Abstract
Some people, known as HIV-exposed seronegative (HESN) individuals, remain uninfected despite high levels of exposure to HIV. Understanding the mechanisms underlying their apparent resistance to HIV infection may inform strategies designed to protect against HIV infection. Natural Killer (NK) cells are innate immune cells whose activation state depends on the integration of activating and inhibitory signals arising from cell surface receptors interacting with their ligands on neighboring cells. Inhibitory NK cell receptors use a subset of major histocompatibility (MHC) class I antigens as ligands. This interaction educates NK cells, priming them to respond to cells with reduced MHC class I antigen expression levels as occurs on HIV-infected cells. NK cells can interact with both autologous HIV-infected cells and allogeneic cells bearing MHC antigens seen as non self by educated NK cells. NK cells are rapidly activated upon interacting with HIV-infected or allogenic cells to elicit anti-viral activity that blocks HIV spread to new target cells, suppresses HIV replication, and kills HIV-infected cells before HIV reservoirs can be seeded and infection can be established. In this manuscript, we will review the epidemiological and functional evidence for a role for NK cells in protection from HIV infection.
1. Introduction
Some people remain HIV seronegative despite high levels of exposure to this virus. They are referred to as HIV-exposed seronegative (HESN) individuals [1]. Studying HESNs may provide information that enhances our understanding of the mechanisms that underlie their apparent resistance to HIV infection. This information has the potential to guide the development of novel prophylactic HIV therapies.
Several groups have screened HESNs for the presence of adaptive HIV-specific T cells and antibody responses. Such responses have been found in many but not all cohorts of HESN subjects [2,3,4,5,6,7]. HIV-specific T cell responses in HESN were often of a low breadth and magnitude, and their presence did not always predict maintenance of seronegative status [8,9,10,11,12]. The STEP HIV vaccine trial, designed to induce strong T cells responses, failed to protect against HIV infection [13,14]. Furthermore, HIV-specific memory responses, even if present in HESNs, would be expected to take days following an exposure to HIV to develop into effector responses able to kill HIV-infected cells and prevent the establishment of HIV infection. In an animal model for HIV infection in humans, i.e., rhesus macaques infected intrarectally with simian immunodeficiency virus, starting antiretroviral therapy (ART) as early as day 3 post infection failed to prevent viral load rebound when ART was stopped 24 weeks later [15]. This observation is consistent with viral reservoirs refractory to eradication being seeded early during the eclipse phase of HIV infection before virions and infected cells are detectable in the circulation [16]. Together, these observations suggest that adaptive HIV-specific immune responses induced by exposure to HIV are likely to be generated too late to prevent the seeding of viral reservoirs and establishment of HIV infection resulting from exposure to HIV [15,16]. Thus, adaptive immune responses detected in HESN more likely identify individuals as having been previously exposed to the virus rather than protect from subsequent HIV exposures [17].
Innate Natural Killer (NK) cell immune responses with anti-HIV activity are induced more rapidly than adaptive immune responses [18]. NK cells, as components of the innate immune response, have the potential to protect against HIV infection. NK cells play an important role in anti-viral and anti-tumor activity [19,20]. They are primed to respond rapidly to HIV-infected cells, which have increased expression of ligands for germline-encoded activating NK receptors (NKRs) as occurs on HIV-infected cells [21]. HIV-infected cells downmodulate their cell surface expression of major histocompatibility complex (MHC) class I antigens [22,23,24,25]. Upon activation, NK cells secrete cytokines with anti-viral activity and chemokines that block the CCR5 co-receptor for HIV entry into new target cells and degranulate, which can lead to the lysis of infected cells [19,26,27,28]. Whether NK cells respond to stimuli or remain resting depends on the integration of signals received from inhibitory and activating NKRs [29]. Signals from inhibitory NKRs tend to dominate those from activating NKRs [29,30,31]. NK cells can be activated directly when the integration of signals from inhibitory and activating NKRs upon binding their ligands on neighboring cells favors activation.
In this review, we will focus on how KIR/HLA combinations that contribute to NK cell education influence NK cell responses to prevent HIV infection.
2. Basis of NK Cell Education
During their development, NK cells are educated through the interaction of their germline encoded inhibitory NKRs with self MHC class I (or human leukocyte antigen (HLA)) ligands expressed on adjoining cells [32,33,34,35]. A consequence of NK cell education is tolerance to self-cells as the interaction of inhibitory NKRs with their HLA ligands transduces inhibitory signals that suppress NK cell activation [36,37]. However, education also primes NK cells to respond rapidly to aberrant cells with reduced levels of HLA ligands due to virus infection, tumor transformation, or cell damage [35,38]. HIV infection downmodulates cell surface MHC expression through the action of HIV negative regulatory factor (Nef) and viral protein U (Vpu), which are HIV-encoded accessory proteins with multiple functions [22,23,24,25].
HLA-E interacts with the inhibitory NKR NKG2A to educate NK cells [39]. Many HLA allotypes are ligands for inhibitory killer immunoglobulin-like receptors (KIRs). KIRs are a large family of receptors that include both activating and inhibitory counterparts expressed on NK cells and other lymphocyte subsets [40]. Binding of inhibitory KIRs to their HLA ligands transduces intracellular inhibitory signals that support tolerance to healthy HLA-expressing autologous cells [29,30,31]. The KIR and HLA regions are polygenic and characterized by a high degree of polymorphism. They are encoded on chromosomes that segregate independently from each other allowing for a great diversity of receptor ligand combinations. [41,42,43]. NKRs, including KIRs, are expressed stochastically on overlapping NK cell subsets. Thus, both educated and uneducated NK cells can co-exist as can NK cells with varied levels of education and responses to stimulation, due to allelic KIR and HLA variation [35,44,45]. The inhibitory KIR+ NK cell subsets unable to interact with self HLA during development remain uneducated and are hyporesponsive unless activating signals predominate [32,34,46,47]. The depth of NK cell education, which depends on the number of inhibitory NKR-ligand interactions, inhibitory KIR/HLA ligand density and the affinity of the interactions between the two during the education process, correlates with the level of activation mature NK cells achieve when they encounter cells with aberrant HLA expression levels [33,35,38].
3. KIR NK Receptors
The genetic KIR region is located within human chromosome 19q13.4. Information generated using full haplotype multiple sequence alignment had identified 14 loci and 1 intergenic locus containing 9 genes in this region [48]. KIR nomenclature reflects their protein structure [49]. After “KIR” is the number of extracellular immunoglobulin-like domains (2D or 3D), followed (S or L) for short or long intracellular domains characteristic of activating or inhibitory receptors, respectively. P refers to pseudogenes in the KIR nomenclature. A number follows, which indicates the sequence in which each allele or protein was named [50]. The long cytoplasmic domains of inhibitory receptors have an immunoreceptor tyrosine-based inhibitory motif that recruits SRC homology 2 (SH2) domain containing protein tyrosine phosphatases such as SHP1 and SHP2 [30]. The short cytoplasmic domain of activating receptors associates with adaptor molecules with immunoreceptor tyrosine-based activating motifs that recruit protein tyrosine kinases [29].
Among the inhibitory KIRs, the ligands for KIR3DL1 are a subset of HLA-A and -B antigens having an HLA-Bw4 motif, defined by amino acids present at positions 77–83 of the HLA heavy chain (Table 1) [51,52]. A dimorphism at position 80 of the Bw4 heavy chain divides these isotypes into those with an isoleucine (*80I) or threonine (*80T) at this position, which influences HLA cell surface expression and binding affinity to KIR3DL1 [35,45,53]. HLA-Bw6 allotypes do not interact with KIR3DL1 and NK cells from HLA-Bw6 homozygotes expressing KIR3DL1, as their only inhibitory KIR remain uneducated [32]. KIR3DL1 allotypes can be categorized as KIR3DL1-null with no detectable cell surface expression, KIR3DL1-low and KIR3DL1-high allotype groups based on their cell surface expression levels [35,54]. In general, KIR3DL1-high receptors have a higher affinity for Bw4*80I than Bw4*80T allotypes while the avidity of KIR3DL1-low receptors to these two HLA-Bw4 groups is similar [35]. The inhibitory KIRs KIR2DL1, KIR2DL2, and KIR2DL3 recognize HLA-C allotypes, which can be dichotomized into C1 and C2 groups. C1 allotypes have an asparagine at position 80 of their heavy chain and are ligands for KIR2DL3. C2 group allotypes have a lysine at this position and are ligands for inhibitory KIR2DL1 and activating KIR2DS1 receptors [44,55,56]. KIR2DL2 is an intermediate receptor, which also binds C1 allotypes and some C2 allotypes (Table 1) [44,55,57].

Table 1.
Receptor-ligand pairs contributing to NK cell education.
Historically viewed, the human KIR system segregates into two groups of haplotypes (A and B), which are distinguished, at least in part, by their activating KIR gene content (Figure 1) [42,58,59,60,61]. The group A haplotypes are dominated by genes encoding inhibitory receptors [60]. The exception to this is the KIR2DS4 locus, which encodes an activating receptor that is often truncated and not cell surface expressed [62,63]. The group B haplotypes encode more variable and greater numbers of genes encoding inhibitory receptors and varying numbers activating receptors [60]. KIR haplotypes share three framework genes and one pseudogene. KIR3DL2 and KIR2DL3 are located at the centromeric and telomeric ends of the KIR region while the pseudogene KIR3DP1 and KIR2DL4 are in the middle, together framing two regions of variability of the KIR locus [64]. A 14 kb intergenic region separates KIR2DP1 and KIR2DL4 and divides the KIR locus into centromeric and telomeric regions [40]. This intergenic region is a site of reciprocal recombination, which allows the centromeric and telomeric regions to reassort into combinations that form new variant KIR haplotypes [58,65]. The genes in the telomeric and centromeric regions are in linkage disequilibrium with each other and tend to be inherited together [59,61]. Full haplotype DNA multiple sequence alignments are now available for 68 KIR haplotypes [48]. This information has refined and confirmed the sequence alignments of several KIR haplotypes [48,59,61]. Of the 13 KIR haplotypes identified by full KIR region sequencing, 6 were found in only one person [48]. More than 90% of the 68 fully sequenced KIR haplotypes are composed of one of four centromeric and one of three telomeric motifs (Figure 1). The less frequent KIR haplotypes include gene duplications, deletions, insertions, and hybridizations of KIR genes present in the more frequent haplotypes [59,61].
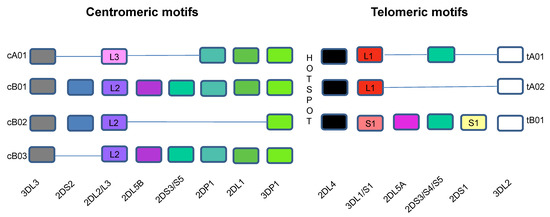
Figure 1.
Killer Immunoglobulin-like Receptor Haplotypes. KIR genes are organized into haplotypes A and B. KIR genes shown in the bottom row are designated without “KIR” before their names. The different colors are used to identify different genes. Haplotype A is comprised of four framework genes present in most KIR haplotypes (KIR3DL3 at the centromeric end, KIR3DL2 at the telomeric end and the pseudogene KIR3DP1 and KIR2DL4 in the middle) plus genes encoding inhibitory KIRs KIR2DL1, KIR2DL3, KIR3DL1, activating KIR KIR2DS4 and pseudogene KIR2DP1. The more diverse group B haplotypes include the framework genes with various combinations of genes encoding inhibitory KIRs KIR2DL2 and KIR2DL5A/B and activating KIRs KIR2DS1, KIR2DS2, KIR2DS3, KIR2DS5, and KIR3DS1. More than 90% of the 68 haplotypes that have been sequenced to date using full haplotype multiple-sequence alignments are composed of one of four centromeric and one of three telomeric KIR motifs that include combinations of KIR genes in linkage disequilibrium with each other, which are usually inherited together. The intergenic region between KIR3DP1 and KIR2DL4 is a hotspot between the centromeric and telomeric regions, which allows for frequent recombination between the two regions. The centromeric region is delimited by the framework genes KIR3DL3 and KIR3DP1, while the telomeric region is delimited by framework genes KIR2DL4 and KIR3DL2. KIR2DP1 and KIR2DP1 are pseudogenes. cA01 = genes in the centromeric region of KIR haplotype A 1; cB01 = genes in the centromeric region of KIR haplotype B 1; cB02 = genes in the centromeric region of KIR haplotype B 2; cB03 = genes in the centromeric region of KIR haplotype B 3; tA01 = genes in the telomeric region of KIR haplotype A 1; tA02 = genes in the telomeric region of KIR haplotype A 2; tB01 = genes in the telomeric region of KIR haplotype B 1.
4. Epidemiological Studies Supporting a Role for NK Cells in Protection from HIV Infection
One of the first publications implicating NK cells in protection from HIV infection showed, in a cohort of highly HIV exposed Vietnamese injection drug users (IDUs), that IDU HESN persons had enhanced NK cell responses to HLA null cells compared to individuals at low risk for HIV infection and IDU persons living with HIV (PLWH) [66]. The elevated NK cell activity in IDU HESN individuals was higher than that in IDU PLWH for NK cells isolated from IDU PLWH at time points both before and after seroconversion [66]. The observation that lower levels of NK cell activity in IDU PLWH predated HIV seroconversion suggested that NK cell-mediated protection from HIV infection was genetically determined.
Epidemiological studies conducted on samples from longitudinally followed PLWH enrolled in the Multicenter AIDS Cohort Study found an association between certain KIR3DL1/HLA and KIR3DS1/HLA combinations with time to the acquired immunodeficiency syndrome (AIDS) (using the 1987 US Centers for Disease Control and Prevention’s definition for AIDS), time to CD4 counts of <200 cells/mm3 and HIV viral load control [67,68]. In these studies, Bw6 homozygotes with uneducated KIR3DL1+ NK cells served as controls for the effect of educated KIR3DL1+ NK cells on HIV disease outcomes. Among the combined KIR/HLA genotypes associated with slow HIV disease progression were co-carriage of the KIR3DL1-null allele KIR3DL1*004 with HLA-Bw4, co-carriage of alleles encoding KIR3DL1-high and KIR3DL1-low allotypes with HLA-B*57 and HLA-B*27 and alleles encoding KIR3DS1 and Bw4*80I [67,68]. These studies implicated a role for NK cells in the natural history of HIV infection. These findings also raised the possibility that genetic combinations encoding certain KIR/HLA combinations may have accounted for the higher NK cell activity seen in IDU HESN individuals than PLWH enrolled in the Vietnamese IDU cohort. This could be the case if the enhanced NK cell activity observed in IDU HESN persons compared to IDU PLWHs pre-seroconversion was due to the contribution of these combined genotypes to the potency of NK cell education, which would have been directly related to levels of NK cell functionality in response to HIV-infected cells present in parenteral exposures.
As innate immune cells, NK cells are primed to respond to HIV-infected cells by eliciting anti-viral activity at the earliest stages of HIV infection in a manner that may prevent the establishment of HIV infection [18,24,28,69]. This prompted an investigation of whether KIR/HLA combinations associated with slower rates of HIV disease progression were also associated with prevention from HIV infection. This question was addressed by comparing the proportion of longitudinally followed IDU HESN persons and recently HIV-infected individuals carrying various KIR/HLA combinations associated with slower time to AIDS [70,71,72,73]. Carriage of the KIR3DL1-high+HLA-B*57 genotype combination, which had the most potent effect on viral load control and slow time to AIDS in PLWH was significantly more frequent among IDU HESNs than in recently infected PLWH [67,70]. These observations implicated a role for potently educated NK cells in protection from HIV infection.
While the combination of at least one copy of KIR3DS1 with HLA Bw4*80I was associated with slower time to AIDS and viral load control, it was not associated with protection from HIV infection [68,71]. However, the carriage of the KIR3DS1 homozygous genotype was linked to a reduced risk of HIV infection [71]. This genotype was also associated with slower time to HIV seroconversion in a highly HIV-exposed longitudinally followed IDU HESN cohort [74]. The proportion of KIR3DL1/S1 heterozygotes and KIR3DL1 homozygotes did not differ between IDU HESNs and recently infected PLWH. Co-carriage of KIR3DS1 with HLA-Bw4*80I did not modify the effect on protection from infection conferred by the KIR3DS1 homozygous genotype [71]. Not all the KIR3DL1/S1/HLA combined genotypes associated with a slower time to AIDS were also associated with protection from infection. This could be due to the size of the IDU HESN and recently infected PLWH populations studies being too small to provide sufficient power to observe significant between-group differences in the frequency of all the KIR3DL1/S1/HLA combinations that were associated with slower time to AIDS in longitudinally followed PLWH. An alternative possibility is that the mechanisms of protection from HIV infection differ from those related to rate of HIV disease progression in PLWH.
5. The Contribution of NK Cell Education to Function
When HLA-null cells stimulate NK cells, negative signaling through inhibitory NKRs is interrupted while signals through activating NKRs persist [75]. In this scenario, NK cells are activated in accordance with the potency with which they were educated, revealing their functional potential. HLA-null cell stimulation of NK cells from HIV uninfected carriers of the KIR3DL1-high+B*57 genotype combination activated a higher frequency of NK cells to secrete interferon-γ (IFN-γ), the chemokine CCL4, and externalize CD107a, a marker for NK cell degranulation than NK cells from carriers of alleles encoding other KIR3DL1/HLA receptor/ligand pairs [76]. HLA-null cells also stimulated a higher frequency of NK cells from KIR3DL1*004+Bw4 carriers than Bw6 homozygotes to secrete IFN-γ and CCL4 and externalize CD107a, though this combination was not more frequent among IDU HESN individuals than in recently infected PLWH [77].
HIV-infected CD4 cells downmodulate cell surface HLA expression and upregulate the expression of ligands for activating NKRs, which together activate NK cells (Figure 2) [21,22,23,25,69]. NK cell activation by autologous HIV-infected cells was observed by gating on NK cells expressing a single inhibitory KIR by flow cytometry. The frequency of single positive KIR3DL1+ NK cells secreting IFN-γ and CCL4 and externalizing CD107a, all of which have anti-HIV activity, was higher in educated than uneducated, KIR3DL1+ NK cells from Bw6 homozygotes responding to autologous HIV-infected CD4 cell stimulation [69]. Expression levels of both the KIR3DL1 receptor and its HLA-B ligand and their binding affinity influenced single positive KIR3DL1+ NK cell education potency and responsiveness to autologous HIV-infected CD4 cells [35,69]. For example, NK cells from KIR3DL1-high+HLA-B*57 carriers had a superior ability to inhibit HIV replication in autologous infected CD4 cells compared to carriers of other KIR/HLA combinations and Bw6 homozygotes [28]. The inhibition of HIV replication was mediated, at least in part, by soluble CCL3, CCL4, and CCL5 chemokines secreted by activated NK cells because blocking all three of these chemokines restored HIV replication [28]. These three chemokines bind CCR5, the co-receptor for HIV entry, thus blocking the infection of new HIV susceptible CD4 target cells [27].
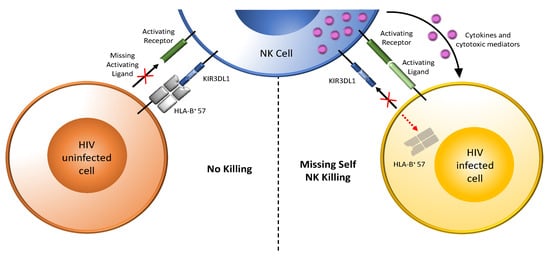
Figure 2.
Target recognition, tolerance, and missing-self recognition. NK cells recognize and kill their targets through the integration of signals received from inhibitory and activating receptors. They can discriminate between healthy cells (tolerance) and eliminate transformed or virally infected targets (killing). As shown to the left of the vertical dotted line that separates the figure into two parts, NK-cell tolerance depends on the interaction of several major histocompatibility complex (MHC) class I ligands (either classical, HLA-A, -B, or -C, or nonclassical, HLA-E) expressed by heathy cells with their inhibitory KIR or NKG2A receptors with minimal activation signals. Shown here as an example is expression of the MHC class I antigen HLA-B*57 on healthy cells engaging the inhibitory receptor KIR3DL1 on NK cells, transducing inhibitory signals that maintain NK cells in a resting state. HIV uninfected cells express few ligands for activating receptors. On the right, HIV-infected cells downmodulated HLA, abrogating inhibitory signals through KIR3DL1. HIV-infected cells also upregulate ligands for activating NK cell receptors. Loss of inhibitory signaling (as indicated by the “+” symbol over the ligand for inhibitory KIR3DL1) and gain of activating signaling activates NK cells to release cytokines and cytotoxic mediators, which kill HIV-infected cells. Dark green bar on the NK cell = activating NK cell receptor; light green bar on HIV infected cell (right panel) = ligand for an activating receptor (this bar in missing on HIV uninfected cells in left hand panel as indicated by the “+” symbol); blue three domain structure on the NK cell = inhibitory KIR3DL1 receptor; grey 4 domain structure on HIV uninfected cells (left-hand panel) = HLA-B*57, a ligand for inhibitory receptor KIR3DL1 (this HLA antigen is missing on the HIV infected cell in left hand panel as indicated by the “+” symbol); purple dots = cytokines and cytotoxic mediators.
IDU HESN individuals, compared to recently infected PLWH, were more likely to carry two copies of KIR3DS1 and one or two copies of a telomeric KIR tB01 motif and were less likely to carry a full length KIR2DS4*001-like allele [78]. A study comparing 25 IDU HESN persons with 19 IDU PLWH and 26 HIV uninfected individuals from the Vietnamese IDU cohort mentioned above found that HESN persons had KIR expression profiles encoding higher ratios of KIR3DS1:KIR3DL1 transcripts [79]. A higher prevalence of KIR3DS1 or lower frequency of KIR3DL1 alleles in HESN individuals than in HIV-susceptible subjects has been reported in several studies, and this is often in the absence of an association with HLA-Bw4 alleles [71,74,79,80,81]. Overall, these studies support the interpretation that HIV resistance may be due to NK cells that are more easily activated, which is consistent with carriage of a group B KIR haplotype in which larger numbers of genes encoding activating KIRs are present.
KIR3DS1 is found on the telomeric KIR tB01 motif in linkage disequilibrium with KIR2DL5A, KIR2DS1, and KIR2DS5 genes (Figure 1). Although these genes are in linkage disequilibrium, their gene products are expressed on NK cells in a stochastic fashion, allowing investigation of the functional potential of NK cells expressing various combinations of the KIRs encoded by these genes following stimulation with the HLA null cell line 721.221. NK cells that responded with the highest frequency of functional cells were KIR3DS1+. Co-expression of KIR2DL5, KIR2DS1, or KIR2DS5 did not modulate the frequency of responding KIR3DS1+ NK cells nor did KIR2DL5+, KIR2DS1+, and KIR2DS5+ NK cells respond to this stimulus any better than their KIR2DL5−, KIR2DS1−, or KIR2DS5− counterparts. Thus, of the KIR receptors encoded by genes present within the tB01 motif, KIR3DS1 contributes most to NK cell responsiveness to 721.221 HLA null cells [78].
Attempts to find ligands for KIR3DS1 among HLA-A, -B and -C allotypes, including HLA-Bw4 and HLA-Bw4*80I allotypes, using beads expressing single HLA antigens failed [82,83]. KIR3DS1 did interact with the HLA-Bw4*80I antigen, HLA-B*57, when certain HIV-derived peptides were present [84]. If the interaction of KIR3DS1 with HLA-B*57 was contributing to NK cell education, this interaction should tune down KIR3DS1+ NK cell responsiveness to 721.221 stimulation as occurs when the activating NKR KIR2DS1 interacts with HLA-C2 group antigens [85,86]. KIR3DS1 binds the open conformation of HLA-F. KIR3DS1+ NK cells are activated by their interaction with HLA-F, expressed on 721.221 cells [82,87,88,89]. NK cells, stimulated with the HLA-F expressing 721.221 cells and autologous HIV-infected CD4 cells, stimulated a higher frequency of NK cells to produce IFN-γ, CCL4, and CD107a than control stimuli [24,82,87]. Blocking this interaction with either antibodies to HLA-F or KIR3DS1-Fc chimeric proteins significantly reduced KIR3DS1+ NK cell activation. Since HLA-F is usually intracellular in resting cells, it would not be expected to contribute to NK cell education in vivo and tune down the responsiveness of KIR3DS1+ NK cells to HLA-F expressing cells [89]. However, HLA-F is cell surface expressed on activated cells, including HIV-infected CD4+ T cells [24,82,89]. The interaction of KIR3DS1 on NK cells with HLA-F on HIV-infected cells may explain why KIR3DS1+ NK cells are superior to KIR3DL1+ NK cells in suppressing HIV replication [82,90]. This interaction may provide a mechanistic explanation for the reduced risk of HIV infection observed in IDU HESN KIR3DS1 homozygotes, i.e., the rapid activation of these NK cells by interactions with HLA-F-expressing HIV-infected cells to induce anti-HIV functions.
NK cells from donors with no KIR2DS4 gene, or who carry only truncated KIR2DS4*003-like alleles do not express the KIR2DS4 receptor on their cell surface, while NK cells from carriers of the full length KIR2DS4*001-like alleles express this receptor on a subset of their NK cells. KIR2DS4 was associated with poor outcome in the context of HIV infection such as higher viral load, lower CD4 counts and HIV transmission in HIV discordant couples [91,92]. This observation was confirmed by Olvera et al. in another group of PLWH from Lima, Peru and may have depended on the co-expression of HLA-Cw4, a presumed ligand for this receptor [93]. Expressed full length KIR2DS4 is linked with reduced resistance to HIV infection [78]. The mechanism underlying this finding is not understood. KIR2DS4 is an activating KIR that is the product of gene conversion between KIR2DL genes and the KIR3DL2 gene that has led to a reduced ability to recognize HLA-C ligands characteristic of KIR2DL receptors and an increased ability to recognize HLA-A*11:02 and HLA-A*03 ligands, the presumed ligands for KIR3DL2 [94]. Further investigation is needed to understand whether, and if so how, KIR2DS4 expression is associated with negative outcomes in the context of HIV infection and HIV exposure. The possibility that the impact of expression of KIR2DS4 on poor HIV outcomes is due to other genes in linkage disequilibrium with KIR2DS4 has not been excluded.
Together, these observations implicate a role for NK cell activation in protection from HIV infection. This can occur through the potent education of NK cells that become activated upon loss of inhibition mediated by the downmodulation of ligands for inhibitory NKR receptors on autologous HIV-infected CD4+ T cells combined with the upregulation of ligands for activating receptors on these cells. KIR3DS1+ NK cells can also be activated by the upregulation of HLA-F and other ligands for activating receptor on HIV-infected CD4 cells [21,24]. Once activated, these NK cells rapidly elicit functions that can not only control HIV but may also prevent infection of new target cells.
6. Alloreactive NK Cells in Protection from Sexual HIV Transmission
Haploidentical hematopoietic stem cell transplants (HSCT) are used to treat acute myeloid leukemia and acute lymphoid leukemia. T cells are depleted from haploidentical HSCTs to reduce graft versus host disease mediated by T cells activated by allogeneic cells. NK cells in these haploidentical HSCTs mediate a graft versus leukemia effect if alloreactive educated NK cells respond to absence of ligands for inhibitory KIR on leukemia cells, leading to a reduced risk of leukemia relapse [95,96,97]. This finding was the basis for investigating whether alloreactive NK cells had a protective effect in the context of sexual HIV transmission. The effect needed to be investigated in sexual partners where the KIR/HLA combinations of both partners was available.
While still debated, studies infecting rhesus macaques with simian immunodeficiency virus favor a model in which simian immunodeficiency virus infected CD4 cells rather than free virions are the predominant mechanism for transmission [98,99,100,101]. By analogy, new HIV infections arising from sexual exposures occur, at least in part, from HIV-infected cells present in genital secretions [101]. Thus, for an immune response to be effective at preventing infection, it needs to be able to target incoming HIV-infected cells early in infection before resting CD4 cell HIV reservoirs are established [15,16]. NK cells can target HIV-infected CD4 cells and are present as resident cells throughout the vaginal, uterine, and gut mucosa [102]. NK cells are rapidly activated by allogeneic HIV-infected CD4 cells. The HIV Nef and Vpu driven downmodulation of HLA-A, -B, and -C antigens from the surface of HIV-infected cells can further contribute the activation of the recipient’s NK cell and the susceptibility of incoming allogenic cells to NK cell-mediated control [23,25,69,103,104,105].
Several reports showed that HLA mismatches between transmission pairs decreased the risk of perinatal and sexual HIV transmission [106,107,108,109,110]. KIR/HLA genetic variation, particularly combinations having an activating KIR/HLA profile such as activating KIRs, a group B KIR haplotype, or inhibitory KIRs in the absence of their ligands were associated with protection from infection in HESN individuals [71,74,78,79,80,91,111]. Jennes et al. investigated the role of NK cells in HIV resistance in the seronegative partners of serodiscordant couples and susceptibility to HIV in seroconcordant couples [105]. They hypothesized that, in certain contexts, NK cells were activated by allogeneic HIV-infected cells to kill infected cells originating from their sexual partners [105]. They found that HIV transmission in heterosexual couples depended on the presence of NK cell educating KIR/HLA combinations in the recipient partner. If the transmitting partner expressed MHC class I allotypes that were not recognized by the recipient partner’s educated NK cells (i.e., were mismatched) they were less likely to seroconvert than if the transmitting partner expressed MHC class I antigens recognized by the recipient partner’s NK cells (i.e., were matched) (Figure 3).
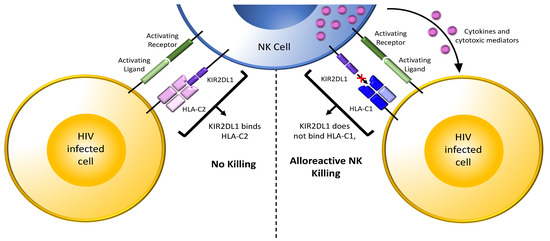
Figure 3.
NK cell responses to allogenic cells. Shown in the top middle is an NK cell from an HIV uninfected person expressing the inhibitory receptor/ligand combination KIR2DL1/HLA-C2. The interaction of KIR2DL1 with HLA-C2 during development educates these NK cells. When educated KIR2DL1+ NK cells in a recipient partner encounter incoming HIV-infected CD4 cells from a transmitting partner who is HLA-C2+ (bottom left-hand cell, matched combination), the interaction of HLA-C2 on infected cells with the KIR2DL1 inhibitory receptor on recipient partner NK cells inhibits NK cell activation despite HIV-infected cells having ligands for activating receptors on NK cells. In this scenario, cytolysis of the HIV-infected cells is inhibited, increasing the chance of HIV transmission. On the right, educated KIR2DL1+ NK cells in a recipient partner encounter incoming HIV-infected CD4 cells from a transmitting partner who is HLA-C1+ (bottom right-hand cell, unmatched combination). Absence of inhibitory signaling through KIR2DL1 together with the presence of activating signals from activating receptors interacting with their ligands on HIV-infected cells activates NK cells. Activated NK cells release cytokines and cytotoxic mediators, which suppress and/or kill incoming HIV-infected cells and reduce the chance of HIV transmission. Light green bar = ligand for an activating receptor; dark green bar on the NK cell = activating NK cell receptor; purple 2 domain structure on the NK cell = inhibitory NK cell receptor KIR2DL2; light pink 4 domain structure on the HIV infected cell (left) = HLA-C2, a ligand for inhibitory receptor KIR2DL1; blue 4 domain structure on the HIV infected cell (right) = HLA-C1, unable to bind inhibitory receptor KIR2DL1; purple dots = cytokines and cytotoxic mediators; “+” symbol between KIR2DL1 and HLA-C1 on the right indicated the absence of an interaction between this receptor and ligand pair.
Presumably, educated NK cells in the recipient partner were more likely to be activated by incoming allogeneic HIV-infected cells to reduce the risk of HIV transmission in the mismatched but not in the matched situation [105]. This idea was tested in vitro by co-culturing NK cells with allogeneic HIV-infected CD4 cells. These experiments confirmed that NK cells killed a higher frequency of missing self than matched HIV-infected cells. Later experiments further refined these observations by showing that both the frequency of lysed HIV-infected CD4 cells and the frequency and intensity of CD107a externalization by NK cells activated by allogeneic HIV-infected CD4 cells were higher in mismatched than in matched combinations [112]. NK cells from subjects carrying haplotype B KIR genotypes with activating KIRs contributed to greater levels of NK cell activation than when no activating KIRs were present [112]. HIV-infected cells also upregulate several ligands for activating NK cell receptors, which could further contribute to activating NK cell responses to HIV-infected cells [21]. MHC class I downmodulation by HIV Nef and Vpu may further contribute to the vulnerability of the incoming infected CD4 cells from transmitting partners who are PLWH to recipient NK cell responses. Together, these findings confirm previous observations that activating NK cell genotypes and inhibitory KIRs in the absence of their HLA ligands were associated with HIV resistance and suggest mechanisms that may underlie these observations [71,78,80,111,113].
It is notable that HIV serodiscordant couples were more likely to be composed of an uninfected recipient partner with an NK cell educating KIR/HLA combination such as KIR2DL1+HLA-C2 and a PLWH partner who was homozygous for C1 than seroconcordant couples (Figure 3) [105]. The contribution of KIR2DL+HLA-C KIR/HLA combinations to protection from HIV infection by the heterosexual transmission route appears to be greater than that of KIR3DL1/S1+Bw4 combinations [112]. The implication of KIR2DL+HLA-C combinations in protection from sexual HIV transmission complements the observation made for KIR3DL1+HLA-B*57 and KIR3DS1 in protection from parenteral HIV infection in IDU HESN individuals. More work is needed to understand whether KIR/HLA combinations contribute differentially to protection from infection according to the route of HIV transmission.
7. Mechanisms of Protection from HIV Infection
NK cells dependent mechanisms of host resistance to HIV infections described above have focused on the role of activated NK cells. Others have also implicated NK cell activation in in protection from HIV infection. Tomescu et al., working with a cohort of IDU HESN persons, found persistently higher frequencies of activated NK cells and monocyte-derived dendritic cells among HESN individuals sharing needles than among low-risk non-sharing IDU or non-drug using controls [114]. The enhanced activation of these cell types in IDU HESN did not appear to be due to the injection drugs used but rather due to other factors related to drug use such as exposure to pathogens or allogeneic cells from injection partners. A proteomic analysis of NK cells from IDU HESN individuals and healthy control donors found that the protein S100A14 was upregulated in the former group. A100A14 is a member of the S100 family that are involved in many cellular processes. Although these proteins have been studied in the context of cancer, little is known about their role in the setting of NK cells and HIV. This molecule activated NK cells in a monocyte-dependent manner that required NK cell monocyte cell contact. S100A14 activated monocytes by signaling through toll-like receptor 4 to secrete TNF-α. These findings suggest that S100A14 is involved in NK cells monocyte crosstalk that promoted NK cell activation in IDU HESN [115].
It should be noted, however, that there is unlikely to be a unified protective mechanism for protection from HIV in HESN, as both immune quiescence [116,117] and immune activation [118,119,120,121,122] have been identified depending on the route of exposure. Studies conducted in a highly HIV-exposed commercial sex worker (CSW) cohort followed in Nairobi, Kenya identified immune quiescence as a mechanism for resistance to HIV infection. A subset of these CSWs remained uninfected despite 7 or more years of HIV exposure [123]. These resistant CSW HESNs are distinguished from their HIV-susceptible counterparts by maintaining an immune profile characterized by quiescence [116]. As HIV infects and replicates preferentially in activated CD4+ T cells, the ability of HIV-susceptible CD4+ cells at the portals of entry to remain quiescent despite HIV exposure may preclude HIV replication in these cells and reduce the risk of establishing a productive infection [116]. A role of NK cells in immune quiescence has not been described.
8. Adaptive NK Cells in Protection from HIV Infection
Human cytomegalovirus (HCMV) infection expands a population of NK cells with adaptive-like features [124]. These adaptive NK (adapNK) cells express the activating receptor, NKG2C, which belongs to the C-type lectin family [125]. The NKG2C activating receptor, like its inhibitory counterpart, NKG2A, is expressed as a heterodimer with CD94 [126]. The ligand for NKG2C and NKG2A is HLA-E, a nonclassical MHC class Ib antigen stabilized by peptides derived from leader sequence of classical MHC class I antigens, the nonclassical MHC Ib HLA-G antigen or epitopes from the HCMV-encoded viral protein UL40 [39,127,128]. The interaction of NKG2C with its ligand transmits signals that activate cells expressing this receptor [39,129]. AdapNK cells undergo DNA methylation-dependent epigenetic modifications, which distinguish them from conventional NK cells and influence their functionality [130,131].
Some individuals do not express cell surface NKG2C due to a homozygous deletion of approximately 16 kb that includes the nkg2c gene encoding NKG2C [132,133]. In several Caucasian populations, a Japanese cohort, and a Tanzanian cohort, the frequency of the NKG2C deletion allele is close to 20% with a homozygous deletion genotype frequency of approximately 4% [133,134,135,136]. Several studies have questioned whether NKG2C+ cells play a role in protection from HIV infection or in rate of HIV disease progression in PLWH. Thomas et al. compared the distribution of NKG2C genotypes (NKG2C+/+, NKG2C+/− and NKG2C−/−) in 433 PLWH and 280 uninfected individuals with no history of HIV exposure. They reported that that carriage of at least one NKG2C- variant (i.e., the NKG2C+/− and NKG2C−/− genotypes) was associated with a higher risk of HIV infection [137]. In a separate study comparing 434 PLWH and 157 HESN persons there were no between-group differences for the frequency of either the NKG2C+/+ or the combined NKG2C+/− and NKG2C−/− genotypes [138]. However, the frequency of the NKG2C−/− genotype was higher among PLWH than HESN individuals. None of the 157 HESN subjects tested carried the NKG2C−/− genotype, which was present in 11 of 434 (2.53%) PLWH. Of note, the distribution of NKG2C genotypes did not differ in the PLWH and HESN sub-populations who were sexually exposed to HIV, while the NKG2C−/− genotype was more frequent in parenterally exposed PLWH than in HESN individuals. These findings suggest that the NKG2C−/− genotype is associated with a higher risk of HIV infection by parenteral, but not by sexual exposure [138].
In Thomas at al., the control population was not HIV-exposed and thus was at a low risk for HIV infection, while in the study by Alsulami et al. the HIV seronegative population were HESN persons at high risk for HIV exposure and more likely to have some level of resistance to HIV infection. Thus, the inclusion of HESN participants allowed for a more direct exploration of whether NKG2C genotypes were associated with HIV susceptibility.
What accounts for the frequency of the NKG2C−/− genotype not differing significantly between sexually exposed PLWH and HESN persons is unclear. To better understand this phenomenon, more information is needed regarding factors that influence the per act risk of HIV transmission by different routes and the NK cell, and specifically the NKG2C+ NK cell, landscape at mucosal portals of HIV entry. Additionally, factors such as treatment status and HCMV serostatus need to be accounted for.
9. Can Vaccines Induce NK Cell Responses That Prevent HIV Infection?
The RV144 HIV vaccine trial induced modest, though significant, protection against HIV infection [139]. Correlates of protection analyses found that vaccine-induced antibodies able to bind HIV Envelope structures and support NK cell-mediated antibody-dependent cellular cytotoxicity (ADCC) were associated with protection from HIV infection [140]. A vaccine regimen similar to the one used in the RV144 vaccine trial, designed for use in South Africa, where HIV clade C predominates, failed to show efficacy in preventing HIV infection [141]. This was despite it inducing immune responses analogous to those seen in the RV144 trial. Nevertheless, the RV144 trial offers an example of a role for NK cells in protection from HIV infection. Presumably, the HIV Envelope-specific IgG antibodies induced by this vaccine were able to bridge HIV-infected cells expressing cell surface HIV Envelope with CD16 on NK cells. Engagement and crosslinking of CD16 would have resulted in NK cell activation and the lysis of incoming HIV-infected cells by ADCC. This finding provides an impetus for investigating strategies that can improve the efficacy of vaccine-induced immune responses that protect from infection.
Early in an immune response, NK cells secrete cytokines and chemokines such as IFN-γ, TNF-α, CCL3, CCL4, CCL5, and granulocyte-macrophage colony-stimulating factor [142]. These secreted factors contribute to recruiting and activating antigen-presenting cells such as dendritic cells, which in turn collaborate with T and B cells to induce adaptive immunity [143]. IFN-γ derived from NK cells promotes type 1 T helper cell differentiation and stimulates isotype class switching in B cells [144,145]. Designing vaccines that harness NK cell activity has the potential to improve the induction of adaptive immune responses able to collaborate with NK cells to prevent HIV infection [146]. The incorporation of novel adjuvants into vaccine formulations may induce NK cell activities that increase adaptive immune responses [147,148,149,150].
One goal would be to design vaccines that activate NK cells to promote B cells to produce antibodies able to bind to Envelope on HIV-infected cells that are ADCC competent. The presence of such antibodies able to opsonize incoming HIV-infected cells would have the potential to lead to their lysis through ADCC.
Human cytomegalovirus (HCMV) infection drives the expansion of adaptive-like NK cells expressing NKG2C and/or that are negative for FcRγ and other signaling molecules. These adaptive-like NK cells have epigenetic changes that confer antibody dependent effector functions such as ADCC activity that is superior to that seen in conventional NK cells [124,130,131,151,152]. Vaccine strategies aimed at inducing NKG2C+ NK cells in the absence of CMV infection are being investigated [153].
However, NK cells can also negatively regulate immune responses through their cytolytic activity. NK cell-mediated cytolysis can reduce vaccine antigen persistence, eradicate responding T cells, suppress the establishment of adaptive memory, and block the evolution of antibody responses [143,154,155]. The dichotomous activity of NK cells adds complexity to targeting these cells through vaccination. Future investigations will need to consider this issue when designing vaccines targeting NK cells.
10. Concluding Remarks and Future Directions
HESN individuals can be found among IDUs, HIV serodiscordant couples, CSWs, and perinatally exposed infants. Studying HESN persons may provide clues to mechanisms that prevent HIV infection. There is evidence of a role of NK cells in protection from HIV infection through parenteral and sexual routes of exposure.
In terms of parenteral exposure, information on the transmitting partners is rarely available. Thus, epidemiological studies have relied on whether certain KIR/HLA combinations are associated with protection from HIV infection. Those that are, tend to be combinations that favor NK cell education for potent responses to autologous HIV-infected cells in the case of the KIR3DL1+HLA-B*57 combination and NK cell activation through ligation of the KIR3DS1 activating receptor on NK cells by ligands on HIV-infected cells as in the case of KIR3DS1+HLA-F combinations. In both these cases, NK cells from carriers of both these KIR/HLA combinations are induced to secrete higher levels of cytokines and chemokines and externalize higher levels of CD107a when co-cultured with autologous HIV-infected cells that all have anti-HIV activity.
In HIV serodiscordant and seroconcordant couples, the identity of both the recipient and transmitting partners is often available. By typing recipient partners for KIR and HLA and transmitting partners for HLA, we have learned that HIV discordance is more likely when the recipient partner has educated NK cells that cannot recognize HLA on the transmitting partner’s cells. In this scenario, recipient NK cells can respond to incoming allogeneic HIV-infected cells in a manner that reduces the risk of HIV infection. Lack of inhibition together with the induction of ligands for activating NK receptors on infected cells can favor NK cell activation in HESN recipient partners in this unmatched allogeneic context [105,112].
Several questions remain to be addressed. The role of the route of HIV exposure in determining whether KIR3DL1/S1 and HLA-Bw4/HLA-F combinations are more potent in parenteral exposures while KIR2DL/HLA-C combinations are more effective at preventing HIV infection by sexual routes of exposure is unclear. We still have much to learn regarding the anatomy of NK (or ILC1) cell distribution at portals of HIV entry through sexual transmission. The contribution of immune quiescence versus NK cell activation to protection from HIV infection require further investigation.
NK cells not only respond directly to autologous and allogeneic cells’ missing ligands for inhibitory receptors but also mediate potent ADCC. To take advantage of this type of NK cell function, antibodies are required. It may be possible to design novel next-generation vaccines that harness the potential of NK cells to induce B cells to secrete antibodies that recognize the Envelope of HIV-infected cells rather than uninfected bystander cells [156,157,158]. The presence of such antibodies able to opsonize incoming HIV-infected cells would be effective targets for NK cell-mediated ADCC activity occurring soon enough after exposure to prevent the establishment of infection. Novel vaccine adjuvants may help induce NK cells to promote protective rather than detrimental NK cell functions. Inducing adaptive NK cells through vaccination may generate NK cells with superior ADCC activity that plays a role in preventing HIV infection.
Funding
Canadian Institutes of Health Research (CIHR) grants # 148491 and # 142494. Fonds de Recherche du Québec-Santé (FRQ-S) AIDS and Infectious Diseases Network.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Meyers, A.F.; Fowke, K.R. International symposium on natural immunity to HIV: A gathering of the HIV-exposed seronegative clan. J. Infect. Dis. 2010, 202 (Suppl. S3), S327–S328. [Google Scholar] [CrossRef]
- Beretta, A.; Furci, L.; Burastero, S.; Cosma, A.; Dinelli, M.E.; Lopalco, L.; DeSantis, C.; Tambussi, G.; Carrow, E.; Sabbatani, S.; et al. HIV-1-specific immunity in persistently seronegative individuals at high risk for HIV infection. Immunol. Lett. 1996, 51, 39–43. [Google Scholar] [CrossRef]
- Clerici, M.; Giorgi, J.V.; Chou, C.C.; Gudeman, V.K.; Zack, J.A.; Gupta, P.; Ho, H.N.; Nishanian, P.G.; Berzofsky, J.A.; Shearer, G.M. Cell-mediated immune response to human immunodeficiency virus (HIV) type 1 in seronegative homosexual men with recent sexual exposure to HIV-1. J. Infect. Dis. 1992, 165, 1012–1019. [Google Scholar] [CrossRef] [PubMed]
- Clerici, M.; Levin, J.M.; Kessler, H.A.; Harris, A.; Berzofsky, J.A.; Landay, A.L.; Shearer, G.M. HIV-specific T-helper activity in seronegative health care workers exposed to contaminated blood. JAMA 1994, 271, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Mazzoli, S.; Trabattoni, D.; Lo, C.S.; Piconi, S.; Ble, C.; Meacci, F.; Ruzzante, S.; Salvi, A.; Semplici, F.; Longhi, R.; et al. HIV-specific mucosal and cellular immunity in HIV-seronegative partners of HIV-seropositive individuals. Nat. Med. 1997, 3, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.A.; Sullivan, J.; Berzofsky, J.A.; Clerici, M.; Kessler, H.A.; Landay, A.L.; Shearer, G.M. ENV-specific cytotoxic T lymphocyte responses in HIV seronegative health care workers occupationally exposed to HIV-contaminated body fluids. J. Clin. Investig. 1995, 96, 867–876. [Google Scholar] [CrossRef]
- Ranki, A.; Mattinen, S.; Yarchoan, R.; Broder, S.; Ghrayeb, J.; Lahdevirta, J.; Krohn, K. T-cell response towards HIV in infected individuals with and without zidovudine therapy, and in HIV-exposed sexual partners. AIDS 1989, 3, 63–69. [Google Scholar] [CrossRef]
- Kaul, R.; Rowland-Jones, S.L.; Kimani, J.; Dong, T.; Yang, H.B.; Kiama, P.; Rostron, T.; Njagi, E.; Bwayo, J.J.; MacDonald, K.S.; et al. Late seroconversion in HIV-resistant Nairobi prostitutes despite pre-existing HIV-specific CD8+ responses. J. Clin. Investig. 2001, 107, 341–349. [Google Scholar] [CrossRef]
- Kaul, R.; MacDonald, K.S.; Nagelkerke, N.J.; Kimani, J.; Fowke, K.; Ball, T.B.; Luo, M.; Kariri, A.; Jaoko, W.; Moses, S.; et al. HIV viral set point and host immune control in individuals with HIV-specific CD8+ T-cell responses prior to HIV acquisition. AIDS 2010, 24, 1449–1454. [Google Scholar] [CrossRef]
- Liu, Y.; Woodward, A.; Zhu, H.; Andrus, T.; McNevin, J.; Lee, J.; Mullins, J.I.; Corey, L.; McElrath, M.J.; Zhu, T. Preinfection human immunodeficiency virus (HIV)-specific cytotoxic T lymphocytes failed to prevent HIV type 1 infection from strains genetically unrelated to viruses in long-term exposed partners. J. Virol. 2009, 83, 10821–10829. [Google Scholar] [CrossRef][Green Version]
- Alimonti, J.B.; Kimani, J.; Matu, L.; Wachihi, C.; Kaul, R.; Plummer, F.A.; Fowke, K.R. Characterization of CD8 T-cell responses in HIV-1-exposed seronegative commercial sex workers from Nairobi, Kenya. Immunol. Cell Biol. 2006, 84, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Kaul, R.; Rowland-Jones, S.L.; Kimani, J.; Fowke, K.; Dong, T.; Kiama, P.; Rutherford, J.; Njagi, E.; Mwangi, F.; Rostron, T.; et al. New insights into HIV-1 specific cytotoxic T-lymphocyte responses in exposed, persistently seronegative Kenyan sex workers. Immunol. Lett. 2001, 79, 3–13. [Google Scholar] [CrossRef]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; del Rio, C.; et al. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef]
- McElrath, M.J.; De Rosa, S.C.; Moodie, Z.; Dubey, S.; Kierstead, L.; Janes, H.; Defawe, O.D.; Carter, D.K.; Hural, J.; Akondy, R.; et al. HIV-1 vaccine-induced immunity in the test-of-concept Step Study: A case-cohort analysis. Lancet 2008, 372, 1894–1905. [Google Scholar] [CrossRef]
- Whitney, J.B.; Hill, A.L.; Sanisetty, S.; Penaloza-MacMaster, P.; Liu, J.; Shetty, M.; Parenteau, L.; Cabral, C.; Shields, J.; Blackmore, S.; et al. Rapid seeding of the viral reservoir prior to SIV viraemia in rhesus monkeys. Nature 2014, 512, 74–77. [Google Scholar] [CrossRef]
- Haase, A.T. Early events in sexual transmission of HIV and SIV and opportunities for interventions. Annu. Rev. Med. 2011, 62, 127–139. [Google Scholar] [CrossRef]
- Tomescu, C.; Abdulhaqq, S.; Montaner, L.J. Evidence for the innate immune response as a correlate of protection in human immunodeficiency virus (HIV)-1 highly exposed seronegative subjects (HESN). Clin. Exp. Immunol 2011, 164, 158–169. [Google Scholar] [CrossRef]
- Alter, G.; Teigen, N.; Ahern, R.; Streeck, H.; Meier, A.; Rosenberg, E.S.; Altfeld, M. Evolution of innate and adaptive effector cell functions during acute HIV-1 infection. J. Infect. Dis 2007, 195, 1452–1460. [Google Scholar] [CrossRef]
- Trinchieri, G. Biology of natural killer cells. Adv. Immunol. 1989, 47, 187–376. [Google Scholar]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef]
- Tremblay-McLean, A.; Bruneau, J.; Lebouche, B.; Lisovsky, I.; Song, R.; Bernard, N.F. Expression Profiles of Ligands for Activating Natural Killer Cell Receptors on HIV Infected and Uninfected CD4(+) T Cells. Viruses 2017, 9, 295. [Google Scholar] [CrossRef] [PubMed]
- Apps, R.; Del Prete, G.Q.; Chatterjee, P.; Lara, A.; Brumme, Z.L.; Brockman, M.A.; Neil, S.; Pickering, S.; Schneider, D.K.; Piechocka-Trocha, A.; et al. HIV-1 Vpu Mediates HLA-C Downregulation. Cell Host. Microbe 2016, 19, 686–695. [Google Scholar] [CrossRef]
- Cohen, G.B.; Gandhi, R.T.; Davis, D.M.; Mandelboim, O.; Chen, B.K.; Strominger, J.L.; Baltimore, D. The selective downregulation of class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells. Immunity 1999, 10, 661–671. [Google Scholar] [CrossRef]
- Kiani, Z.; Bruneau, J.; Geraghty, D.E.; Bernard, N.F. HLA-F on Autologous HIV-Infected Cells Activates Primary NK Cells Expressing the Activating Killer Immunoglobulin-Like Receptor KIR3DS1. J. Virol. 2019, 93, e00933-19. [Google Scholar] [CrossRef]
- Bonaparte, M.I.; Barker, E. Killing of human immunodeficiency virus-infected primary T-cell blasts by autologous natural killer cells is dependent on the ability of the virus to alter the expression of major histocompatibility complex class I molecules. Blood 2004, 104, 2087–2094. [Google Scholar] [CrossRef] [PubMed]
- Biron, C.A.; Nguyen, K.B.; Pien, G.C.; Cousens, L.P.; Salazar-Mather, T.P. Natural killer cells in antiviral defense: Function and regulation by innate cytokines. Annu. Rev. Immunol. 1999, 17, 189–220. [Google Scholar] [CrossRef]
- Oliva, A.; Kinter, A.L.; Vaccarezza, M.; Rubbert, A.; Catanzaro, A.; Moir, S.; Monaco, J.; Ehler, L.; Mizell, S.; Jackson, R.; et al. Natural killer cells from human immunodeficiency virus (HIV)-infected individuals are an important source of CC-chemokines and suppress HIV-1 entry and replication in vitro. J. Clin. Investig. 1998, 102, 223–231. [Google Scholar] [CrossRef]
- Song, R.; Lisovsky, I.; Lebouche, B.; Routy, J.P.; Bruneau, J.; Bernard, N.F. HIV Protective KIR3DL1/S1-HLA-B Genotypes Influence NK Cell-Mediated Inhibition of HIV Replication in Autologous CD4 Targets. PLoS Pathog. 2014, 10, e1003867. [Google Scholar] [CrossRef]
- Long, E.O.; Kim, H.S.; Liu, D.; Peterson, M.E.; Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 2013, 31, 227–258. [Google Scholar] [CrossRef]
- Long, E.O. Regulation of immune responses through inhibitory receptors. Annu. Rev. Immunol. 1999, 17, 875–904. [Google Scholar] [CrossRef]
- Watzl, C.; Stebbins, C.C.; Long, E.O. NK cell inhibitory receptors prevent tyrosine phosphorylation of the activation receptor 2B4 (CD244). J. Immunol. 2000, 165, 3545–3548. [Google Scholar] [CrossRef] [PubMed]
- Anfossi, N.; Andre, P.; Guia, S.; Falk, C.S.; Roetynck, S.; Stewart, C.A.; Breso, V.; Frassati, C.; Reviron, D.; Middleton, D.; et al. Human NK cell education by inhibitory receptors for MHC class I. Immunity 2006, 25, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Joncker, N.T.; Fernandez, N.C.; Treiner, E.; Vivier, E.; Raulet, D.H. NK cell responsiveness is tuned commensurate with the number of inhibitory receptors for self-MHC class I: The rheostat model. J. Immunol. 2009, 182, 4572–4580. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Poursine-Laurent, J.; Truscott, S.M.; Lybarger, L.; Song, Y.J.; Yang, L.; French, A.R.; Sunwoo, J.B.; Lemieux, S.; Hansen, T.H.; et al. Licensing of natural killer cells by host major histocompatibility complex class I molecules. Nature 2005, 436, 709–713. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Mulrooney, T.J.; Le Luduec, J.B.; Barker, E.; Hsu, K.C. KIR3DL1 and HLA-B Density and Binding Calibrate NK Education and Response to HIV. J. Immunol. 2016, 196, 3398–3410. [Google Scholar] [CrossRef]
- Raulet, D.H.; Vance, R.E. Self-tolerance of natural killer cells. Nat. Rev. Immunol. 2006, 6, 520–531. [Google Scholar] [CrossRef]
- Joncker, N.T.; Raulet, D.H. Regulation of NK cell responsiveness to achieve self-tolerance and maximal responses to diseased target cells. Immunol. Rev. 2008, 224, 85–97. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Hsu, K.C. Natural Killer Cell Education and the Response to Infection and Cancer Therapy: Stay Tuned. Trends Immunol. 2018, 39, 222–239. [Google Scholar] [CrossRef]
- Braud, V.M.; Allan, D.S.; O’Callaghan, C.A.; Soderstrom, K.; D’Andrea, A.; Ogg, G.S.; Lazetic, S.; Young, N.T.; Bell, J.I.; Phillips, J.H.; et al. HLA-E binds to natural killer cell receptors CD94/NKG2A, B and C. Nature 1998, 391, 795–799. [Google Scholar] [CrossRef]
- Trowsdale, J.; Barten, R.; Haude, A.; Stewart, C.A.; Beck, S.; Wilson, M.J. The genomic context of natural killer receptor extended gene families. Immunol. Rev. 2001, 181, 20–38. [Google Scholar] [CrossRef]
- Raulet, D.H.; Vance, R.E.; McMahon, C.W. Regulation of the natural killer cell receptor repertoire. Annu. Rev. Immunol. 2001, 19, 291–330. [Google Scholar] [CrossRef]
- Jiang, W.; Johnson, C.; Jayaraman, J.; Simecek, N.; Noble, J.; Moffatt, M.F.; Cookson, W.O.; Trowsdale, J.; Traherne, J.A. Copy number variation leads to considerable diversity for B but not A haplotypes of the human KIR genes encoding NK cell receptors. Genome Res. 2012, 22, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Sunwoo, J.B.; Yang, L.; Choi, T.; Song, Y.J.; French, A.R.; Vlahiotis, A.; Piccirillo, J.F.; Cella, M.; Colonna, M.; et al. HLA alleles determine differences in human natural killer cell responsiveness and potency. Proc. Natl. Acad. Sci. USA 2008, 105, 3053–3058. [Google Scholar] [CrossRef] [PubMed]
- Moesta, A.K.; Norman, P.J.; Yawata, M.; Yawata, N.; Gleimer, M.; Parham, P. Synergistic polymorphism at two positions distal to the ligand-binding site makes KIR2DL2 a stronger receptor for HLA-C than KIR2DL3. J. Immunol. 2008, 180, 3969–3979. [Google Scholar] [CrossRef] [PubMed]
- Carr, W.H.; Pando, M.J.; Parham, P. KIR3DL1 polymorphisms that affect NK cell inhibition by HLA-Bw4 ligand. J. Immunol. 2005, 175, 5222–5229. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Heller, G.; Chewning, J.; Kim, S.; Yokoyama, W.M.; Hsu, K.C. Hierarchy of the human natural killer cell response is determined by class and quantity of inhibitory receptors for self-HLA-B and HLA-C ligands. J. Immunol. 2007, 179, 5977–5989. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, N.C.; Lozier, A.; Flament, C.; Ricciardi-Castagnoli, P.; Bellet, D.; Suter, M.; Perricaudet, M.; Tursz, T.; Maraskovsky, E.; Zitvogel, L. Dendritic cells directly trigger NK cell functions: Cross-talk relevant in innate anti-tumor immune responses in vivo. Nat. Med. 1999, 5, 405–411. [Google Scholar] [CrossRef]
- Roe, D.; Vierra-Green, C.; Pyo, C.W.; Geraghty, D.E.; Spellman, S.R.; Maiers, M.; Kuang, R. A Detailed View of KIR Haplotype Structures and Gene Families as Provided by a New Motif-Based Multiple Sequence Alignment. Front. Immunol. 2020, 11, 585731. [Google Scholar] [CrossRef]
- Marsh, S.G.; Parham, P.; Dupont, B.; Geraghty, D.E.; Trowsdale, J.; Middleton, D.; Vilches, C.; Carrington, M.; Witt, C.; Guethlein, L.A.; et al. Killer-cell immunoglobulin-like receptor (KIR) nomenclature report, 2002. Hum. Immunol. 2003, 64, 648–654. [Google Scholar] [CrossRef]
- Carrington, M.; Norman, P. The KIR Gene Cluster; National Center for Biotechnology Information US: Bethesda MD, USA, 2003; pp. 1–165. [Google Scholar]
- Cella, M.; Longo, A.; Ferrara, G.B.; Strominger, J.L.; Colonna, M. NK3-specific natural killer cells are selectively inhibited by Bw4-positive HLA alleles with isoleucine 80. J. Exp. Med. 1994, 180, 1235–1242. [Google Scholar] [CrossRef]
- Gumperz, J.E.; Litwin, V.; Phillips, J.H.; Lanier, L.L.; Parham, P. The Bw4 public epitope of HLA-B molecules confers reactivity with natural killer cell clones that express NKB1, a putative HLA receptor. J. Exp. Med. 1995, 181, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, D.H.; Allen, T.M.; Vogel, T.U.; Jing, P.; DeSouza, I.P.; Dodds, E.; Dunphy, E.J.; Melsaether, C.; Mothe, B.; Yamamoto, H.; et al. Acute phase cytotoxic T lymphocyte escape is a hallmark of simian immunodeficiency virus infection. Nat. Med. 2002, 8, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Trundley, A.; Frebel, H.; Jones, D.; Chang, C.; Trowsdale, J. Allelic expression patterns of KIR3DS1 and 3DL1 using the Z27 and DX9 antibodies. Eur. J. Immunol. 2007, 37, 780–787. [Google Scholar] [CrossRef]
- Colonna, M.; Borsellino, G.; Falco, M.; Ferrara, G.B.; Strominger, J.L. HLA-C is the inhibitory ligand that determines dominant resistance to lysis by NK1- and NK2-specific natural killer cells. Proc. Natl. Acad. Sci. USA 1993, 90, 12000–12004. [Google Scholar] [CrossRef]
- Winter, C.C.; Gumperz, J.E.; Parham, P.; Long, E.O.; Wagtmann, N. Direct binding and functional transfer of NK cell inhibitory receptors reveal novel patterns of HLA-C allotype recognition. J. Immunol. 1998, 161, 571–577. [Google Scholar]
- Moesta, A.K.; Graef, T.; Abi-Rached, L.; Older Aguilar, A.M.; Guethlein, L.A.; Parham, P. Humans differ from other hominids in lacking an activating NK cell receptor that recognizes the C1 epitope of MHC class I. J. Immunol. 2010, 185, 4233–4237. [Google Scholar] [CrossRef]
- Hsu, K.C.; Chida, S.; Geraghty, D.E.; Dupont, B. The killer cell immunoglobulin-like receptor (KIR) genomic region: Gene-order, haplotypes and allelic polymorphism. Immunol. Rev. 2002, 190, 40–52. [Google Scholar] [CrossRef]
- Pyo, C.W.; Guethlein, L.A.; Vu, Q.; Wang, R.; Abi-Rached, L.; Norman, P.J.; Marsh, S.G.; Miller, J.S.; Parham, P.; Geraghty, D.E. Different patterns of evolution in the centromeric and telomeric regions of group A and B haplotypes of the human killer cell Ig-like receptor locus. PLoS ONE 2010, 5, e15115. [Google Scholar] [CrossRef]
- Uhrberg, M.; Valiante, N.M.; Shum, B.P.; Shilling, H.G.; Lienert-Weidenbach, K.; Corliss, B.; Tyan, D.; Lanier, L.L.; Parham, P. Human diversity in killer cell inhibitory receptor genes. Immunity 1997, 7, 753–763. [Google Scholar] [CrossRef]
- Pyo, C.W.; Wang, R.; Vu, Q.; Cereb, N.; Yang, S.Y.; Duh, F.M.; Wolinsky, S.; Martin, M.P.; Carrington, M.; Geraghty, D.E. Recombinant structures expand and contract inter and intragenic diversification at the KIR locus. BMC Genom. 2013, 14, 89. [Google Scholar] [CrossRef]
- Maxwell, L.D.; Wallace, A.; Middleton, D.; Curran, M.D. A common KIR2DS4 deletion variant in the human that predicts a soluble KIR molecule analogous to the KIR1D molecule observed in the rhesus monkey. Tissue Antigens 2002, 60, 254–258. [Google Scholar] [CrossRef]
- Middleton, D.; Gonzalez, A.; Gilmore, P.M. Studies on the expression of the deleted KIR2DS4*003 gene product and distribution of KIR2DS4 deleted and nondeleted versions in different populations. Hum. Immunol. 2007, 68, 128–134. [Google Scholar] [CrossRef]
- Wilson, M.J.; Torkar, M.; Haude, A.; Milne, S.; Jones, T.; Sheer, D.; Beck, S.; Trowsdale, J. Plasticity in the organization and sequences of human KIR/ILT gene families. Proc. Natl. Acad. Sci. USA 2000, 97, 4778–4783. [Google Scholar] [CrossRef]
- Yawata, M.; Yawata, N.; McQueen, K.L.; Cheng, N.W.; Guethlein, L.A.; Rajalingam, R.; Shilling, H.G.; Parham, P. Predominance of group A KIR haplotypes in Japanese associated with diverse NK cell repertoires of KIR expression. Immunogenetics 2002, 54, 543–550. [Google Scholar] [CrossRef]
- Truong, L.X.; Luong, T.T.; Scott-Algara, D.; Versmisse, P.; David, A.; Perez-Bercoff, D.; Nguyen, N.V.; Tran, H.K.; Cao, C.T.; Fontanet, A.; et al. CD4 cell and CD8 cell-mediated resistance to HIV-1 infection in exposed uninfected intravascular drug users in Vietnam. AIDS 2003, 17, 1425–1434. [Google Scholar] [CrossRef]
- Martin, M.P.; Qi, Y.; Gao, X.; Yamada, E.; Martin, J.N.; Pereyra, F.; Colombo, S.; Brown, E.E.; Shupert, W.L.; Phair, J.; et al. Innate partnership of HLA-B and KIR3DL1 subtypes against HIV-1. Nat. Genet. 2007, 39, 733–740. [Google Scholar] [CrossRef]
- Martin, M.P.; Gao, X.; Lee, J.H.; Nelson, G.W.; Detels, R.; Goedert, J.J.; Buchbinder, S.; Hoots, K.; Vlahov, D.; Trowsdale, J.; et al. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat. Genet. 2002, 31, 429–434. [Google Scholar] [CrossRef]
- Kiani, Z.; Dupuy, F.P.; Bruneau, J.; Lebouche, B.; Retiere, C.; Geraghty, D.E.; Bernard, N.F. The Education of NK Cells Determines Their Responsiveness to Autologous HIV-Infected CD4 T Cells. J. Virol. 2019, 93, e01185-19. [Google Scholar] [CrossRef]
- Boulet, S.; Kleyman, M.; Kim, J.Y.; Kamya, P.; Sharafi, S.; Simic, N.; Bruneau, J.; Routy, J.P.; Tsoukas, C.M.; Bernard, N.F. A combined genotype of KIR3DL1 high expressing alleles and HLA-B*57 is associated with a reduced risk of HIV infection. AIDS 2008, 22, 1487–1491. [Google Scholar] [CrossRef]
- Boulet, S.; Sharafi, S.; Simic, N.; Bruneau, J.; Routy, J.P.; Tsoukas, C.M.; Bernard, N.F. Increased proportion of KIR3DS1 homozygotes in HIV-exposed uninfected individuals. AIDS 2008, 22, 595–599. [Google Scholar] [CrossRef]
- Bruneau, J.; Daniel, M.; Abrahamowicz, M.; Zang, G.; Lamothe, F.; Vincelette, J. Trends in human immunodeficiency virus incidence and risk behavior among injection drug users in montreal, Canada: A 16-year longitudinal study. Am. J. Epidemiol. 2011, 173, 1049–1058. [Google Scholar] [CrossRef]
- Mehraj, V.; Cox, J.; Lebouche, B.; Costiniuk, C.; Cao, W.; Li, T.; Ponte, R.; Thomas, R.; Szabo, J.; Baril, J.G.; et al. Socio-economic status and time trends associated with early ART initiation following primary HIV infection in Montreal, Canada: 1996 to 2015. J. Int. AIDS Soc. 2018, 21, e25034. [Google Scholar] [CrossRef]
- Tallon, B.J.; Bruneau, J.; Tsoukas, C.M.; Routy, J.P.; Kiani, Z.; Tan, X.; Bernard, N.F. Time to seroconversion in HIV-exposed subjects carrying protective versus non protective KIR3DS1/L1 and HLA-B genotypes. PLoS ONE 2014, 9, e110480. [Google Scholar] [CrossRef]
- Tremblay-McLean, A.; Coenraads, S.; Kiani, Z.; Dupuy, F.P.; Bernard, N.F. Expression of ligands for activating natural killer cell receptors on cell lines commonly used to assess natural killer cell function. BMC Immunol. 2019, 20, 8. [Google Scholar] [CrossRef]
- Boulet, S.; Song, R.; Kamya, P.; Bruneau, J.; Shoukry, N.H.; Tsoukas, C.M.; Bernard, N.F. HIV protective KIR3DL1 and HLA-B genotypes influence NK cell function following stimulation with HLA-devoid cells. J. Immunol. 2010, 184, 2057–2064. [Google Scholar] [CrossRef]
- Parsons, M.S.; Boulet, S.; Song, R.; Bruneau, J.; Shoukry, N.H.; Routy, J.P.; Tsoukas, C.M.; Bernard, N.F. Mind the gap: Lack of association between KIR3DL1*004/HLA-Bw4-induced natural killer cell function and protection from HIV infection. J. Infect. Dis. 2010, 202 (Suppl. S3), S356–S360. [Google Scholar] [CrossRef]
- Jackson, E.; Zhang, C.X.; Kiani, Z.; Lisovsky, I.; Tallon, B.; Del Corpo, A.; Gilbert, L.; Bruneau, J.; Thomas, R.; Cote, P.; et al. HIV exposed seronegative (HESN) compared to HIV infected individuals have higher frequencies of telomeric Killer Immunoglobulin-like Receptor (KIR) B motifs; Contribution of KIR B motif encoded genes to NK cell responsiveness. PLoS ONE 2017, 12, e0185160. [Google Scholar] [CrossRef]
- Ravet, S.; Scott-Algara, D.; Bonnet, E.; Tran, H.K.; Tran, T.; Nguyen, N.; Truong, L.X.; Theodorou, I.; Barre-Sinoussi, F.; Pancino, G.; et al. Distinctive NK-cell receptor repertoires sustain high-level constitutive NK-cell activation in HIV-exposed uninfected individuals. Blood 2007, 109, 4296–4305. [Google Scholar] [CrossRef]
- Guerini, F.R.; Lo, C.S.; Gori, A.; Bandera, A.; Mazzotta, F.; Uglietti, A.; Zanzottera, M.; Maserati, R.; Clerici, M. Under Representation of the Inhibitory KIR3DL1 Molecule and the KIR3DL1+/BW4+ Complex in HIV Exposed Seronegative Individuals. J. Infect. Dis. 2011, 203, 1235–1239. [Google Scholar] [CrossRef]
- Habegger de, S.A.; Sinchi, J.L.; Marinic, K.; Lopez, R.; Iliovich, E. KIR-HLA-A and B alleles of the Bw4 epitope against HIV infection in discordant heterosexual couples in Chaco Argentina. Immunology 2013, 140, 273–279. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Holzemer, A.; Martrus, G.; Chung, A.W.; Pacheco, Y.; Simoneau, C.R.; Rucevic, M.; Lamothe-Molina, P.A.; Pertel, T.; Kim, T.E.; et al. Open conformers of HLA-F are high-affinity ligands of the activating NK-cell receptor KIR3DS1. Nat. Immunol. 2016, 17, 1067–1074. [Google Scholar] [CrossRef]
- Gillespie, G.M.; Bashirova, A.; Dong, T.; McVicar, D.W.; Rowland-Jones, S.L.; Carrington, M. Lack of KIR3DS1 binding to MHC class I Bw4 tetramers in complex with CD8+ T cell epitopes. AIDS Res. Hum. Retrovir. 2007, 23, 451–455. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, G.M.; Vivian, J.P.; Gostick, E.; Pymm, P.; Lafont, B.A.; Price, D.A.; Rossjohn, J.; Brooks, A.G.; McVicar, D.W. Peptide-Dependent Recognition of HLA-B*57:01 by KIR3DS1. J. Virol. 2015, 89, 5213–5221. [Google Scholar] [CrossRef] [PubMed]
- Fauriat, C.; Ivarsson, M.A.; Ljunggren, H.G.; Malmberg, K.J.; Michaelsson, J. Education of human natural killer cells by activating killer cell immunoglobulin-like receptors. Blood 2010, 115, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.A.; Laugier-Anfossi, F.; Vely, F.; Saulquin, X.; Riedmuller, J.; Tisserant, A.; Gauthier, L.; Romagne, F.; Ferracci, G.; Arosa, F.A.; et al. Recognition of peptide-MHC class I complexes by activating killer immunoglobulin-like receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 13224–13229. [Google Scholar] [CrossRef] [PubMed]
- Kiani, Z.; Dupuy, F.P.; Bruneau, J.; Lebouche, B.; Zhang, C.X.; Jackson, E.; Lisovsky, I.; da Fonseca, S.; Geraghty, D.E.; Bernard, N.F. HLA-F on HLA-Null 721.221 Cells Activates Primary NK Cells Expressing the Activating Killer Ig-like Receptor KIR3DS1. J. Immunol. 2018, 201, 113–123. [Google Scholar] [CrossRef]
- Lee, N.; Ishitani, A.; Geraghty, D.E. HLA-F is a surface marker on activated lymphocytes. Eur. J. Immunol. 2010, 40, 2308–2318. [Google Scholar] [CrossRef]
- Burian, A.; Wang, K.L.; Finton, K.A.; Lee, N.; Ishitani, A.; Strong, R.K.; Geraghty, D.E. HLA-F and MHC-I Open Conformers Bind Natural Killer Cell Ig-Like Receptor KIR3DS1. PLoS ONE 2016, 11, e0163297. [Google Scholar] [CrossRef]
- Alter, G.; Martin, M.P.; Teigen, N.; Carr, W.H.; Suscovich, T.J.; Schneidewind, A.; Streeck, H.; Waring, M.; Meier, A.; Brander, C.; et al. Differential natural killer cell-mediated inhibition of HIV-1 replication based on distinct KIR/HLA subtypes. J. Exp. Med. 2007, 204, 3027–3036. [Google Scholar] [CrossRef]
- Merino, A.; Malhotra, R.; Morton, M.; Mulenga, J.; Allen, S.; Hunter, E.; Tang, J.; Kaslow, R.A. Impact of a functional KIR2DS4 allele on heterosexual HIV-1 transmission among discordant Zambian couples. J. Infect. Dis. 2011, 203, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Merino, A.M.; Dugast, A.S.; Wilson, C.M.; Goepfert, P.A.; Alter, G.; Kaslow, R.A.; Tang, J. KIR2DS4 promotes HIV-1 pathogenesis: New evidence from analyses of immunogenetic data and natural killer cell function. PLoS ONE 2014, 9, e99353. [Google Scholar] [CrossRef]
- Olvera, A.; Perez-Alvarez, S.; Ibarrondo, J.; Ganoza, C.; Lama, J.R.; Lucchetti, A.; Cate, S.; Hildebrand, W.; Bernard, N.; Gomez, L.; et al. The HLA-C*04: 01/KIR2DS4 gene combination and human leukocyte antigen alleles with high population frequency drive rate of HIV disease progression. AIDS 2015, 29, 507–517. [Google Scholar] [CrossRef]
- Graef, T.; Moesta, A.K.; Norman, P.J.; Abi-Rached, L.; Vago, L.; Older Aguilar, A.M.; Gleimer, M.; Hammond, J.A.; Guethlein, L.A.; Bushnell, D.A.; et al. KIR2DS4 is a product of gene conversion with KIR3DL2 that introduced specificity for HLA-A*11 while diminishing avidity for HLA-C. J. Exp. Med. 2009, 206, 2557–2572. [Google Scholar] [CrossRef] [PubMed]
- Aversa, F.; Terenzi, A.; Tabilio, A.; Falzetti, F.; Carotti, A.; Ballanti, S.; Felicini, R.; Falcinelli, F.; Velardi, A.; Ruggeri, L.; et al. Full haplotype-mismatched hematopoietic stem-cell transplantation: A phase II study in patients with acute leukemia at high risk of relapse. J. Clin. Oncol. 2005, 23, 3447–3454. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, F.; Pende, D.; Mingari, M.C.; Bertaina, A.; Falco, M.; Moretta, A.; Moretta, L. Cellular and molecular basis of haploidentical hematopoietic stem cell transplantation in the successful treatment of high-risk leukemias: Role of alloreactive NK cells. Front. Immunol. 2013, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Moretta, L.; Locatelli, F.; Pende, D.; Marcenaro, E.; Mingari, M.C.; Moretta, A. Killer Ig-like receptor-mediated control of natural killer cell alloreactivity in haploidentical hematopoietic stem cell transplantation. Blood 2011, 117, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Hubner, W.; Spinelli, M.A.; Chen, B.K. Predominant mode of human immunodeficiency virus transfer between T cells is mediated by sustained Env-dependent neutralization-resistant virological synapses. J. Virol. 2007, 81, 12582–12595. [Google Scholar] [CrossRef]
- Mazurov, D.; Ilinskaya, A.; Heidecker, G.; Lloyd, P.; Derse, D. Quantitative comparison of HTLV-1 and HIV-1 cell-to-cell infection with new replication dependent vectors. PLoS Pathog. 2010, 6, e1000788. [Google Scholar] [CrossRef] [PubMed]
- Bernard-Stoecklin, S.; Gommet, C.; Corneau, A.B.; Guenounou, S.; Torres, C.; Dejucq-Rainsford, N.; Cosma, A.; Dereuddre-Bosquet, N.; Le Grand, R. Semen CD4+ T cells and macrophages are productively infected at all stages of SIV infection in macaques. PLoS Pathog. 2013, 9, e1003810. [Google Scholar] [CrossRef]
- Anderson, D.J.; Politch, J.A.; Nadolski, A.M.; Blaskewicz, C.D.; Pudney, J.; Mayer, K.H. Targeting Trojan Horse leukocytes for HIV prevention. AIDS 2010, 24, 163–187. [Google Scholar] [CrossRef]
- Ivanova, D.; Krempels, R.; Ryfe, J.; Weitzman, K.; Stephenson, D.; Gigley, J.P. NK cells in mucosal defense against infection. Biomed. Res. Int. 2014, 2014, 413982. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.T.; Zhang, T.H.; Carmona, C.; Lee, B.; Seet, C.S.; Kostelny, M.; Shah, N.; Chen, H.; Farrell, K.; Soliman, M.S.A.; et al. Latency reversal plus natural killer cells diminish HIV reservoir in vivo. Nat. Commun. 2022, 13, 121. [Google Scholar] [CrossRef] [PubMed]
- Barker, E.; Evans, D.T. HLA-C Downmodulation by HIV-1 Vpu. Cell Host. Microbe 2016, 19, 570–571. [Google Scholar] [CrossRef]
- Jennes, W.; Verheyden, S.; Mertens, J.W.; Camara, M.; Seydi, M.; Dieye, T.N.; Mboup, S.; Demanet, C.; Kestens, L. Inhibitory KIR/HLA incompatibility between sexual partners confers protection against HIV-1 transmission. Blood 2013, 121, 1157–1164. [Google Scholar] [CrossRef]
- MacDonald, K.S.; Embree, J.; Njenga, S.; Nagelkerke, N.J.; Ngatia, I.; Mohammed, Z.; Barber, B.H.; Ndinya-Achola, J.; Bwayo, J.; Plummer, F.A. Mother-child class I HLA concordance increases perinatal human immunodeficiency virus type 1 transmission. J. Infect. Dis. 1998, 177, 551–556. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mackelprang, R.D.; John-Stewart, G.; Carrington, M.; Richardson, B.; Rowland-Jones, S.; Gao, X.; Mbori-Ngacha, D.; Mabuka, J.; Lohman-Payne, B.; Farquhar, C. Maternal HLA homozygosity and mother-child HLA concordance increase the risk of vertical transmission of HIV-1. J. Infect. Dis. 2008, 197, 1156–1161. [Google Scholar] [CrossRef]
- Lockett, S.F.; Robertson, J.R.; Brettle, R.P.; Yap, P.L.; Middleton, D.; Leigh Brown, A.J. Mismatched human leukocyte antigen alleles protect against heterosexual HIV transmission. J. Acquir. Immune Defic. Syndr. 2001, 27, 277–280. [Google Scholar] [CrossRef]
- Dorak, M.T.; Tang, J.; Penman-Aguilar, A.; Westfall, A.O.; Zulu, I.; Lobashevsky, E.S.; Kancheya, N.G.; Schaen, M.M.; Allen, S.A.; Kaslow, R.A. Transmission of HIV-1 and HLA-B allele-sharing within serodiscordant heterosexual Zambian couples. Lancet 2004, 363, 2137–2139. [Google Scholar] [CrossRef]
- Polycarpou, A.; Ntais, C.; Korber, B.T.; Elrich, H.A.; Winchester, R.; Krogstad, P.; Wolinsky, S.; Rostron, T.; Rowland-Jones, S.L.; Ammann, A.J.; et al. Association between maternal and infant class I and II HLA alleles and of their concordance with the risk of perinatal HIV type 1 transmission. AIDS Res. Hum. Retrovir. 2002, 18, 741–746. [Google Scholar] [CrossRef]
- Jennes, W.; Verheyden, S.; Demanet, C.; dje-Toure, C.A.; Vuylsteke, B.; Nkengasong, J.N.; Kestens, L. Cutting edge: Resistance to HIV-1 infection among African female sex workers is associated with inhibitory KIR in the absence of their HLA ligands. J. Immunol. 2006, 177, 6588–6592. [Google Scholar] [CrossRef]
- Hens, J.; Goovaerts, O.; Ceulemans, A.; Jennes, W.; Kestens, L. Impact of the Variable Killer Ig-Like Receptor-Human Leukocyte Antigen Interactions on Natural Killer Cell Cytotoxicity Toward Foreign CD4 T Cells. Front. Immunol. 2018, 9, 1588. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Klein, F.; Pietzsch, J.; Seaman, M.S.; Nussenzweig, M.C.; Ravetch, J.V. Broadly neutralizing anti-HIV-1 antibodies require Fc effector functions for in vivo activity. Cell 2014, 158, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Tomescu, C.; Seaton, K.E.; Smith, P.; Taylor, M.; Tomaras, G.D.; Metzger, D.S.; Montaner, L.J. Innate activation of MDC and NK cells in high-risk HIV-1-exposed seronegative IV-drug users who share needles when compared with low-risk nonsharing IV-drug user controls. J. Acquir. Immune Defic. Syndr. 2015, 68, 264–273. [Google Scholar] [CrossRef]
- Colon, K.; Speicher, D.W.; Smith, P.; Taylor, M.; Metzger, D.S.; Montaner, L.J.; Tomescu, C. S100A14 Is Increased in Activated NK Cells and Plasma of HIV-Exposed Seronegative People Who Inject Drugs and Promotes Monocyte-NK Crosstalk. J. Acquir. Immune Defic. Syndr. 2019, 80, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Card, C.M.; Ball, T.B.; Fowke, K.R. Immune quiescence: A model of protection against HIV infection. Retrovirology 2013, 10, 141. [Google Scholar] [CrossRef]
- Camara, M.; Dieye, T.N.; Seydi, M.; Diallo, A.A.; Fall, M.; Diaw, P.A.; Sow, P.S.; Mboup, S.; Kestens, L.; Jennes, W. Low-level CD4+ T cell activation in HIV-exposed seronegative subjects: Influence of gender and condom use. J. Infect. Dis. 2010, 201, 835–842. [Google Scholar] [CrossRef]
- Tran, H.K.; Chartier, L.; Troung, L.X.; Nguyen, N.N.; Fontanet, A.; Barre-Sinoussi, F.E.; Pancino, G.; Scott-Algara, D. Systemic immune activation in HIV-1-exposed uninfected Vietnamese intravascular drug users. AIDS Res. Hum. Retrovir. 2006, 22, 255–261. [Google Scholar] [CrossRef]
- Restrepo, C.; Rallon, N.I.; del Romero, J.; Rodriguez, C.; Hernando, V.; Lopez, M.; Peris, A.; Lozano, S.; Sempere-Ortells, J.M.; Soriano, V.; et al. Low-level exposure to HIV induces virus-specific T cell responses and immune activation in exposed HIV-seronegative individuals. J. Immunol. 2010, 185, 982–989. [Google Scholar] [CrossRef]
- Saulle, I.; Biasin, M.; Gnudi, F.; Rainone, V.; Ibba, S.V.; Lo Caputo, S.; Mazzotta, F.; Trabattoni, D.; Clerici, M. Short Communication: Immune Activation Is Present in HIV-1-Exposed Seronegative Individuals and Is Independent of Microbial Translocation. AIDS Res. Hum. Retrovir. 2016, 32, 129–133. [Google Scholar] [CrossRef]
- Biasin, M.; Caputo, S.L.; Speciale, L.; Colombo, F.; Racioppi, L.; Zagliani, A.; Ble, C.; Vichi, F.; Cianferoni, L.; Masci, A.M.; et al. Mucosal and systemic immune activation is present in human immunodeficiency virus-exposed seronegative women. J. Infect. Dis. 2000, 182, 1365–1374. [Google Scholar] [CrossRef][Green Version]
- Tomescu, C.; Duh, F.M.; Lanier, M.A.; Kapalko, A.; Mounzer, K.C.; Martin, M.P.; Carrington, M.; Metzger, D.S.; Montaner, L.J. Increased plasmacytoid dendritic cell maturation and natural killer cell activation in HIV-1 exposed, uninfected intravenous drug users. AIDS 2010, 24, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Fowke, K.R.; Nagelkerke, N.J.; Kimani, J.; Simonsen, J.N.; Anzala, A.O.; Bwayo, J.J.; MacDonald, K.S.; Ngugi, E.N.; Plummer, F.A. Resistance to HIV-1 infection among persistently seronegative prostitutes in Nairobi, Kenya. Lancet 1996, 348, 1347–1351. [Google Scholar] [CrossRef]
- Guma, M.; Cabrera, C.; Erkizia, I.; Bofill, M.; Clotet, B.; Ruiz, L.; Lopez-Botet, M. Human cytomegalovirus infection is associated with increased proportions of NK cells that express the CD94/NKG2C receptor in aviremic HIV-1-positive patients. J. Infect. Dis. 2006, 194, 38–41. [Google Scholar] [CrossRef]
- Lopez-Verges, S.; Milush, J.M.; Schwartz, B.S.; Pando, M.J.; Jarjoura, J.; York, V.A.; Houchins, J.P.; Miller, S.; Kang, S.M.; Norris, P.J.; et al. Expansion of a unique CD57(+)NKG2Chi natural killer cell subset during acute human cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2011, 108, 14725–14732. [Google Scholar] [CrossRef]
- Chang, C.; Rodríguez, A.; Carretero, M.; López-Botet, M.; Phillips, J.H.; Lanier, L.L. Molecular characterization of human CD94: A type II membrane glycoprotein related to the C-type lectin superfamily. Eur. J. Immunol. 1995, 25, 2433–2437. [Google Scholar] [CrossRef] [PubMed]
- Llano, M.; Lee, N.; Navarro, F.; Garcia, P.; Albar, J.P.; Geraghty, D.E.; Lopez-Botet, M. HLA-E-bound peptides influence recognition by inhibitory and triggering CD94/NKG2 receptors: Preferential response to an HLA-G-derived nonamer. Eur. J. Immunol. 1998, 28, 2854–2863. [Google Scholar] [CrossRef]
- Hammer, Q.; Ruckert, T.; Borst, E.M.; Dunst, J.; Haubner, A.; Durek, P.; Heinrich, F.; Gasparoni, G.; Babic, M.; Tomic, A.; et al. Peptide-specific recognition of human cytomegalovirus strains controls adaptive natural killer cells. Nat. Immunol. 2018, 19, 453–463. [Google Scholar] [CrossRef]
- Lanier, L.L.; Corliss, B.; Wu, J.; Phillips, J.H. Association of DAP12 with activating CD94/NKG2C NK cell receptors. Immunity 1998, 8, 693–701. [Google Scholar] [CrossRef]
- Schlums, H.; Cichocki, F.; Tesi, B.; Theorell, J.; Beziat, V.; Holmes, T.D.; Han, H.; Chiang, S.C.; Foley, B.; Mattsson, K.; et al. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity 2015, 42, 443–456. [Google Scholar] [CrossRef]
- Lee, J.; Zhang, T.; Hwang, I.; Kim, A.; Nitschke, L.; Kim, M.; Scott, J.M.; Kamimura, Y.; Lanier, L.L.; Kim, S. Epigenetic modification and antibody-dependent expansion of memory-like NK cells in human cytomegalovirus-infected individuals. Immunity 2015, 42, 431–442. [Google Scholar] [CrossRef]
- Hikami, K.; Tsuchiya, N.; Yabe, T.; Tokunaga, K. Variations of human killer cell lectin-like receptors: Common occurrence of NKG2-C deletion in the general population. Genes Immun. 2003, 4, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, R.; Tsuchiya, N.; Hikami, K.; Kuroki, K.; Fukazawa, T.; Bijl, M.; Kallenberg, C.G.; Hashimoto, H.; Yabe, T.; Tokunaga, K. Molecular genetic analyses of human NKG2C (KLRC2) gene deletion. Int. Immunol. 2004, 16, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.L.; Landskron, J.; Ask, E.H.; Enqvist, M.; Sohlberg, E.; Traherne, J.A.; Hammer, Q.; Goodridge, J.P.; Larsson, S.; Jayaraman, J.; et al. Critical Role of CD2 Co-stimulation in Adaptive Natural Killer Cell Responses Revealed in NKG2C-Deficient Humans. Cell Rep. 2016, 15, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Moraru, M.; Canizares, M.; Muntasell, A.; de Pablo, R.; Lopez-Botet, M.; Vilches, C. Assessment of copy-number variation in the NKG2C receptor gene in a single-tube and characterization of a reference cell panel, using standard polymerase chain reaction. Tissue Antigens 2012, 80, 184–187. [Google Scholar] [CrossRef]
- Goncalves, A.; Makalo, P.; Joof, H.; Burr, S.; Ramadhani, A.; Massae, P.; Malisa, A.; Mtuy, T.; Derrick, T.; Last, A.R.; et al. Differential frequency of NKG2C/KLRC2 deletion in distinct African populations and susceptibility to Trachoma: A new method for imputation of KLRC2 genotypes from SNP genotyping data. Hum. Genet. 2016, 135, 939–951. [Google Scholar] [CrossRef]
- Thomas, R.; Low, H.Z.; Kniesch, K.; Jacobs, R.; Schmidt, R.E.; Witte, T. NKG2C deletion is a risk factor of HIV infection. AIDS Res. Hum. Retrovir. 2012, 28, 844–851. [Google Scholar] [CrossRef]
- Alsulami, K.; Bolastig, N.; Dupuy, F.P.; Mabanga, T.; Gilbert, L.; Kiani, Z.; Routy, J.P.; Bruneau, J.; Thomas, R.; Tremblay, C.; et al. Influence of NKG2C Genotypes on HIV Susceptibility and Viral Load Set Point. J. Virol. 2021, 95, e0041721. [Google Scholar] [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de, S.M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef]
- Gray, G.E.; Bekker, L.G.; Laher, F.; Malahleha, M.; Allen, M.; Moodie, Z.; Grunenberg, N.; Huang, Y.; Grove, D.; Prigmore, B.; et al. Vaccine Efficacy of ALVAC-HIV and Bivalent Subtype C gp120-MF59 in Adults. N. Engl. J. Med. 2021, 384, 1089–1100. [Google Scholar] [CrossRef]
- Goodier, M.R.; Riley, E.M. Regulation of the human NK cell compartment by pathogens and vaccines. Clin. Trans. Immunol. 2021, 10, e1244. [Google Scholar] [CrossRef]
- Rydyznski, C.E.; Waggoner, S.N. Boosting vaccine efficacy the natural (killer) way. Trends Immunol. 2015, 36, 536–546. [Google Scholar] [CrossRef]
- Martin-Fontecha, A.; Thomsen, L.L.; Brett, S.; Gerard, C.; Lipp, M.; Lanzavecchia, A.; Sallusto, F. Induced recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1 priming. Nat. Immunol. 2004, 5, 1260–1265. [Google Scholar] [CrossRef] [PubMed]
- Farsakoglu, Y.; Palomino-Segura, M.; Latino, I.; Zanaga, S.; Chatziandreou, N.; Pizzagalli, D.U.; Rinaldi, A.; Bolis, M.; Sallusto, F.; Stein, J.V.; et al. Influenza Vaccination Induces NK-Cell-Mediated Type-II IFN Response that Regulates Humoral Immunity in an IL-6-Dependent Manner. Cell Rep. 2019, 26, 2307–2315.e5. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, M.C.; Minute, L.; Rodriguez, I.; Garasa, S.; Perez-Ruiz, E.; Inoges, S.; Melero, I.; Berraondo, P. Antibody-dependent cell cytotoxicity: Immunotherapy strategies enhancing effector NK cells. Immunol. Cell Biol. 2017, 95, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Li, A.P.Y.; Cohen, C.A.; Leung, N.H.L.; Fang, V.J.; Gangappa, S.; Sambhara, S.; Levine, M.Z.; Iuliano, A.D.; Perera, R.; Ip, D.K.M.; et al. Immunogenicity of standard, high-dose, MF59-adjuvanted, and recombinant-HA seasonal influenza vaccination in older adults. NPJ Vaccines 2021, 6, 25. [Google Scholar] [CrossRef] [PubMed]
- Coccia, M.; Collignon, C.; Herve, C.; Chalon, A.; Welsby, I.; Detienne, S.; van Helden, M.J.; Dutta, S.; Genito, C.J.; Waters, N.C.; et al. Cellular and molecular synergy in AS01-adjuvanted vaccines results in an early IFNgamma response promoting vaccine immunogenicity. NPJ Vaccines 2017, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Howard, L.M.; Hoek, K.L.; Goll, J.B.; Samir, P.; Galassie, A.; Allos, T.M.; Niu, X.; Gordy, L.E.; Creech, C.B.; Prasad, N.; et al. Cell-Based Systems Biology Analysis of Human AS03-Adjuvanted H5N1 Avian Influenza Vaccine Responses: A Phase I Randomized Controlled Trial. PLoS ONE 2017, 12, e0167488. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.; Cevik, H.; Feldman, H.A.; Canaday, L.M.; Lakes, N.; Waggoner, S.N. Targeting natural killer cells to enhance vaccine responses. Trends Pharmacol. Sci. 2021, 42, 789–801. [Google Scholar] [CrossRef]
- Luetke-Eversloh, M.; Hammer, Q.; Durek, P.; Nordstrom, K.; Gasparoni, G.; Pink, M.; Hamann, A.; Walter, J.; Chang, H.D.; Dong, J.; et al. Human cytomegalovirus drives epigenetic imprinting of the IFNG locus in NKG2Chi natural killer cells. PLoS Pathog 2014, 10, e1004441. [Google Scholar] [CrossRef]
- Zhang, T.; Scott, J.M.; Hwang, I.; Kim, S. Cutting edge: Antibody-dependent memory-like NK cells distinguished by FcRgamma deficiency. J. Immunol. 2013, 190, 1402–1406. [Google Scholar] [CrossRef]
- Gyurova, I.E.; Ali, A.; Waggoner, S.N. Natural Killer Cell Regulation of B Cell Responses in the Context of Viral Infection. Viral Immunol 2020, 33, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Rydyznski, C.; Daniels, K.A.; Karmele, E.P.; Brooks, T.R.; Mahl, S.E.; Moran, M.T.; Li, C.; Sutiwisesak, R.; Welsh, R.M.; Waggoner, S.N. Generation of cellular immune memory and B-cell immunity is impaired by natural killer cells. Nat. Commun. 2015, 6, 6375. [Google Scholar] [CrossRef] [PubMed]
- Rydyznski, C.E.; Cranert, S.A.; Zhou, J.Q.; Xu, H.; Kleinstein, S.H.; Singh, H.; Waggoner, S.N. Affinity Maturation Is Impaired by Natural Killer Cell Suppression of Germinal Centers. Cell Rep. 2018, 24, 3367–3373.e4. [Google Scholar] [CrossRef]
- Dupuy, F.P.; Kant, S.; Barbe, A.; Routy, J.P.; Bruneau, J.; Lebouche, B.; Tremblay, C.; Pazgier, M.; Finzi, A.; Bernard, N.F. Antibody-Dependent Cellular Cytotoxicity-Competent Antibodies against HIV-1-Infected Cells in Plasma from HIV-Infected Subjects. MBio 2019, 10, e02690-19. [Google Scholar] [CrossRef]
- Richard, J.; Prevost, J.; Alsahafi, N.; Ding, S.; Finzi, A. Impact of HIV-1 Envelope Conformation on ADCC Responses. Trends Microbiol. 2018, 26, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.; Prevost, J.; Baxter, A.E.; von Bredow, B.; Ding, S.; Medjahed, H.; Delgado, G.G.; Brassard, N.; Sturzel, C.M.; Kirchhoff, F.; et al. Uninfected Bystander Cells Impact the Measurement of HIV-Specific Antibody-Dependent Cellular Cytotoxicity Responses. MBio 2018, 9, e00358-18. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).