Abstract
Orthohantaviruses are zoonotic pathogens that play a significant role in public health. These viruses can cause haemorrhagic fever with renal syndrome in Eurasia. In the Republic of Kazakhstan, the first human cases were registered in the year 2000 in the West Kazakhstan region. Small mammals can be reservoirs of orthohantaviruses. Previous studies showed orthohantavirus antigens in wild-living small mammals in four districts of West Kazakhstan. Clinical studies suggested that there might be further regions with human orthohantavirus infections in Kazakhstan, but genetic data of orthohantaviruses in natural foci are limited. The aim of this study was to investigate small mammals for the presence of orthohantaviruses by molecular biological methods and to provide a phylogenetic characterization of the circulating strains in Kazakhstan. Small mammals were trapped at 19 sites in West Kazakhstan, four in Almaty region and at seven sites around Almaty city during all seasons of 2018 and 2019. Lung tissues of small mammals were homogenized and RNA was extracted. Orthohantavirus RT-PCR assays were applied for detection of partial S and L segment sequences. Results were compared to published fragments. In total, 621 small mammals from 11 species were analysed. Among the collected small mammals, 2.4% tested positive for orthohantavirus RNA, one sample from West Kazakhstan and 14 samples from Almaty region. None of the rodents caught in Almaty city were infected. Sequencing parts of the small (S) and large (L) segments specified Tula virus (TULV) in these two regions. Our data show that geographical distribution of TULV is more extended as previously thought. The detected sequences were found to be split in two distinct genetic clusters of TULV in West Kazakhstan and Almaty region. TULV was detected in the common vole (Microtus arvalis) and for the first time in two individuals of the forest dormouse (Dryomys nitedula), interpreted as a spill-over infection in Kazakhstan.
1. Introduction
The genus Orthohantavirus (family Hantaviridae, order Bunyavirales) includes zoonotic pathogens. This group of viruses plays an important role in causing human diseases worldwide. Orthohantaviruses are single-stranded negative polarity RNA viruses, and the genome consists of three segments. The large (L) segment encodes a viral RNA-dependent RNA polymerase, the medium (M) segment encodes the glycoprotein precursor (GPC), which is processed to the glycoproteins Gn and Gc, and the small (S) segment encodes the nucleocapsid (N) protein [1].
Small mammal species are a reservoir for orthohantaviruses. Orthohantaviruses are presently known to infect rodents (subfamilies Murinae, Arvicolinae, Sigmodontinae, and Neotominae), but are also detected in different shrews and moles [2,3,4]. In Eurasia, humans are infected either by rare direct contact or indirectly by inhalation of orthohantaviruses containing dust from dried excreta [5,6].
Old World orthohantaviruses can cause haemorrhagic fever with renal syndrome (HFRS) and are mainly transmitted by members of the Murinae and Arvicolinae subfamilies [2,3]. In Europe, the main causative agent of HFRS is Puumala virus (PUUV) causing nephropathia epidemica (NE), a mild form of HFRS. A mild to severe form of HFRS is caused by Dobrava-Belgrade virus (DOBV). In Asia, the most relevant species is Hantaan virus (HNTV) that causes a severe form of HFRS. Seoul virus (SEOV) is distributed worldwide and can cause a moderate form of HFRS [6,7,8,9]. Pathogenicity of Tula virus (TULV) to humans is limited, only few reports of human cases were described in Europe [10,11,12,13], despite the fact that TULV is found in Asia and Europe. In North America, the TULV-related Prospect Hill virus was identified in a Microtus species (M. pennsylvanicus) but no human infections have been reported here either [2,7,14,15].
The Central Asian Republic of Kazakhstan has a vast territory and contains several types of landscapes such as forest-steppes, steppes, semi-deserts, deserts, and mountain ranges [16,17]. In these different geographic settings, Kazakhstan has numerous natural foci of important zoonotic pathogens such as Yersinia pestis, Bacillus anthracis, Francisella tularensis, Leptospira, Listeria monocytogenes, tick-borne encephalitis virus (TBEV), Crimean-Congo haemorrhagic fever virus (CCHFV), and orthohantaviruses [17,18].
An investigation of small mammals on the Dzungarian Alatau mountain range in Almaty region in 1990–1993 showed that some rodents contain orthohantavirus antigens (n = 644, 5.3%) [19]. Twenty years later, a study conducted in the same region using antigen assays found traces of orthohantavirus antigens in 2.2% of investigated tissue suspensions of rodents collected in 2010–2016 [20,21]. Furthermore, the existence of Tula virus was proven in tissue samples of Microtus arvalis in Almaty region (periphery of Taldykorgan city and Karatal village) [22].
The first human case of HFRS was detected in the year 2000 in the Zharsuat village in the Borili district, a part of the West Kazakhstan region [23,24]. Further investigations of host reservoirs were started, and from 2001 to 2011 almost 50,000 small mammals including 30 species were screened for the presence of orthohantavirus antigen. A total of 1.53% of different species, mostly Myodes glareolus, Microtus arvalis, Microtus minutus, Apodemus uralensis, and Mus musculus were positive. Therefore, so far, natural foci of orthohantaviruses were described in the four northern districts of the West Kazakhstan region (Borili, Bayterek, Shyngyrlau, and Terekti) and very preliminary in the Aktobe region [25,26]. However, in all investigations on orthohantaviruses in West Kazakhstan, contemporary molecular methods were never applied.
To date, there have been no officially registered human cases of HFRS in the Almaty region. However, an investigation of patients with fever of unknown origin (FUO) in Almaty and Kyzylorda regions showed orthohantavirus-reactive antibodies in sera of patients. This indicates that orthohantaviruses might also be endemic in the southeast of Kazakhstan [23].
The aim of this study was to investigate small mammals for the presence of orthohantaviruses by molecular biological methods in the Almaty region, including Almaty city and in West Kazakhstan, representing an officially endemic region for orthohantavirus infections in humans.
2. Materials and Methods
2.1. Study Setting and Rodent Sampling
Small mammals were trapped in 2018 and 2019 in West Kazakhstan (Bayterek, Borili, Terekti, and Taskaly districts: 19 trapping sites), Almaty region (surroundings of Tekeli city, Rudniychniy, and Bakanas: four trapping sites) and Almaty city (seven trapping sites) during spring, summer, autumn, and winter seasons (Figure 1 and Supplementary Table S1).
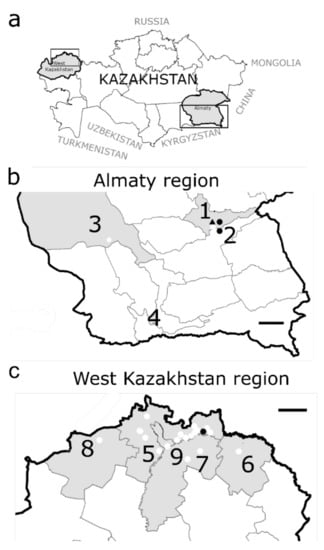
Figure 1.
Geographical location of the sampling points for small mammals in Kazakhstan. (a): Kazakhstan is divided in 14 oblasts (=regions) and located in Central Asia. (b): Almaty region and Almaty city: 1. Tekeli city: 2 trapping sites; 2. village Rudniychniy: 1 trapping site; 3. village Bakanas: 1 trapping site; 4. Almaty city: 7 trapping sites; (c): West Kazakhstan region: 5. district Bayterek: 12 trapping sites; 6. district Borili: 1 trapping site; 7. district Terekti: 2 trapping sites; 8. district Taskala: 1 trapping site; 9. Oral city: 3 trapping sites. Sampling locations: white dots. Species and location of infected rodents: ● Microtus arvalis, ▲ Dryomys nitedula. Black frames = regions magnified in (b) and (c), size marker = 150 km.
Snap traps were set overnight at 5 m intervals baited with cured pork fat. In the early morning, captured small mammals were collected, stored on dry ice, and transported to the laboratory for immediate processing. After morphological identification of the species, necropsy was performed, and internal organs (lung, heart, brain, kidney, liver, spleen, ears, and transudate) were aseptically collected and stored in RNA later (Thermo Scientific, Langenselbold, Germany) at −20 °C until further use [27].
2.2. RNA Extraction, PCR Amplification and Sequencing
Lung tissue samples were homogenized in 1 mL MEM for 2 min at 30 Hz in a TissueLyser II (Qiagen, Hilden, Germany). RNA was extracted from 140 µL homogenized supernatant using a commercial QiAmp Viral RNA Mini Kit (Qiagen, Hilden, Germany) according to manufacturer’s instructions. To determine the sequences of parts of the S and L segments, RNA was reverse-transcribed and amplified using primers detecting a variety of orthohantaviruses and subsequently sequenced using terminator cycle sequencing. In detail, for the S segment, a conventional PCR was applied using Superscript III one step RT-PCR system with Platinum Taq high fidelity polymerase (Invitrogen, Langenselbold, Germany) and the primers DOBV-M6 (5′-AGYCCWGTNATGRGWGTRATTGG-3′) and DOBV-M8 (5′-GAKGCCATRATNGTRTTYCKCATRTCCTG-3′), as described elsewhere [28,29]. The RT-PCR products were analysed using a 1.5% agarose gel with an expected amplicon size of 380 base pairs (bp). To detect a partial L-segment sequence (230 bp), a real-time RT-PCR using a Qiagen One Step RT-PCR mix was performed. Here, the primer-mix contained forward (1a-fw: 5′-TGATGCATATTGTGTGCAGAC-3′, 1b-fw: 5′-TGATGCATACTGTGTGCAAAC-3′, 1c-fw: 5′-CAGTATGATGCATACTGTGTCCAA-3′, 1d-fw: 5′-TGATGCCTATTGTGTTCAGAC-3′) and reverse (1a-rev: 5′-CTTGCTCTGTTTTGAATCTCA-3′, 1b-rev: 5′-CTTGCTCGGTGTTGAATCGCA-3′, 1c-rev: 5′-CCTGTTCTGTATTAAATCTCA-3′, 1d-rev: 5′-CTTGTTCAGTCTTGAATCTCA-3′) (0.125 µM each) primers, complemented with EvaGreen (VWR International, Vienna, Austria) as PCR reagents [30].
To confirm the species determination of the small mammals, a cytochrome b (mt-Cytb) gene sequencing was applied as described in [31]. For analysis of the mitochondrially encoded Cytb, supernatant from homogenised rodent lung tissue in elution buffer (Qiagen, Hilden, Germany) was used. A total of 400 ng of DNA were amplified by PCR using the primer combination Cytb-Uni-fw (5′-TCATCMTGATGAAAYTTYGG-3′) and Cytb-Uni-rev (5′-ACTGGYTGDCCBCCRATTCA-3′) targeting an approximately 1000 bp long fragment. The PCR was enabled by using the Invitrogen Platinum Taq High Fidelity DNA Polymerase (ThermoFisher Scientific, Langenselbold, Germany).
All positive PCR products (fragments of the S and L segment, Cytb fragments) were purified using a QIAquick PCR purification Kit (Qiagen, Hilden, Germany) and sequenced according to the manufacturer’s instructions by using a BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Langenselbold, Germany) and a 3730xl DNA Analyzer (Applied Biosystems, Langenselbold, Germany).
2.3. Phylogenetic Analysis
The generated nucleotide sequences were aligned using the ClustalW method in Bioedit 7.2.5. Prior to alignment, the sequences were trimmed for the primers resulting in final sequence lengths of 346 nucleotides (nt) for the S segment and 184 nt for the L segment that were then used for the phylogenetic analysis. Phylogenetic trees were constructed in MEGA X with the Maximum Likelihood method based on the Tamura 3-parameter model [32]. These analyses involved published S and L segment nucleotide sequences from GenBank trimmed to the same length with accession numbers listed in the captions to Figure 2 and Figure 3. To set an outgroup in the phylogenetic trees, sequences of PUUV S and L segments, also trimmed to the respective lengths, were used (NC005224 and NC005225, respectively).
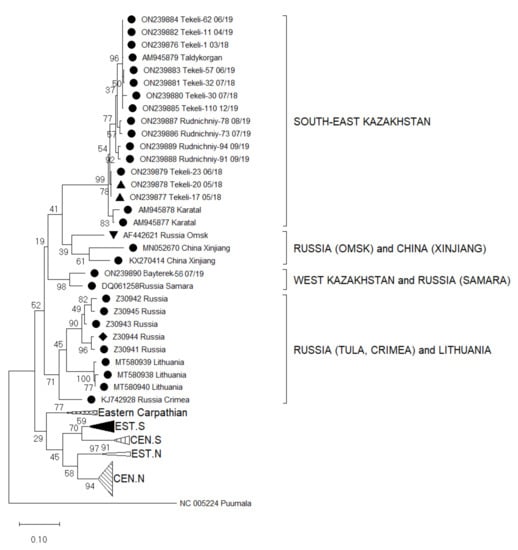
Figure 2.
Phylogenetic analysis by Maximum Likelihood method of the S segments (346 nucleotides (nt), positions of sequences 715–1061 nt in regard to the reference sequence AM945879) of Tula virus in Kazakhstan. The tree with the highest log likelihood (−5756.38) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. This analysis involved 92 nucleotide sequences: Central North (CEN.N): KU139579, KU139576, KU139577, KU139578, DQ662094, HQ697346, HQ697344, HQ697347, HQ697351, GU300137, GU300136, EU439952, EU439947, EU439949, EU439948, EU439950, EU439946, EU439951, KU139534, KU139535, KU139537, KU139538, KU139598, KU139595, KU139596, KU139599, KU139529, KU139528, KU139531, KU139530, KU139533, DQ662087, DQ768143; Eastern North (EST.N): AF063897, AF289819, AF289820, AF289821; Central South (CEN.S): AF164093, HQ697350, HQ697348, HQ697349, HQ697355, HQ697353, HQ697354, HQ697357; Eastern South (EST.S): AJ223601, U95312, KF184327, KF184328, NC005227, Z69991, Z49915, Z48741, AJ223600, Z48574, KU139560; Eastern Carpathian: AF017659, Y13980, KF557547, Y13979; Russia Tula: Z30941, Z30942, Z30943, Z30944, Z30945; Russia Crimea: KJ742928; Lithuania: MT580938, MT580939, MT580940; Russia Samara: DQ061258; Russia Omsk: AF442621; China Xinjiang: MN052670, KX270414; South-East Kazakhstan: AM945877, AM945878, AM945879, outgroup Puumala NC005224. Host Species: ● Microtus arvalis, ▲ Dryomys nitedula, ♦ Microtus rossiaemeridionalis, ▼ Microtus gregalis.
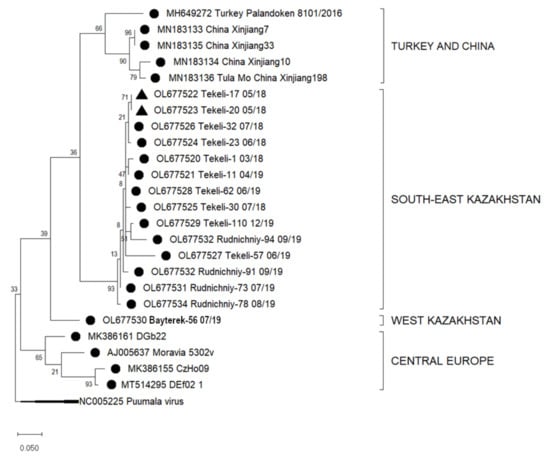
Figure 3.
Phylogenetic analysis by Maximum Likelihood method of the L segments (184 nucleotides (nt), positions of sequences 5187–5371 nt in regard to the reference sequence NC005226) of Tula virus in Kazakhstan. The tree with the highest log likelihood (−1345.67) is shown. The percentage of trees in which the associated taxa clustered together is shown next to the branches. This analysis involved 25 nucleotide sequences: Turkey: MH649272; China: MN183133, MN183135, MN183134, MN183136; Europe: AJ005637, MK386161, MK386155, MT514295, outgroup Puumala NC005225. Host Species: ● Microtus arvalis, ▲ Dryomys nitedula..
3. Results
In total, 621 small mammals were collected in nine sampling areas, at all together 30 trapping sites during the years of 2018–2019 (Table 1).

Table 1.
All species captured in snap traps in the sampling areas of interest.
These small mammals represent eleven species from four families: Cricetidae (M. arvalis, M. glareolus, M. kirgisorum), Muridae (A. uralensis, M. musculus, R. norvegicus, M. meridianus), Gliridae (D. nitedula) and Soricidae (S. araneus, S. minutus, C. suaveolens). Sex distribution of collected mammals was almost equal with 59% male and 41% female.
Out of all 621 collected small mammals 15 (2.4%) were positive for orthohantavirus RNA (Supplementary Table S1). In Almaty city, all analysed rodents failed to yield a positive result. The infected individuals represented two species, M. arvalis (n = 13, 15.1%) and D. nitedula (n = 2, 13.3%) (Table 2). Three M. arvalis and both of the orthohantavirus carrying D. nitedula samples were further tested by cytochrome b gene-specific PCR and subsequent sequence analysis [31] to confirm the morphological determination. The Cytb sequence of Tekeli23 M. arvalis (ON513439) was 99% similar to a nucleotide sequence of M. arvalis originating from Russia, Ekaterinburg (MG703092). Both the D. nitedula Tekeli17 (ON513437) and Tekeli20 (ON513438) species were also confirmed by mitochondrial cytochrome b sequencing. The two sequences are 98% identical to a sequence from D. nitedula described from Mongolia (LR131101). All orthohantavirus infected specimens where either adults (n = 11) or sub-adults (n = 4).
Table 2.
Result of the molecular biological screen for orthohantavirus RNA among small mammals captured in the regions of interest.
A partial S segment sequence analysis revealed that all 15 small mammals harboured RNA of TULV. The obtained sequences were aligned with published TULV partial S segments available for Central Asia, Eastern and Central Europe, and China. These included clades from different geographic regions such as Central North (CEN.N), Eastern North (EST.N), Central South (CEN.S), Eastern South (EST.S), Eastern Carpathian, Russia (Tula, Crimea, Samara, and Omsk), Lithuania, and China (Xinjiang) (Figure 2). A nucleotide sequence identity matrix of the detected S segments compared with sequences of geographically relevant regions reveals that the sequences have an identity range from 78.9–100% (Table 3).
Table 3.
Nucleotide sequence identity of the partial Tula virus (TULV) S-segments detected from Kazakhstan in comparison with published sequences from other Eurasian regions (%).
By comparing the newly identified TULV sequences with published genomes, four clusters can be classified that are geographically relevant for Kazakhstan (Figure 2): (I) The South-East Kazakhstan cluster consists of new virus sequences from Tekeli and Rudnichniy and already published sequences from Taldykorgan (AM945879) and Karatal (AM945877, AM945878) with a nucleotide sequence identity range of 94.3–100%. (II) The second neighbouring cluster from China and Russia includes sequences from Xinjiang (KX270414, MN052670) and from Omsk in Russian Siberia (AF442621) with a nucleotide sequence identity ranging from 84.5–87.5% within the cluster. (III) The third cluster are sequences from the Tula area of Russia (Z30941-4) and from Crimea (KJ742928) with an identity range of 87.5–98.5%. (IV) One positive sample (M. arvalis, Bayterek-56 07/19) from West Kazakhstan had a 93.4% sequence identity with the Samara virus from Russia (DQ061258). These two virus sequences form a separate cluster from all the other sequenced viruses (Figure 2).
A 78.9–99.4% nucleotide sequence identity is noticeable between the cluster of southeast Kazakhstan (I) that contains genomes form China and Siberia (II), as well as among the clade of Tula and Crimea area of Russia (III) and with the new sequence from West Kazakhstan (IV). The sequences from southeast Kazakhstan (I) are 75.8–99.1% similar to the Samara virus of Russia (IV).
The sequences from West Kazakhstan have 84.5–98.5% identity with variants from the Tula region and Crimea (III) and 82.1–87.5% identity with genomes from China and Siberia (II), respectively.
In silico translated S segment sequences of all TULV sequences included in this study showed 86–100% amino acid sequence identity for the N protein to other variants (Supplementary Figure S1).
Similarly, the sequences of parts of the L segment from Almaty and West Kazakhstan regions were aligned with other L segment sequences available from GenBank. These resulted in four clusters of TULV from various geographic locations. Sequences of the 14 samples from Almaty region grouped in one subcluster (South-East Kazakhstan, I), sequences from China (Xinjiang, MN183133-6) and Turkey (Palandoken, MH649272) in a second cluster (II). These sequences show nucleotide sequence similarities of 80–99.3%. One sample from West Kazakhstan (Bayterek-56 07/19, M. arvalis, III) grouped distant from the other sequences (Figure 3) and had a nucleotide sequence similarity of 80.6–99.3% to the samples from South-East Kazakhstan (I) (Table 4).
Table 4.
Nucleotide sequence identity of the partial Tula virus (TULV) L segment sequences in Kazakhstan and other Eurasian regions (%).
By translating these nucleotide sequences into its short peptide sequence of 61 amino acids, two recurring substitutions become apparent. The sequences Tekeli-110 (OL677529) and Rudnichniy-94 (OL677532) show at position 1760 a P versus R exchange and at position 1773 a K versus E aberration in comparison to published consensus sequences (Supplemental Figure S2).
4. Discussion
We designed a study to screen for orthohantavirus RNA in small mammals in the Republic of Kazakhstan regions West Kazakhstan, Almaty region, and Almaty city. Here, we demonstrate for the first time the presence of TULV in West Kazakhstan and confirm it in the Almaty region in Kazakhstan. The rate of positive individuals of M. arvalis is 15.1% (13/86), which agrees with previous studies [33,34]. Among all positive samples, males accounted for 60% (n = 9), which is consistent with other studies showing that male small mammals have a greater infection rate for orthohantaviruses (Table 2) [35].
West Kazakhstan is the only official orthohantavirus endemic region with registered human cases of orthohantaviruses infections so far [36,37]. Long-term investigations of host reservoirs starting from 2001 by colleagues from the Oral antiplague station revealed natural foci of orthohantaviruses in the floodplains of the Ural River. This area directly borders the Russian Orenburg and Samara regions, where orthohantavirus is also endemic [26,38]. Several small mammals that are also spread in this region such as M. glareolus, M. arvalis, A. uralensis, and M. musculus contained orthohantavirus antigens [26]. Our study could confirm the existence of TULV in West Kazakhstan region in M. arvalis, but only in one specimen. Actually, we expected to find the presence of PUUV, due to clinical manifestations of hospitalized patients with HFRS that is primarily caused by PUUV. Additionally, M. glareolus, the main host reservoir of PUUV is very common in this region. However, the number of captured M. glareolus and other small mammals was rather low to draw a statistically convincing picture on the spread of orthohantavirus in this area. Still, this study is the first to perform molecular-biological methods in the region of West Kazakhstan and generated the first orthohantavirus sequence from TULV [26,37].
In this study, for the first time, small mammals were screened for the presence of orthohantaviruses in Almaty city, but no positive results were revealed in the captured rodent species that were M. kirgisorum, A. uralensis, R. norvegicus, and M. musculus. The latter where the most captured animals in Almaty in this study. All these species might carry different orthohantaviruses such as, e.g., SEOV, but the primers used in this study are detecting all species of orthohantaviruses as shown in an internal validation of the primer sets for certified diagnostics [39]. The reason why there were no traces of orthohantavirus detected in the city are manifold but may rest in the different living conditions and species composition of the rodent population. However, as PUUV-reactive antibodies were found in a retrospective study in patients with fever of unknown origin [23], further studies have to be conducted in different geographic areas of Almaty city in order to unveil the real prevalence in the city.
Nevertheless, in the Almaty region, an area stretching north of Almaty city, TULV was identified and sequenced in several specimens captured in Tekeli city and Rudnichniy village. All TULV RNA was detected in two different species of small mammals, M. arvalis and D. nitedula. M. arvalis is a commonly known host for TULV. Interestingly, however, we also found TULV in D. nitedula of the Gliridae family that represents a novel host species for TULV. A cytochrome b sequence analysis confirmed the species. So far, the literature only reports on TULV in species belonging to the Arvicolinae subfamily, such as Microtus spp. and Lagurus lagurus [40,41]. However, by comparing the capture sites of those two infected specimens, it becomes apparent that the spots in Tekeli had a spatial distance of only 325 m. In this region, D. nitedula is a common mammal, mostly living on trees but also reported to hunt for edibles on the ground, since also the traps were only located on terra firma. There, it may have indirect contact with M. arvalis that builds nests in subterraneous burrows but also gathers edibles on the ground. The infection of atypical host species with orthohantavirus is designated as a spill-over infection and is reported in high incidence areas in Europe [41]. Since we identified several infected rodents in the Tekeli area and the S segment sequences derived from D. nitedula and M. arvalis are almost identical, such a spill-over event is in the scope of possibilities [42,43,44]. Nevertheless, this result implies the need for a more extensive follow-up host-study for infected small mammals in the area of the Almaty region to obtain information on the actual distribution of orthohantaviruses in this area.
To further estimate the connection of these viruses, we performed sequencing of parts of the S and L segments. Sequence similarities for the partial S segments of the clusters of South-East Kazakhstan (I) and West Kazakhstan/Samara (IV) resemble these of previous studies [41]. Furthermore, the phylogenetic analysis of the partial S segment sequences enabled the classification of TULV in a broad geographical range [43,45,46]. Our results highlight that TULV from West Kazakhstan is indeed in close evolutionary relationship with TULV described in Samara, the adjoining region in the Russian Federation (DQ061258). Almaty region (Tekeli and Rudnichniy) has its own cluster separated from all other TULV sequences for the S segment (Figure 2). Additionally, it is evident that the West Kazakhstan TULV S segment sequence is only distantly related to other Kazakhstan sequences as, for instance, from the Almaty region, a region over 2000 km apart from West Kazakhstan. Sequences from the Tekeli city and Rudnichniy village in the Almaty region shared a close relationship with previously published sequences of M. arvalis sampled in the village of Karatal and Taldykorgan city, located also in the Almaty region [22]. It is highly probably that there exist different geographic lineages of TULV in Kazakhstan transmitted by different lineages of rodents as recently shown for TULV sequences in Europe [33,40,41].
The sequence relationships identified for the S sequence analysis can also be identified in the analysis of the partial L segment sequences, where we could show that the TULV L segment sequence from West Kazakhstan region formed its own distinct geographic cluster. In general, published sequences for the L segment in this region are sparse and for the Almaty region, we describe for the first time also TULV L segment sequences, in comparison to a previous study that only analysed the S segment [22]. Sequences from Tekeli and Rudnichniy in Almaty region cluster in an individual branch in one big cluster with sequences from China and Turkey (Figure 3) [47,48]. This finding goes along with previous studies who have illustrated that genetic clustering of TULV is largely according to geographic regions [22,33].
5. Conclusions
Here, we screened 621 small mammals for their orthohantavirus infection rate. Interestingly, we only identified the relatively benign TULV species, a finding that is contrary to the expectation risen by patients with episodes of haemorrhagic fever in Kazakhstan hospitals. Knowledge on the pathogenicity of TULV and the impact of TULV-associated disease in humans is limited. Only few cases, mostly mild, were described in Europe, some of them in immunocompromised patients [10,11,12,13,49]. In certain risk groups, e.g., forest workers, a higher antibody prevalence against TULV was found in comparison to the normal population [11,12]. However, the severe cases of HFRS observed in the hospitals in West Kazakhstan are most probably not induced by an infection with TULV but rather by PUUV. The exact endemic areas for this virus in Kazakhstan remain obscure.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/v14061258/s1, Figure S1: All available amino acid S segment sequences from Kazakhstan and close geographic regions in Russia (Omsk, Samara) and China; Figure S2: All available amino acid L segment sequences from Kazakhstan and close geographic regions in Russia (Omsk, Samara) and China; Table S1: Detailed information on trapping sites of small mammals.
Author Contributions
N.T. (Nur Tukhanova), S.F., L.P., E.W. and S.E. conceived the layout of the project. N.T. (Nur Tukhanova), E.W., V.S., A.B., N.M., A.Z. and I.L. participated in the fieldwork and the preparation and analysis of collected small mammals. I.L. performed morphological determination of the captured small mammals. N.T. (Nur Tukhanova), A.S. (Anna Shin), E.W., N.T. (Nurkeldi Turebekov), K.A. and T.N. contributed tissue homogenization and RNA extraction of collected samples. N.T. (Nur Tukhanova) performed molecular biology testing and analysis. A.S. (Alexandr Shevtsov) was in charge of the sequencing. N.T. (Nur Tukhanova) wrote the draft manuscript. N.T. (Nur Tukhanova) and L.P. created the figures and tables. T.Y., G.T., L.Y., G.F., M.H. and S.F. contributed additional information and reviewed the manuscript. S.E. and L.P. supervised the project. L.P. was in charge of the revision process. All authors have read and agreed to the published version of the manuscript.
Funding
This study is supported by the German Biosecurity Program of the German Federal Foreign Office. The authors also express gratitude to the German Federal Ministry for Economic Cooperation and Development (BMZ) and the German Academic Exchange Services (DAAD) through the CIH LMU—Center for International Health, Ludwig-Maximilian University, Munich, Germany.
Institutional Review Board Statement
Rodent trapping occurred after ethical approval of Kazakhstan local ethics committee at National Scientific Center Especially Dangerous Infectious in Almaty, Kazakhstan (protocol #4, 08.01.18) and the ethical committee of the Ludwig-Maximilian University in Munich, Germany (18-631).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used and/or analysed during the current study are available from the corresponding author on reasonable request. All determined sequences were uploaded to GenBank and are accessible as OL677520, OL677521, OL677522, OL677523, OL677524, OL677525, OL677526, OL677527, OL677528, OL677529, OL677530, OL677531, OL677532, OL677533, OL677534, ON239876, ON239877, ON239878, ON239879, ON239880, ON239881, ON239882, ON239883, ON239884, ON239885, ON239886, ON239887, ON239888, ON239889, ON239890, ON513437, ON513438, and ON513439.
Acknowledgments
We thank the staff members of the Taldykorgan and Oral Antiplague Stations for excellent assistance in gathering samples.
Conflicts of Interest
The authors declare no conflict of interest. The authors declare that there is no financial or personal relationship with other people or organizations that could inappropriately influence the work. Opinions, interpretations, conclusions, and recommendations are those of the authors and are not necessarily endorsed by Bundeswehr Joint Medical Service or any other governmental institutions.
References
- Vaheri, A.; Henttonen, H.; Voutilainen, L.; Mustonen, J.; Sironen, T.; Vapalahti, O. Hantavirus infections in Europe and their impact on public health. Rev. Med. Virol. 2013, 23, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current Classification and Future Perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Krautkrämer, E. Hantaviruses—Infections, Epidemiology and Hosts. In Zoonooses: Infections Affecting Men and Animals—A Focus on Public Health Aspects; Springer: Berlin/Heidelberg, Germany, 2015; pp. 749–783. [Google Scholar]
- Schlegel, M.; Jacob, J.; Krüger, D.H.; Rang, A.; Ulrich, R.G. Hantavirus Emergence in Rodents, Insectivores and Bats. In The Role of Animals in Emerging Viral Diseases; Academic Press: Cambridge, MA, USA, 2014; pp. 235–292. [Google Scholar] [CrossRef]
- Avšič-Županc, T.; Saksida, A.; Korva, M. Hantavirus infections. Clin. Microbiol. Infect. 2019, 21S, e6–e16. [Google Scholar] [CrossRef] [PubMed]
- Vaheri, A.; Henttonen, H.; Mustonen, J. Hantavirus Research in Finland: Highlights and Perspectives. Viruses 2021, 13, 1452. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Numan, A.; Sharma, D.; Shukla, R.; Alexander, A.; Jain, G.K.; Ahmad, F.J.; Kesharwani, P. Epidemiology, virology and clinical aspects of hantavirus infections: An overview. Int. J. Environ. Health Res. 2021, 22, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.G.; Anjum, F. Hemorrhagic Fever Renal Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Ermonval, M.; Baychelier, F.; Tordo, N. What Do We Know about How Hantaviruses Interact with Their Different Hosts? Viruses 2016, 8, 223. [Google Scholar] [CrossRef]
- Klempa, B.; Meisel, H.; Räth, S.; Bartel, J.; Ulrich, R.; Krüger, D.H. Occurrence of Renal and Pulmonary Syndrome in a Region of Northeast Germany Where Tula Hantavirus Circulates. J. Clin. Microbiol. 2003, 41, 4894–4897. [Google Scholar] [CrossRef]
- Mertens, M.; Hofmann, J.; Petraityte-Burneikiene, R.; Ziller, M.; Sasnauskas, K.; Friedrich, R.; Niederstrasser, O.; Krüger, D.H.; Groschup, M.H.; Petri, E.; et al. Seroprevalence study in forestry workers of a non-endemic region in eastern Germany reveals infections by Tula and Dobrava-Belgrade hantaviruses. Med. Microbiol. Immunol. 2011, 200, 263–268. [Google Scholar] [CrossRef]
- Zelená, H.; Mrázek, J.; Kuhn, T. Tula Hantavirus Infection in Immunocompromised Host, Czech Republic. Emerg. Infect. Dis. 2013, 19, 1873–1876. [Google Scholar] [CrossRef]
- Hofmann, J.; Kramer, S.; Herrlinger, K.R.; Jeske, K.; Kuhns, M.; Weiss, S.; Ulrich, R.G.; Krüger, D.H. Tula Virus as Causative Agent of Hantavirus Disease in Immunocompetent Person, Germany. Emerg. Infect. Dis. 2021, 27, 1234–1237. [Google Scholar] [CrossRef]
- Akram, S.M.; Mangat, R.; Huang, B. Hantavirus Cardiopulmonary Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Burek, K.A.; Rossi, C.A.; Leduc, J.W.; Yuill, T.M. Serologic and Virologic Evidence of a Prospect Hill-like Hantavirus in Wisconsin and Minnesota. Am. J. Trop. Med. Hyg. 1994, 51, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Peintner, L.; Wagner, E.; Shin, A.; Tukhanova, N.; Turebekov, N.; Abdiyeva, K.; Spaiser, O.; Serebrennikova, Y.; Tintrup, E.; Dmitrovskiy, A.; et al. Eight Years of Collaboration on Biosafety and Biosecurity Issues Between Kazakhstan and Germany as Part of the German Biosecurity Programme and the G7 Global Partnership Against the Spread of Weapons and Materials of Mass Destruction. Front. Public Health 2021, 9, 1102. [Google Scholar] [CrossRef] [PubMed]
- Aikimbayev, M. Atlas of Bacterial and Virus Zoonotic Infections Distribution in Kazakhstan; Kazakh Scientific Centre for Quarantine and Zoonotic Diseases under CSSES of the MPH of the Republic of Kazakhstan: Almaty, Kazakhstan, 2010. [Google Scholar]
- Abdiyeva, K.; Turebekov, N.; Dmitrovsky, A.; Tukhanova, N.; Shin, A.; Yeraliyeva, L.; Heinrich, N.; Hoelscher, M.; Yegemberdiyeva, R.; Shapiyeva, Z.; et al. Seroepidemiological and molecular investigations of infections with Crimean–Congo haemorrhagic fever virus in Kazakhstan. Int. J. Infect. Dis. 2019, 78, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Bezverhnii, A. Comprehensive Epidemiological Investigation of Zoonotic Infections in the Dzungarian Alatau. Ph.D. Thesis, Kazakhstan National University, Almaty, Kazakhstan, 1995. [Google Scholar]
- Sutyagin, V. Hemorrhagic fever with renal syndrome in Dzungarian Alatau. Quar. Zoonotic Infect. Kaz. 2010, 1, 120–121. [Google Scholar]
- Sutyagin, V.; Belyaev, A.; Kim, I.; Berdibekov, A. Distribution of hemorrhagic fever with renal syndrome in Almaty region Dzungarian Alatau. Environ. Public Health 2017, 3, 56–58. [Google Scholar]
- Plyusnina, A.; Laakkonen, J.; Niemimaa, J.; Henttonen, H.; Plyusnin, A. New Genetic Lineage of Tula Hantavirus in Microtus arvalis obscurus in Eastern Kazakhstan. Open Virol. J. 2008, 2, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Tukhanova, N.; Shin, A.; Abdiyeva, K.; Turebekov, N.; Yeraliyeva, L.; Yegemberdiyeva, R.; Shapiyeva, Z.; Froeschl, G.; Hoelscher, M.; Wagner, E.; et al. Serological investigation of orthohantaviruses in patients with fever of unknown origin in Kazakhstan. Zoonoses Public Health 2020, 67, 271–279. [Google Scholar] [CrossRef]
- Grazhdanov, A.; Zakharov, A.; Biryukov, A. First cases of Hemorrhagic fever with renal syndrome in Kazakhstan. J. Quar. Zoonotic Infect. Kaz. 2001, 3, 94–98. [Google Scholar]
- Bidashko, F.; Grazhdanov, A.; Rakhmankulov, R. Some aspects of the epizootology of the Ural-Ilek foci of Hemorrhagic fever with renal syndrome. J. Quar. Zoonotic Infect. Kaz. 2004, 1, 96–104. [Google Scholar]
- Grazhdanov, A.K.; Ayazbaev, T.Z.; Toporkov, A.V.; Bidashko, F.G.; Zakharov, A.V.; Belonozhkina, L.; Pak, M.V.; Andryushchenko, A.V. Concerning the Allocation of Emerging Natural Foci of the Currently Important Infectious Diseases in the West of Kazakhstan. Probl. Part. Danger. Infect. 2014, 3, 20–24. [Google Scholar] [CrossRef]
- Gromov, M.; Erbajeva, M. The Mammals of the Russia and Adjacent Territories (Lagomorphs and Rodents); Russian Academy of Sciences Zoological Institute: Saint Petersburg, Russia, 1995. [Google Scholar]
- Scharninghausen, J.J.; Meyer, H.; Pfeffer, M.; Davis, D.S.; Honeycutt, R.L. Genetic Evidence of Dobrava Virus in Apodemus agrarius in Hungary. Emerg. Infect. Dis. 1999, 5, 468–470. [Google Scholar] [CrossRef] [PubMed]
- Essbauer, S.; Schmidt, J.; Conraths, F.J.; Friedrich, R.; Koch, J.; Hautmann, W.; Pfeffer, M.; Wölfel, R.; Finke, J.; Dobler, G.; et al. A new Puumala hantavirus subtype in rodents associated with an outbreak of Nephropathia epidemica in South-East Germany in 2004. Epidemiol. Infect. 2006, 134, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Mossbrugger, I.; Felder, E.; Gramsamer, B.; Wölfel, R. EvaGreen based real-time RT-PCR assay for broad-range detection of hantaviruses in the field. J. Clin. Virol. 2013, 58, 334–335. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, M.; Ali, H.S.; Stieger, N.; Groschup, M.H.; Wolf, R.; Ulrich, R.G. Molecular identification of small mammal species using novel cytochrome B gene-derived degenerated primers. Biochem. Genet. 2012, 50, 440–447. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Schmidt, S.; Saxenhofer, M.; Drewes, S.; Schlegel, M.; Wanka, K.M.; Frank, R.; Klimpel, S.; Von Blanckenhagen, F.; Maaz, D.; Herden, C.; et al. High genetic structuring of Tula hantavirus. Arch. Virol. 2016, 161, 1135–1149. [Google Scholar] [CrossRef]
- Maas, M.; de Vries, A.; van Roon, A.; Takumi, K.; van der Giessen, J.; Rockx, B. High Prevalence of Tula Hantavirus in Common Voles in The Netherlands. Vector Borne Zoonotic Dis. 2017, 17, 200–205. [Google Scholar] [CrossRef]
- Scharninghausen, J.J.; Pfeffer, M.; Meyer, H.; Davis, D.S.; Honeycutt, R.L.; Faulde, M. Genetic Evidence for Tula Virus in Microtus arvalis and Microtus agrestis Populations in Croatia. Vector-Borne Zoonotic Dis. 2002, 2, 19–27. [Google Scholar] [CrossRef]
- Bekmukhambetov, S.K. Experience of diagnosis and treatment of epidemic hemorrhagic fever in Kazakhstan. J. Med. 2012, 4, 58–66. (In Russian) [Google Scholar]
- Zakharov, A.V.; Grazhdanov, A.K.; Zakharov, V.M.; Nazhimova, G.S. Clinical manifestations of acute renal failure in hemorrhagic fever with renal syndrome. J. Quar. Zoonotic Infect. Kaz. 2010, 1–2, 18–22. (In Russian) [Google Scholar]
- Kariwa, H.; Tkachenko, E.A.; Morozov, V.G.; Seto, T.; Tanikawa, Y.; Kolominov, S.I.; Belov, S.N.; Nakamura, I.; Hashimoto, N.; Balakiev, A.E.; et al. Epidemiological Study of Hantavirus Infection in the Samara Region of European Russia. J. Vet. Med. Sci. 2009, 71, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Rabenau, H.F.; Kessler, H.H.; Kortenbusch, M.; Steinhorst, A.; Raggam, R.B.; Berger, A. Verification and validation of diagnostic laboratory tests in clinical virology. J. Clin. Virol. 2007, 40, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-W.; Gligic, A.; Yanagihara, R. Identification of Tula hantavirus in Pitymys subterraneus captured in the Cacak region of Serbia-Yugoslavia. Int. J. Infect. Dis. 2002, 6, 31–36. [Google Scholar] [CrossRef]
- Schmidt-Chanasit, J.; Essbauer, S.; Petraityte, R.; Yoshimatsu, K.; Tackmann, K.; Conraths, F.J.; Sasnauskas, K.; Arikawa, J.; Thomas, A.; Pfeffer, M.; et al. Extensive Host Sharing of Central European Tula Virus. J. Virol. 2010, 84, 459–474. [Google Scholar] [CrossRef]
- Saxenhofer, M.; Labutin, A.; White, T.A.; Heckel, G. Host genetic factors associated with the range limit of a European hantavirus. Mol. Ecol. 2021, 31, 252–265. [Google Scholar] [CrossRef]
- Saxenhofer, M.; Schmidt, S.; Ulrich, R.G.; Heckel, G. Secondary contact between diverged host lineages entails ecological speciation in a European hantavirus. PLoS Biol. 2019, 10, e3000142. [Google Scholar] [CrossRef]
- Schmidt, S.; Reil, D.; Jeske, K.; Drewes, S. Spatial and Temporal Dynamics and Molecular Evolution of Tula orthohantavirus in German Vole Populations. Viruses 2021, 13, 1132. [Google Scholar] [CrossRef]
- Saxenhofer, M.; de Melo, W.V.; Ulrich, R.G.; Heckel, G. Revised time scales of RNA virus evolution based on spatial information. Proc. R. Soc. B 2017, 284, 20170857. [Google Scholar] [CrossRef]
- Hiltbrunner, M.; Heckel, G. Assessing Genome-Wide Diversity in European Hantaviruses through Sequence Capture from Natural Host Samples. Viruses 2020, 12, 749. [Google Scholar] [CrossRef]
- Chen, J.-T.; Qin, J.; Li, K.; Xu, Q.-Y.; Wang, X.-P.; Plyusnin, A.; Hou, W.; Zhang, Y.-Z. Identification and characterization of a novel subtype of Tula virus in Microtus arvalis obscurus voles sampled from Xinjiang, China. Infect. Genet. Evol. 2019, 75, 104012. [Google Scholar] [CrossRef]
- Polat, C.; Ergünay, K.; Irmak, S.; Erdin, M.; Brinkmann, A.; Çetintaş, O.; Çoğal, M.; Sözen, M.; Matur, F.; Nitsche, A.; et al. A novel genetic lineage of Tula orthohantavirus in Altai voles (Microtus obscurus) from Turkey. Infect. Genet. Evol. 2019, 67, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Schultze, D.; Lundkvist, A.; Blauenstein, U.; Heyman, P. Tula virus infection associated with fever and exanthema after a wild rodent bite. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 304–306. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).