
Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sampling and Study Area
2.3. Molecular Analysis and Virus Isolation
2.4. Sequencing and Phylogenetic Analysis
2.5. Serological Analysis
3. Results
3.1. Sampling
3.2. Serology
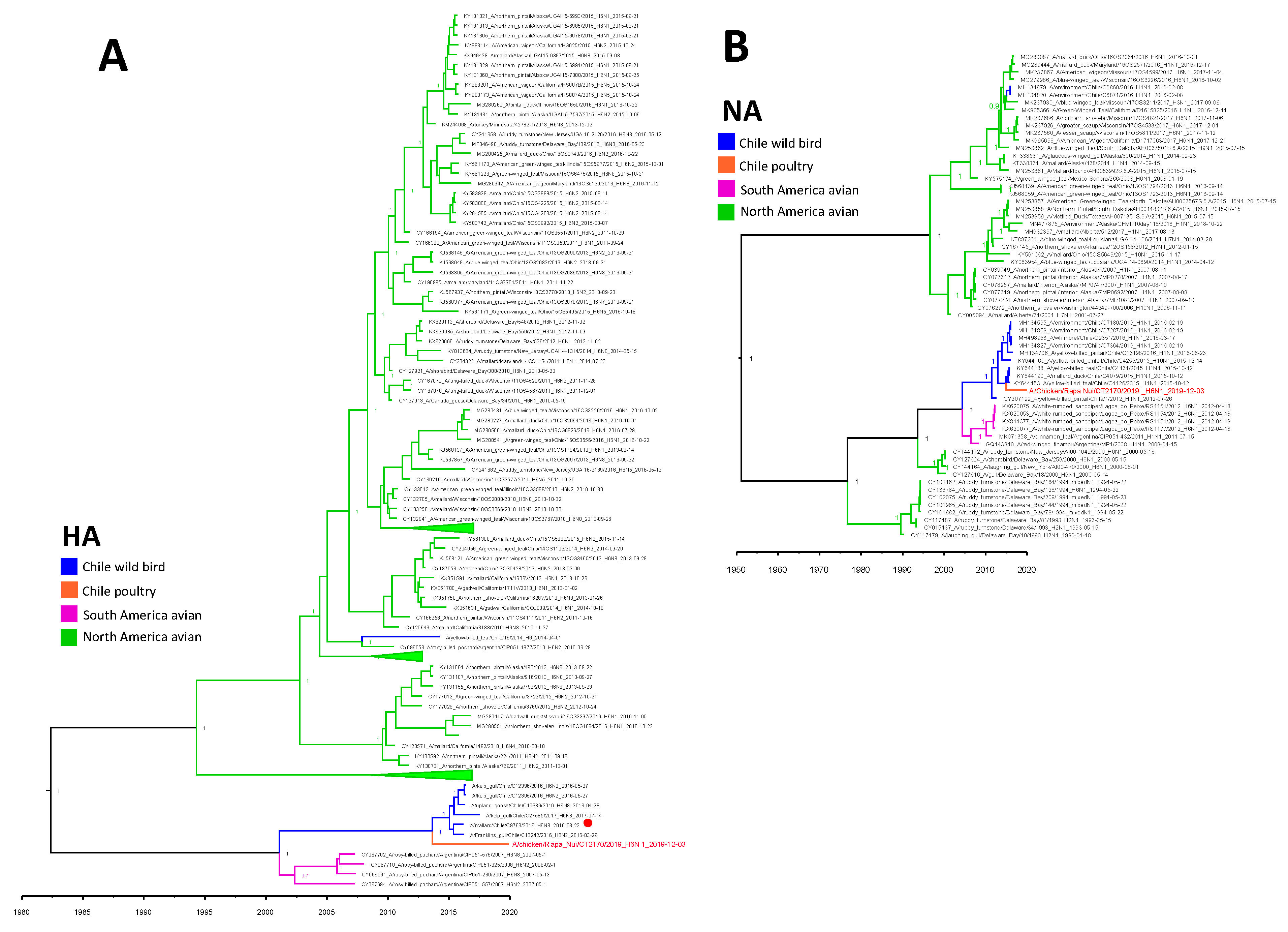
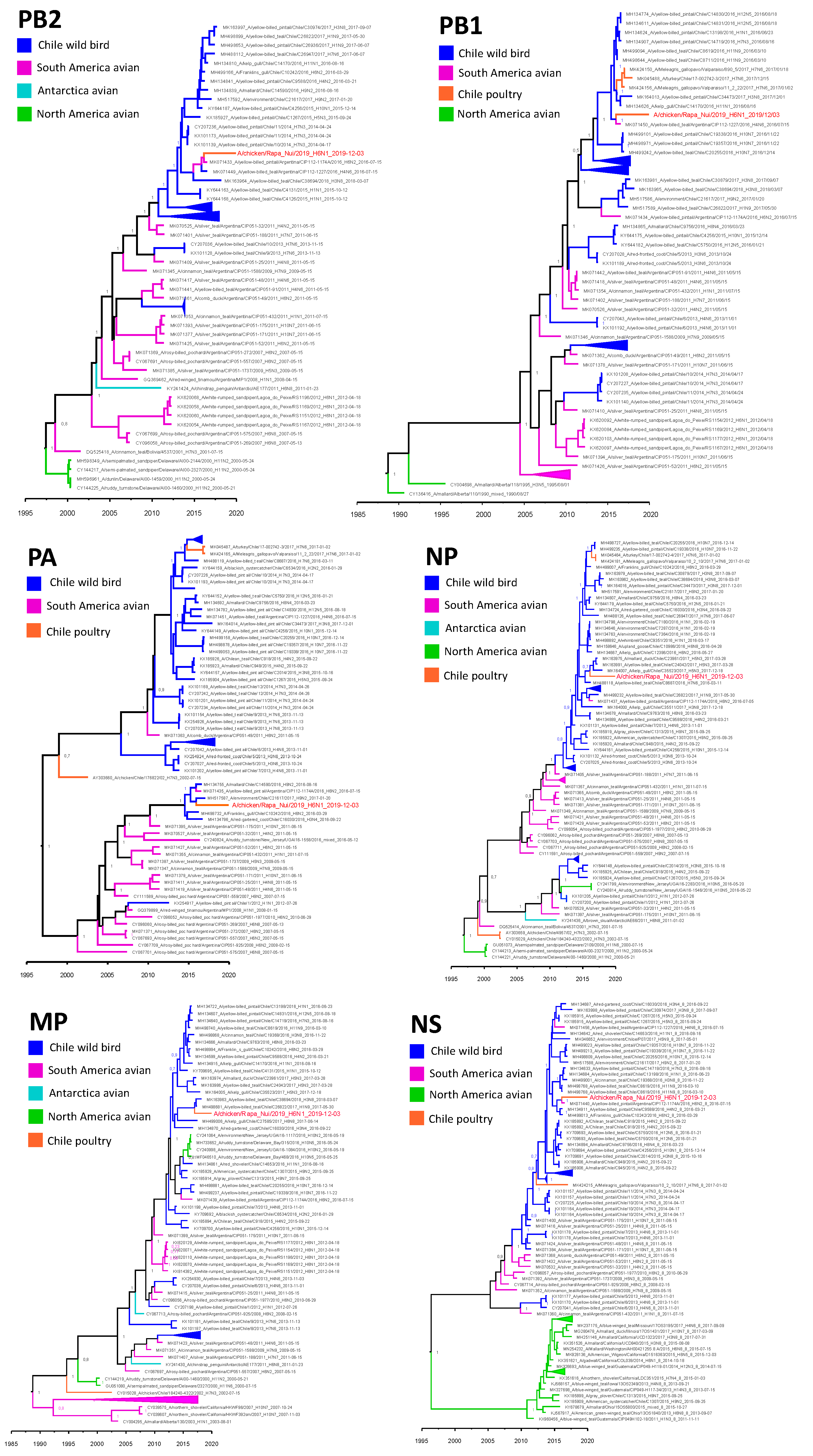
3.3. Molecular and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morin, C.W.; Stoner-Duncan, B.; Winker, K.; Scotch, M.; Hess, J.J.; Meschke, J.S.; Ebi, K.L.; Rabinowitz, P.M. Avian influenza virus ecology and evolution through a climatic lens. Environ. Int. 2018, 119, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Nao, N.; Yamagishi, J.; Miyamoto, H.; Igarashi, M.; Manzoor, R.; Ohnuma, A.; Tsuda, Y.; Furuyama, W.; Shigeno, A.; Kajihara, M.; et al. Genetic Predisposition to Acquire a Polybasic Cleavage Site for Highly Pathogenic Avian Influenza Virus Hemagglutinin. mBio 2017, 8, e02298-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krauss, S.; Walker, D.; Pryor, S.P.; Niles, L.; Cenghong, L.; Hinshaw, C.S.; Webster, R.G. Influenza A viruses of migrating wild aquatic birds in North America. Vector-Borne Zoonot. Dis. 2004, 4, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Pereda, A.J.; Uhart, M.; Perez, A.A.; Zaccagnini, M.E.; La Sala, L.; Decarre, J.; Goijman, A.; Solari, L.; Suarez, R.; Craig, M.I.; et al. Avian influenza virus isolated in wild waterfowl in Argentina: Evidence of a potentially unique phylogenetic lineage in South America. Virology 2008, 378, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.I.; Pollett, S.; Ghersi, B.; Silva, M.; Simons, M.P.; Icochea, E.; Gonzalez, A.; Segovia, K.; Kasper, M.R.; Montgomery, J.M.; et al. The Genetic Diversity of Influenza A Viruses in Wild Birds in Peru. PLoS ONE 2016, 11, e0146059. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Karlsson, E.A.; Pamela Freiden, P.; Sharp, B.; Di Pillo, F.; Jorge, E.; Osorio, E.; Hamilton, C.; Schultz-Cherry, S. Wild birds in Chile Harbor diverse avian influenza A viruses. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Rimondi, A.; Gonzalez-Reiche, A.; Olivera, V.S.; Decarre, J.; Castresana, G.J.; Romano, M.; Nelson, M.I.; van Bakel, H.; Pereda, A.J.; Ferreri, L.; et al. Evidence of a fixed internal gene constellation in influenza A viruses isolated from wild birds in Argentina (2006–2016). Emerg. Microbes Infect. 2018, 7, 1–13. [Google Scholar] [CrossRef]
- Ruiz, S.; Jimenez-Bluhm, P.; Di Pillo, F.; Baumberger, C.; Galdames, P.; Marambio, V.; Salazar, C.; Mattar, C.; Sanhueza, J.; Schultz-Cherry, S.; et al. Temporal dynamics and the influence of environmental variables on the prevalence of avian influenza virus in main wetlands in central Chile. Transbound. Emerg. Dis. 2020, 68, 1601–1614. [Google Scholar] [CrossRef]
- Bravo-Vasquez, N.; Di Pillo, F.; Lazo, A.; Jiménez-Bluhm, P.; Schultz-Cherry, S.; Hamilton-West, C. Presence of influenza viruses in backyard poultry and swine in El Yali wetland, Chile. Prev. Vet. Med. 2016, 134, 211–215. [Google Scholar] [CrossRef]
- Jimenez-Bluhm, P.; Di Pillo, F.; Bahl, J.; Osorio, J.; Schultz-Cherry, S.; Hamilton-West, C. Circulation of influenza in backyard productive systems in central Chile and evidence of spillover from wild birds. Prev. Vet. Med. 2018, 153, 1–6. [Google Scholar] [CrossRef]
- Bravo-Vasquez, N.; Baumberger, C.; Jimenez-Bluhm, P.; Di Pillo, F.; Lazo, A.; Sanhueza, J.; Schultz-Cherry, S.; Hamilton-West, C. Risk factors and spatial relative risk assessment for influenza A virus in poultry and swine in backyard production systems of central Chile. Vet. Med. Sci. 2020, 6, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concha, R. El Presente Cultural y Turístico en Isla de Pascua. Boletín OPCA 2014. pp. 38–47. Available online: https://opca.uniandes.edu.co/el-presente-cultural-y-turistico-en-isla-de-pascua/ (accessed on 4 November 2014).
- Jaramillo, A.; Johnson, M.T.; Rothfels, C.J.; Johnson, R.A. The native and exotic avifauna of Easter Island: Then and now. Bol. Chil. Ornitol. Unión Ornitól. Chile 2009, 14, 8–21. [Google Scholar]
- Jarman, C.L.; Larsen, T.; Hunt, T.; Lipo, C.; Solsvik, R.; Wallsgrove, N.; Ka’Apu-Lyons, C.; Close, H.; Popp, B.N. Diet of the prehistoric population of Rapa Nui (Easter Island, Chile) shows environmental adaptation and resilience. Am. J. Phys. Anthr. 2017, 164, 343–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, M.; Caceres, P. Sobre las aves de Isla de Pascua. Bol. Mus. Nac. Hist. Nat. 2010, 59, 75–95. [Google Scholar]
- Harrisson, T. Easter Island: A Last Outpost. Oryx 1971, 11, 111–116. [Google Scholar] [CrossRef]
- Hoque, M.A.; Burgess, G.W.; Cheam, A.L.; Skerratt, L.F. Epidemiology of avian influenza in wild aquatic birds in a biosecurity hotspot, North Queensland, Australia. Prev. Vet. Med. 2015, 118, 169–181. [Google Scholar] [CrossRef]
- Senne, D.A. Avian Influenza in the Western Hemisphere Including the Pacific Islands and Australia. Avian Dis. 2003, 47 (Suppl. 3), 798–805. [Google Scholar] [CrossRef]
- Stanislawek, W.L.; Wilks, C.R.; Meers, J.; Horner, G.W.; Alexander, D.J.; Manvell, R.J.; Kattenbelt, J.A.; Gould, A.R. Avian paramyxoviruses and influenza viruses isolated from mallard ducks (Anas platyrhynchos) in New Zealand. Arch. Virol. 2002, 147, 1287–1302. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. CDC Protocol of Realtime RTPCR for Influenza A (H1N1); Centers for Disease Control and Prevention: Atlanta, GA, USA, 2009.
- Shu, B.; Wu, K.-H.; Emery, S.; Villanueva, J.; Johnson, R.; Guthrie, E.; Berman, L.; Warnes, C.; Barnes, N.; Klimov, A.; et al. Design and Performance of the CDC Real-Time Reverse Transcriptase PCR Swine Flu Panel for Detection of 2009 A (H1N1) Pandemic Influenza Virus. J. Clin. Microbiol. 2011, 49, 2614–2619. [Google Scholar] [CrossRef] [Green Version]
- Moresco, K.A.; Stallknecht, D.E.; Swayne, D. Evaluation and Attempted Optimization of Avian Embryos and Cell Culture Methods for Efficient Isolation and Propagation of Low Pathogenicity Avian Influenza Viruses. Avian Dis. 2010, 54, 622–626. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Kaplan, B.S.; Russier, M.; Jeevan, T.; Marathe, B.; Govorkova, E.; Russell, C.J.; Kim-Torchetti, M.; Choi, Y.K.; Brown, I.; Saito, T.; et al. Novel Highly Pathogenic Avian A(H5N2) and A(H5N8) Influenza Viruses of Clade 2.3.4.4 from North America Have Limited Capacity for Replication and Transmission in Mammals. mSphere 2016, 1, e00003-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prjibelski, A.; Antipov, D.; Meleshko, D.; Lapidus, A.; Korobeynikov, A. Using SPAdes De Novo Assembler. Curr. Protoc. Bioinform. 2020, 70, e102. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, F.; Miller, W.; Myres, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Bao, Y.; Bolotov, P.; Dernovoy, D.; Kiryutin, B.; Zaslavsky, L.; Tatusova, T.; Ostell, J.; Lipman, D. The Influenza Virus Resource at the National Center for Biotechnology Information. J. Virol. 2008, 82, 596–601. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. c1979–c2000. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Kingman, J.F.C. The coalescent. Stochastic processes and their applications. Stoch. Proc. Appl. 1982, 13, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Kishino, H.; Yano, T.-A. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed Phylogenetics and Dating with Confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Blanvillain, C.; Saavedra, S.; Withers, T.; Votýpka, J.; Laroucau, K.; Lowenski, S.; Modrý, D. Screening of diseases in wild exotic birds on Tahiti Island—implications for French Polynesian conservation. Pac. Conserv. Biol. 2021, 27, 284. [Google Scholar] [CrossRef]
- Brioudes, A.; Warner, J.; Hedlefs, R.; Gummow, B. A review of domestic animal diseases within the Pacific Islands region. Acta Trop. 2014, 132, 23–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandegrift, K.J.; Sokolow, S.; Daszak, P.; Kilpatrick, A.M. Ecology of avian influenza viruses in a changing world. Ann. New York Acad. Sci. 2010, 1195, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M. Controlling avian influenza infections: The challenge of the backyard poultry. J. Mol. Genet. Med. 2009, 3, 119–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yancovic, S. Aves de Rapa Nui; Manu Project Ediciones: Santiago, Chile, 2020. [Google Scholar]



{kind=link}
{kind=link}
| Protein Coding Segment | Closest Reference Sequence | tMRCA of A/Chicken/Rapa Nui/CT2170/2019 to Closest Reference Sequence (95% HDP) | Country of Isolation | Nucleotide and Amino Acid Identity (%) | |
|---|---|---|---|---|---|
| PB2 | MK071433-A/yellow-billed pintail/Argentina/CIP112-1174A/2016 (H6N2) | 2016.1474 (2015.7938, 2016.49) | Argentina | 98.5 | 100 |
| PB1 | MK071450-A/yellow-billed teal/Argentina/CIP112-1227/2016 (H4N6) | 2015.9905 (2015.5587, 2016.3852) | Argentina | 97.7 | 99.4 |
| PA | MH517587-A/environment/Chile/C21617/2017 (H9N2) | 2015.9333 (2015.2742, 2016.5603) | Chile | 88.9 | 96.8 |
| HA | MH499098-A/Franklins gull/Chile/C10242/2016 (H6N2) | 2013.6342 (2011.8845, 2015.0703) | Chile | 96.1 | 98.5 |
| NP | MK163991-A/yellow-billed teal/Chile/C24042/2017 (H5N3) | 2016.1022 (2015.5233, 2016.6413) | Chile | 99.7 | 99.5 |
| NA | KY644153-A/yellow-billed teal/Chile/C4126/2015 (H1N1) | 2014.972 (2014.318, 2015.6045) | Chile | 98.2 | 98.5 |
| MP | MH498681-A/yellow-billed teal/Chile/C26822/2017 (H1N9) | 2016.9625 (2016.3621, 2017.4081) | Chile | 99.4 | 100 |
| NS | MK071440-A/yellow-billed pintail/Argentina/CIP112 CIP112-1174A/2016 (H6N2) | 2016.0634 (2015.5987, 2016.4783) | Argentina | 98.8 | 99.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Pillo, F.; Baumberger, C.; Salazar, C.; Galdames, P.; Ruiz, S.; Sharp, B.; Freiden, P.; Tan, S.; Schultz-Cherry, S.; Hamilton-West, C.; et al. Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia. Viruses 2022, 14, 718. https://doi.org/10.3390/v14040718
Di Pillo F, Baumberger C, Salazar C, Galdames P, Ruiz S, Sharp B, Freiden P, Tan S, Schultz-Cherry S, Hamilton-West C, et al. Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia. Viruses. 2022; 14(4):718. https://doi.org/10.3390/v14040718
Chicago/Turabian StyleDi Pillo, Francisca, Cecilia Baumberger, Carla Salazar, Pablo Galdames, Soledad Ruiz, Bridgett Sharp, Pamela Freiden, Shaoyuan Tan, Stacey Schultz-Cherry, Christopher Hamilton-West, and et al. 2022. "Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia" Viruses 14, no. 4: 718. https://doi.org/10.3390/v14040718
APA StyleDi Pillo, F., Baumberger, C., Salazar, C., Galdames, P., Ruiz, S., Sharp, B., Freiden, P., Tan, S., Schultz-Cherry, S., Hamilton-West, C., & Jimenez-Bluhm, P. (2022). Novel Low Pathogenic Avian Influenza H6N1 in Backyard Chicken in Easter Island (Rapa Nui), Chilean Polynesia. Viruses, 14(4), 718. https://doi.org/10.3390/v14040718