Primary bronchus | + | ++ | ++ | ++ | |
| Secondary bronchus | ++ | ++ | ++ | ++ | |
| Parabronchus atria | ± | ± | ± | ± | |
| Air capillaries | + | ± | ± | ± | |
| Air sac | ++ | ++ | ++ | ++ | |
| Tufted duck | Trachea | ++ | + | ++ | ++ |
| Primary bronchus | ++ | ++ | ++ | ++ | |
| Secondary bronchus | ++ | ++ | ++ | ± | |
| Parabronchus atria | + | ± | ± | ± | |
| Air capillaries | + | ± | ± | ± | |
| Air sac | ++ | + | ++ | ++ | |
| Graylag goose | Trachea | ++ | ++ | ++ | ++ |
| Primary bronchus | nd | nd | nd | nd | |
| Secondary bronchus | ++ | ++ | ++ | ++ | |
| Parabronchus atria | ++ | + | + | + | |
| Air capillaries | + | + | ++ | + | |
| Air sac | ++ | ++ | ++ | ++ | |
| Chicken | Trachea | ++ | ++ | ++ | ++ |
| Primary bronchus | + | ++ | ++ | ++ | |
| Secondary bronchus | + | ++ | ++ | ++ | |
| Parabronchus atria | ++ | + | ++ | ++ | |
| Air capillaries | + | − | ± | + | |
| Air sac | ++ | ++ | ++ | ++ | |
| Samples | Eurasian Wigeons | Barnacle Geese | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W1 | W2 | W3 | W4 | W5 | G1 | G2 | G3 | G4 | G5 | G6 | G7 | G8 | G9 | |
| Virus | H5N8 | H5N1 | H5N8 | H5N8 | H5N8 | H5N8 | H5N8 | H5N8 | H5N8 | H5N8 | H5N8 | H5 | H5N8 | H5N8 |
| Pharyngeal swab | 22 | 30 | 28 | 33 | 23 | np | 27 | 29 | 33 | np | 33 | np | 22 | 27 |
| Cloacal swab | 30 | 27 | 26 | 24 | 27 | 28 | 29 | np | 28 | 30 | 29 | 34 | 28 | 28 |
| Lung | 25 | 32 | 23 | 31 | 27 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Liver | 29 | 30 | 35 | 34 | 22 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Heart | 23 | 30 | 23 | 32 | 27 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Jejunum | 27 | 30 | 23 | 30 | 22 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Brain | 28 | 34 | 24 | 32 | 28 | nd | nd | nd | nd | nd | nd | nd | nd | nd |
| Number of Birds with Gross (G) and Histological (H) Lesions in the: | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Respiratory System | Digestive System | Other Systems | |||||||||||||||
| Lung | Air Sac | Intestine | Pancreas | Liver | Brain * | Heart | Kidney | ||||||||||
| Species | No of Birds | G | H | G | H | G | H | G | H | G | H | G | H | G | H | G | H |
| Eurasian wigeon | 5 | 3 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 3 | 1 | 1 | 0 | 0 |
| Barnacle goose | 9 * | 3 | 2 | 0 | 0 | 0 | 0 | 5 | 3 | 0 | 1 | 3 | 4 | 2 | 0 | 0 | 1 |
| Number of Birds Expressing Influenza Virus Antigen in a Cell Type of an Organ | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Respiratory System | Digestive System | Other Systems | ||||||||||||||||
| Lung | Air Sac | Intestine | Pancreas | Liver | Brain * | Heart | ||||||||||||
| Species | No of Birds | EP | E | N | EP | E | N | EP | E | N | E | EP | E | H | E | N | E | M |
| Eurasian wigeon | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 3 | 0 | 1 |
| Barnacle goose | 9 * | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 4 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caliendo, V.; Leijten, L.; van de Bildt, M.; Germeraad, E.; Fouchier, R.A.M.; Beerens, N.; Kuiken, T. Tropism of Highly Pathogenic Avian Influenza H5 Viruses from the 2020/2021 Epizootic in Wild Ducks and Geese. Viruses 2022, 14, 280. https://doi.org/10.3390/v14020280
Caliendo V, Leijten L, van de Bildt M, Germeraad E, Fouchier RAM, Beerens N, Kuiken T. Tropism of Highly Pathogenic Avian Influenza H5 Viruses from the 2020/2021 Epizootic in Wild Ducks and Geese. Viruses. 2022; 14(2):280. https://doi.org/10.3390/v14020280
Chicago/Turabian StyleCaliendo, Valentina, Lonneke Leijten, Marco van de Bildt, Evelien Germeraad, Ron A. M. Fouchier, Nancy Beerens, and Thijs Kuiken. 2022. "Tropism of Highly Pathogenic Avian Influenza H5 Viruses from the 2020/2021 Epizootic in Wild Ducks and Geese" Viruses 14, no. 2: 280. https://doi.org/10.3390/v14020280