
Variable Gene Expression in Human Ganglia Latently Infected with Varicella-Zoster Virus


Abstract
:1. Introduction
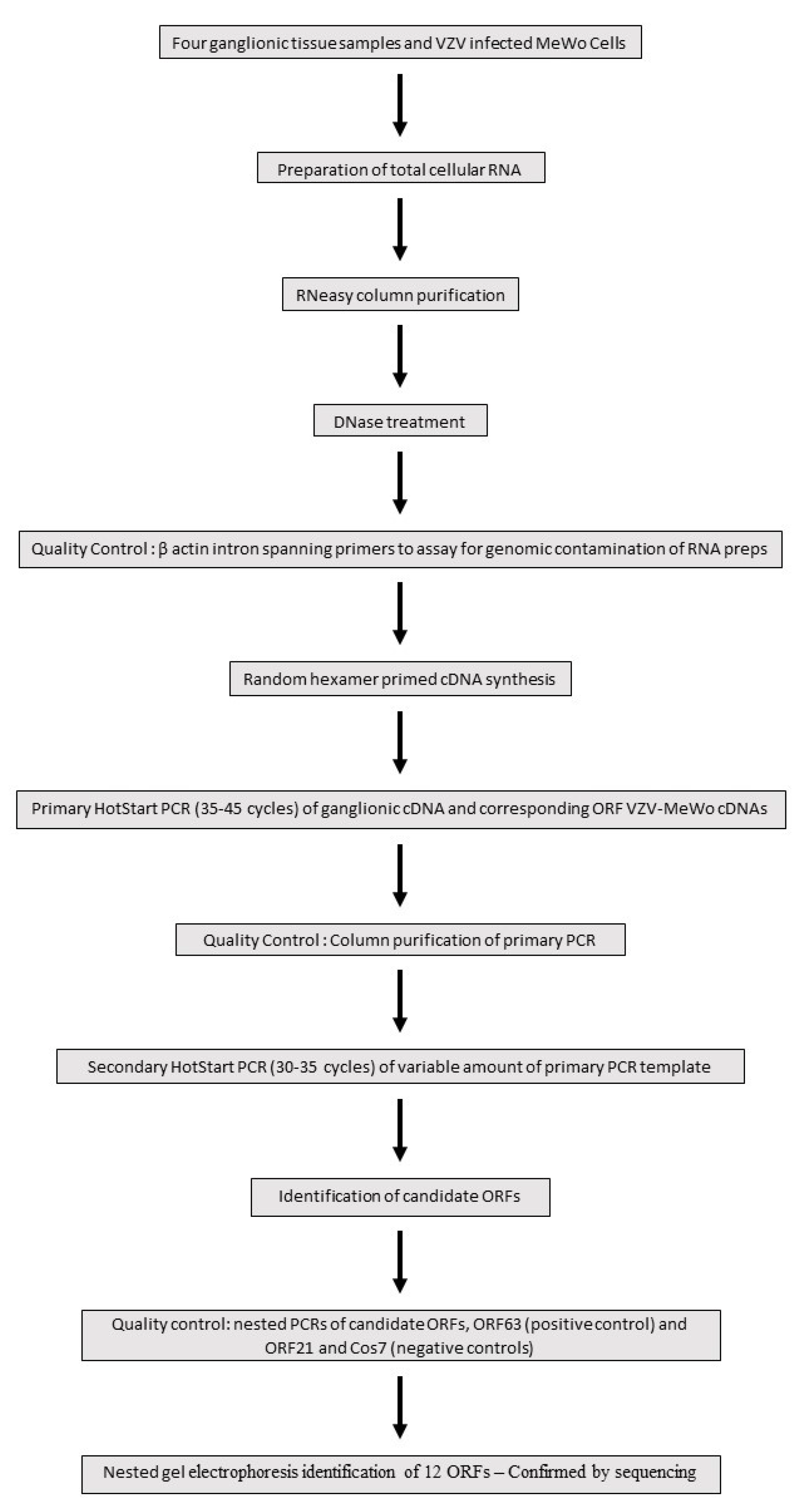
2. Materials and Methods
2.1. Tissues and RNA Extraction
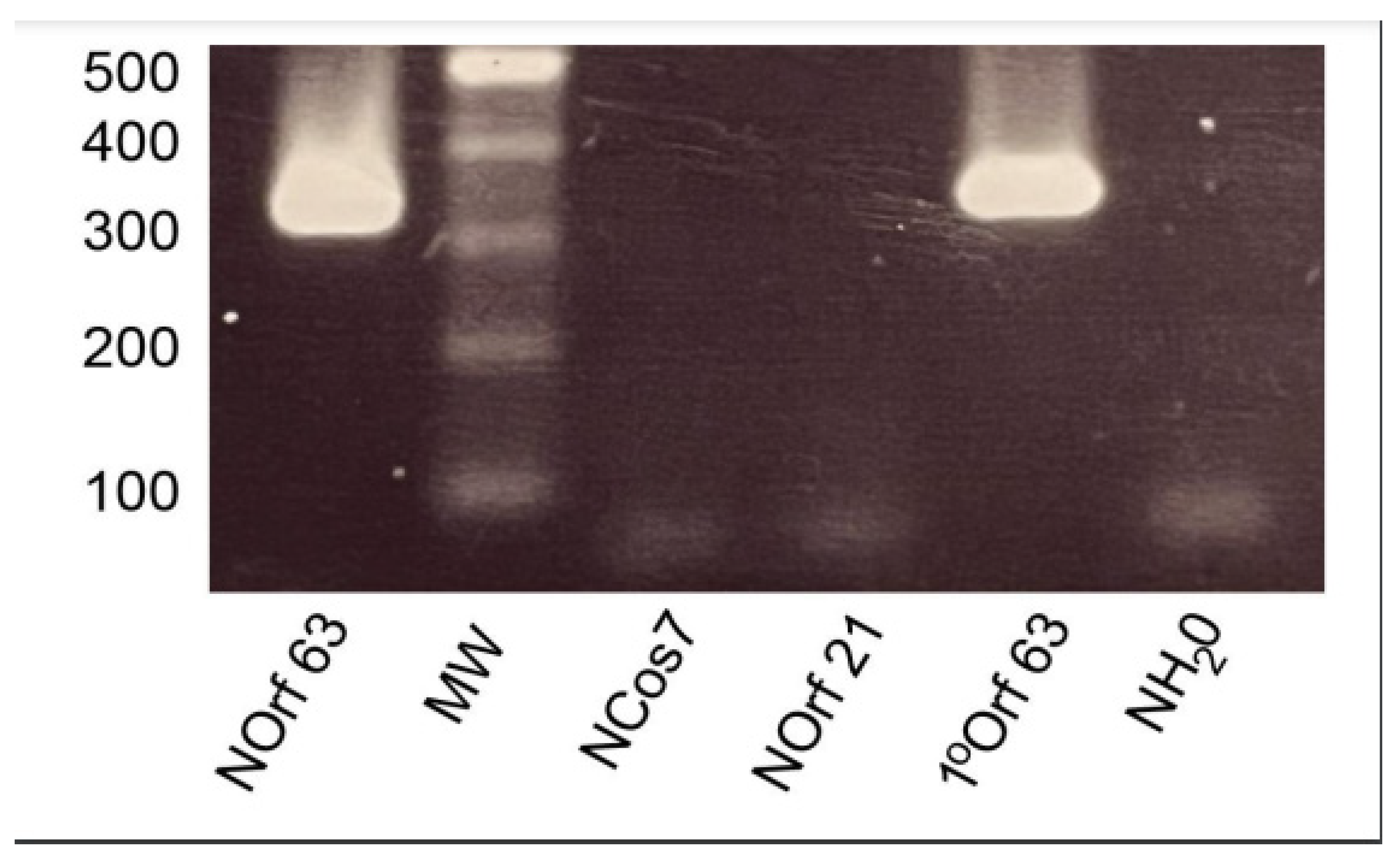
2.2. Nested RT-PCR Assays
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.E.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Prim. 2015, 1, 15016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E.; Gershon, A.A. Clinical features of Varicella-Zoster virus infection of the nervous system. Viruses 2018, 10, 609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E.; Grinfield, E.; Gow, J.W. Latent varicella-zoster virus is located predominantly in neurons in human trigeminal ganglia. Proc. Natl. Acad. Sci. USA 1998, 95, 4658–4662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E.; Mogensen, T.H.; Cohrs, R.J. Recent issues in Varicella-Zoster Virus latency. Virology 2021, 13, 2018. [Google Scholar] [CrossRef]
- Cohrs, R.J.; Srock, K.; Barbour, M.B.; Owens, G.; Mahalingam, R.; Devlin, M.E.; Wellish, M.; Gilden, D.H. Varicella-zoster virus (VZV) transcription during latency in human ganglia: Construction of a cDNA library from latently infected human trigeminal ganglia and detection of a VZV transcript. J. Virol. 1994, 68, 7900–7908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohrs, R.J.; Barbour, M.; Gilden, D.H. Varicella-zoster virus (VZV) transcription during latency in human ganglia: Detection of transcripts mapping to genes 21, 29, 62, and 63 in a cDNA library enriched for VZV RNA. J. Virol. 1996, 70, 2789–2796. [Google Scholar] [CrossRef] [Green Version]
- Meier, J.L.; Holman, R.P.; Croen, K.D.; Smialek, J.E.; Straus, S.E. Varicella-zoster virus transcription in human trigeminal ganglia. Virology 1993, 193, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G.E.; Grinfeld, E.; Bell, J.E. Varicella-zoster virus gene expression in latently infected andexplanted human ganglia. J. Virol. 2000, 74, 11893–11898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohrs, R.J.; Gilden, D.H.; Kinchington, P.R.; Grinfeld, E.; Kennedy, P.G.E. Varicella-Zoster Virus Gene 66Transcription and Translation in Latently Infected Human Ganglia. J. Virol. 2003, 77, 6660–6665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, M.A.; Choe, A.; Traktinskiy, I.; Cordery-Cotter, R.; Gilden, D.; Cohrs, R.J. Varicella-zoster virus transcriptome in latently infected human ganglia. J. Virol. 2011, 85, 2276–2287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depledge, D.P.; Ouwendijk, W.; Sadaoka, T.; Braspenning, S.E.; Mori, Y.; Cohrs, R.J.; Verjans, G.M.G.M.; Breuer, J. A spliced latency associated VZV transcript maps antisense to the viral transactivator gene 61. Nat. Commun. 2018, 9, 1167. [Google Scholar] [CrossRef] [PubMed]
- Storlie, J.; Maresova, L.; Jackson, W.; Grose, C. Comparative analyses of the 9 glycoprotein genes found in wild-type and vaccine strains of varicella-zoster virus. J. Infect. Dis. 2008, 197 (Suppl. S2), S49–S53. [Google Scholar] [CrossRef] [PubMed]
- Visalli, R.J.; Nicolosi, D.M.; Irven, K.L.; Goshorn, B.; Khan, T.; Visalli, M.A. The Varicella-zoster virus DNA encapsidation genes: Identification and characterization of the putative terminase subunits. Virus Res. 2007, 129, 200–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.M.; Cox, E.; Iofin, I.; Soong, W.; Cohen, J.I. Varicella-zoster virus (VZV) ORF32 encodes a phosphoprotein that is posttranslationally modified by the VZV ORF47 protein kinase. J. Virol. 1998, 72, 8083–8088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshii, H.; Sadaoka, K.; Matsuura, M.; Nagaike, K.; Takahashi, M.; Yamanishi, K.; Mori, Y. Varicella-zoster virus ORF 58 gene is dispensable for viral replication in cell culture. Virol. J. 2008, 5, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, P.G.E.; Rovnak, J.; Badani, H.; Cohrs, R.J. A comparison of herpes simplex virus type 1 and varicella-zoster virus latency and reactivation. J. Gen. Virol. 2015, 96 Pt 7, 1581–1602. [Google Scholar] [CrossRef] [PubMed]
- Ouwendijk, W.J.D.; Depledge, D.P.; Rajbhandari, L.; Lenac Roviš, T.; Jonjic, S.; Breuer, J.; Venkatesan, A.; Verjans, G.M.G.M.; Sadaoka, T. Varicella-zoster virus VLT-ORF63 fusion transcript induces broad viral gene expression during reactivation from neuronal latency. Nat. Commun. 2020, 11, 6324. [Google Scholar] [CrossRef] [PubMed]
- Bisht, P.; Das, B.; Kinchington, P.R.; Goldstein, R.S. Varicella-Zoster Virus (VZV) Small Noncoding RNAs Antisense to the VZV Latency-Encoded Transcript VLT Enhance Viral Replication. J. Virol. 2020, 94, e00123-20. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Donor Designated Ganglionic Samples | |||||
|---|---|---|---|---|---|
| Classic ORFs | Proposed Function | S24 | S27 | S35 | S36 |
| ORF4 | IE Transcriptional activator | – | + | + | + |
| ORF21 | E Replication in culture | – | – | – | – |
| ORF29 | E Transcriptional modulator | + | – | + | + |
| ORF40 | L Encapsidation | – | – | – | – |
| ORF62 | IE Transcriptional activator | + | – | + | – |
| ORF63 | IE Transcriptional activator | + | + | + | + |
| ORF66 | E Protein kinase—Unknown | – | – | – | – |
| Nagel et al. [10] | |||||
| ORF11 | Tegument protein | – | – | + | + |
| ORF41 | Essential for growth in culture—Unknown | – | + | + | + |
| ORF43 | Essential for growth in culture—Unknown | – | – | – | – |
| ORF57 | Unknown | – | + | – | + |
| ORF68 | Glycoprotein | – | – | – | – |
| Novel candidates | |||||
| ORF32 | Substrate for ORF47 | + | – | + | + |
| ORF37 | L Glycoprotein | – | – | + | + |
| ORF42 | Unknown | + | – | + | – |
| ORF58 | Dispensable for replication—Unknown | – | – | + | – |
| ORF60 | L Protein kinase—viral replication | – | – | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kennedy, P.G.E.; Montague, P. Variable Gene Expression in Human Ganglia Latently Infected with Varicella-Zoster Virus. Viruses 2022, 14, 1250. https://doi.org/10.3390/v14061250
Kennedy PGE, Montague P. Variable Gene Expression in Human Ganglia Latently Infected with Varicella-Zoster Virus. Viruses. 2022; 14(6):1250. https://doi.org/10.3390/v14061250
Chicago/Turabian StyleKennedy, Peter G. E., and Paul Montague. 2022. "Variable Gene Expression in Human Ganglia Latently Infected with Varicella-Zoster Virus" Viruses 14, no. 6: 1250. https://doi.org/10.3390/v14061250
APA StyleKennedy, P. G. E., & Montague, P. (2022). Variable Gene Expression in Human Ganglia Latently Infected with Varicella-Zoster Virus. Viruses, 14(6), 1250. https://doi.org/10.3390/v14061250