
Eye Infection with SARS-CoV-2 as a Route to Systemic Immunization?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Groups
2.2. Virus Source, Handling and Quantity
2.3. Ante-Mortem Assessment
2.3.1. Clinical Scoring
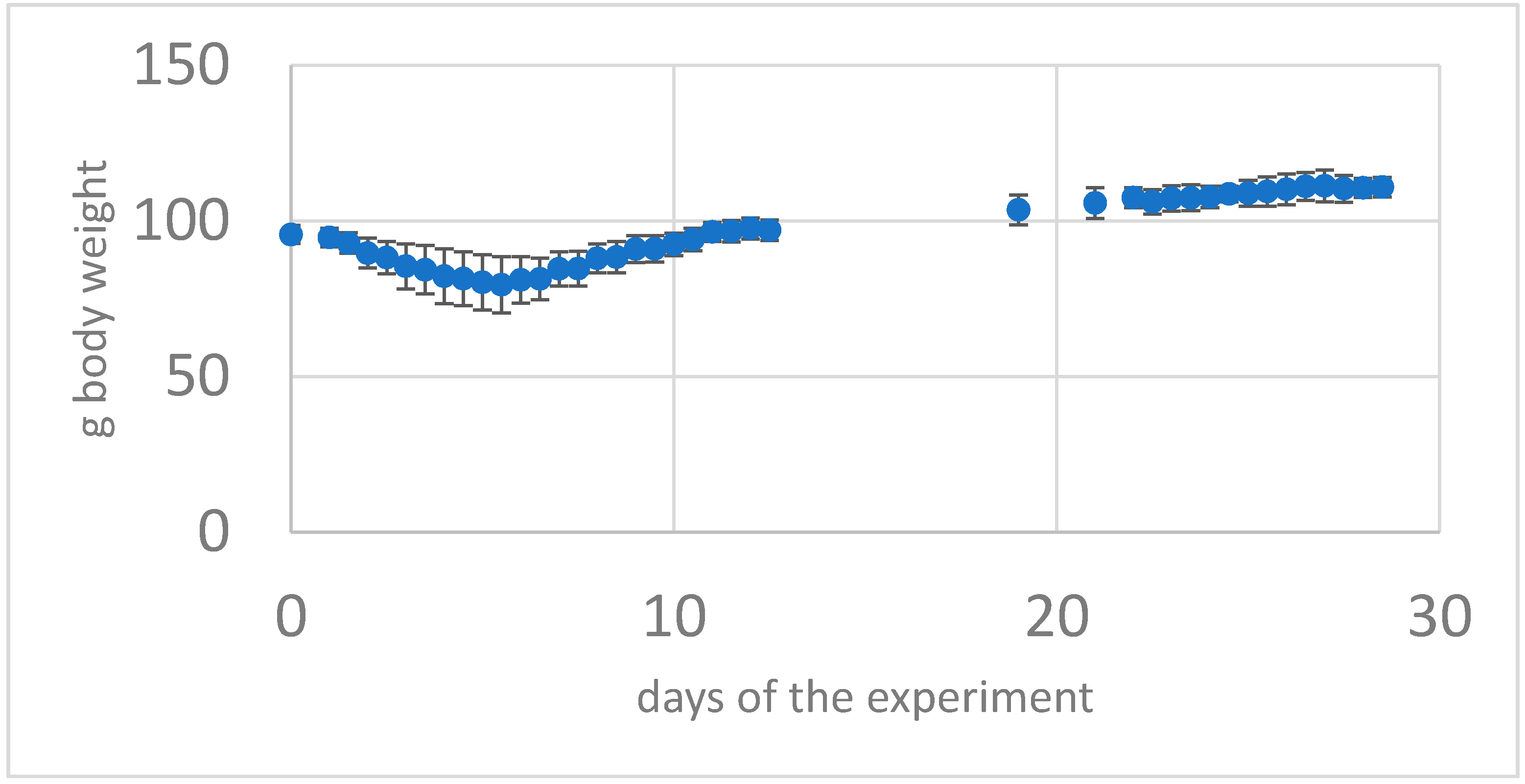
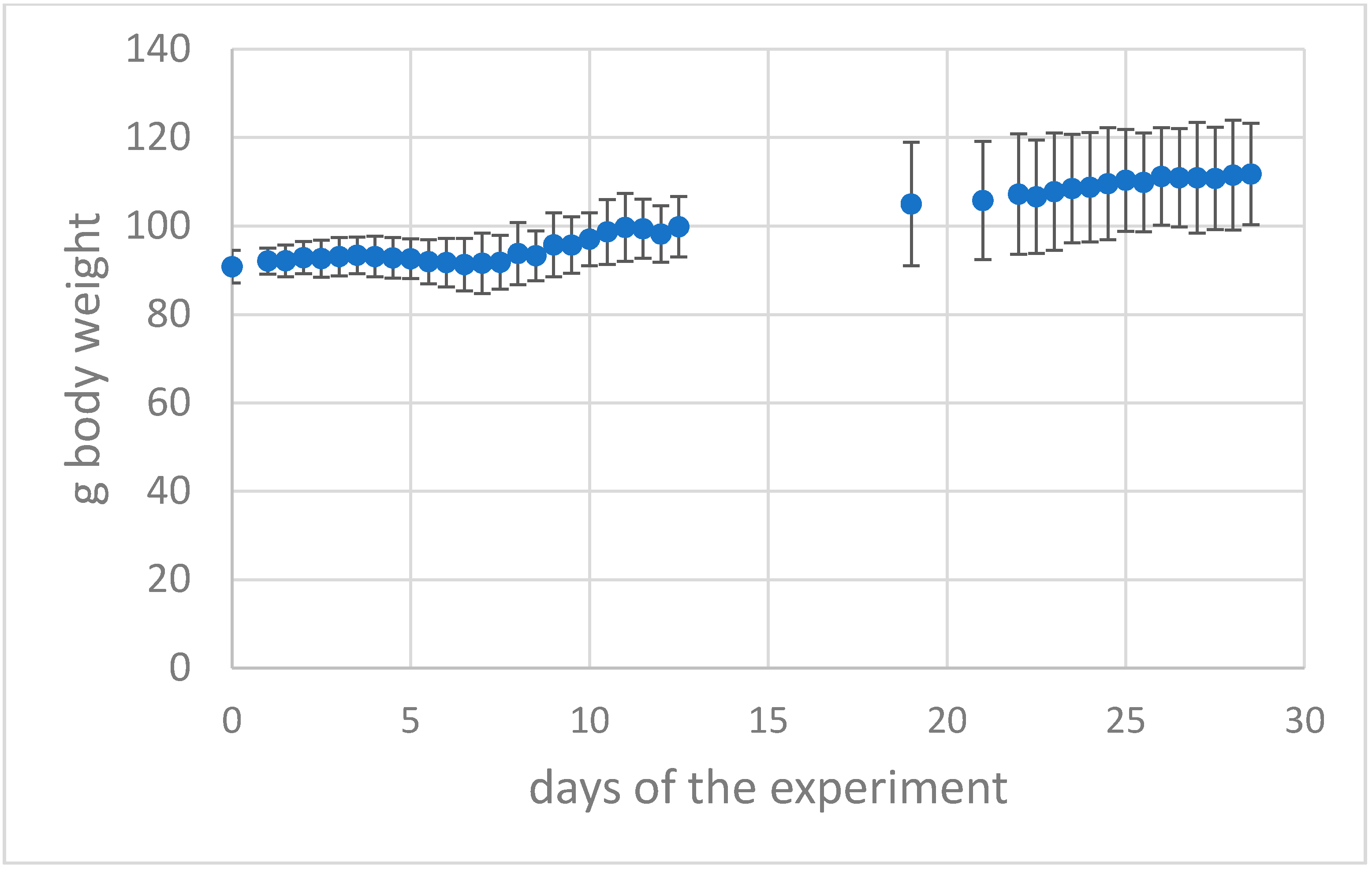
2.3.2. Weight Gain/Deficit
2.4. Plethysmography
2.4.1. Overview
2.4.2. Analysis of Individual Breathing Patterns
2.4.3. Animal Welfare
2.5. Postmortem Assessment
2.5.1. Autopsy
2.5.2. Histopathology and Immunohistochemistry
2.5.3. Test for Infectious Viruses in the Lungs
- Seeding of culture plates with host cells. Seed 7.5 × 103 cells per 100 µL in growth media (DMEM/FBS10%) in each well of 96-well plates. Gently swirl the plates so that the cells are evenly distributed. Grow the cells overnight. The next day, check under a light microscope to see if the cells are evenly distributed and have reached a degree of confluence of about 75%.
- Serial dilutions of the lung homogenate. Prepare a series of 1:10 dilutions of the lung homogenates to be titrated. The first tube is filled with 2.0 mL of infection medium (DMEM/FBS2%), and another six tubes are filled in series with 1.8 mL of infection medium. Prepare the lung homogenate with the vortex mixer and then transfer 200 μL of the suspension to the first tube. Briefly mix the first tube and prepare a 1:100 dilution by transferring 200 μL of the first to the second tube. Then transfer 200 μL of the 1:100 dilution to the next tube in the series. Repeat the procedure to make a serial 1:10 dilution of the lung suspension, e.g., from 10–1 to 10–7.
- Pipetting into 96-well plates. The successive 1:10 dilutions are pipetted onto the Vero cell monolayers and labeled. Provide four negative wells on each plate (which will not be in contact with the dilutions of lung homogenates). Carefully remove the growth medium from each well. Then add 100 µL of the lung dilutions per well and infect 4 wells per dilution by sweeping backwards through the dilutions. Subsequently, the cells adsorb the virus for approximately two hours at 37 °C, then 100 µL of infection medium is added to each well and the plates are placed back in the CO2 incubator at 37 °C or to monitor CPE for five days.
- Visualization and calculation of TCID50. The endpoint is reached when the CPE value appears the same on three separate readings per dilution. The titer is calculated according to the method of Reed and Muench. A titer expressed as 103 TCID/mL 50 in five days in the VeroE6 cell line can be translated as follows: 1 mL of lung homogenate diluted 1:1000 infects 50% of the cells in five days using the Vero-E6 cell line.
2.5.4. Determination of Total Viral Genomic Load of the Lung
2.5.5. Quantification of the Negative Strand
2.5.6. Evaluation of Neutralizing Antibodies in Serum
3. Results
- epi-ocular infection with SARS-CoV-2 induces immunization with or without disease in the animal; and
- general immunity to SARS-CoV-2 is developed after primary epi-ocular infection that protects against re-infection.
3.1. Clinical Follow-Up
3.1.1. First Intranasal and Single Epi-Ocular Viral Exposure
3.1.2. Epi-Ocular Virus Exposure
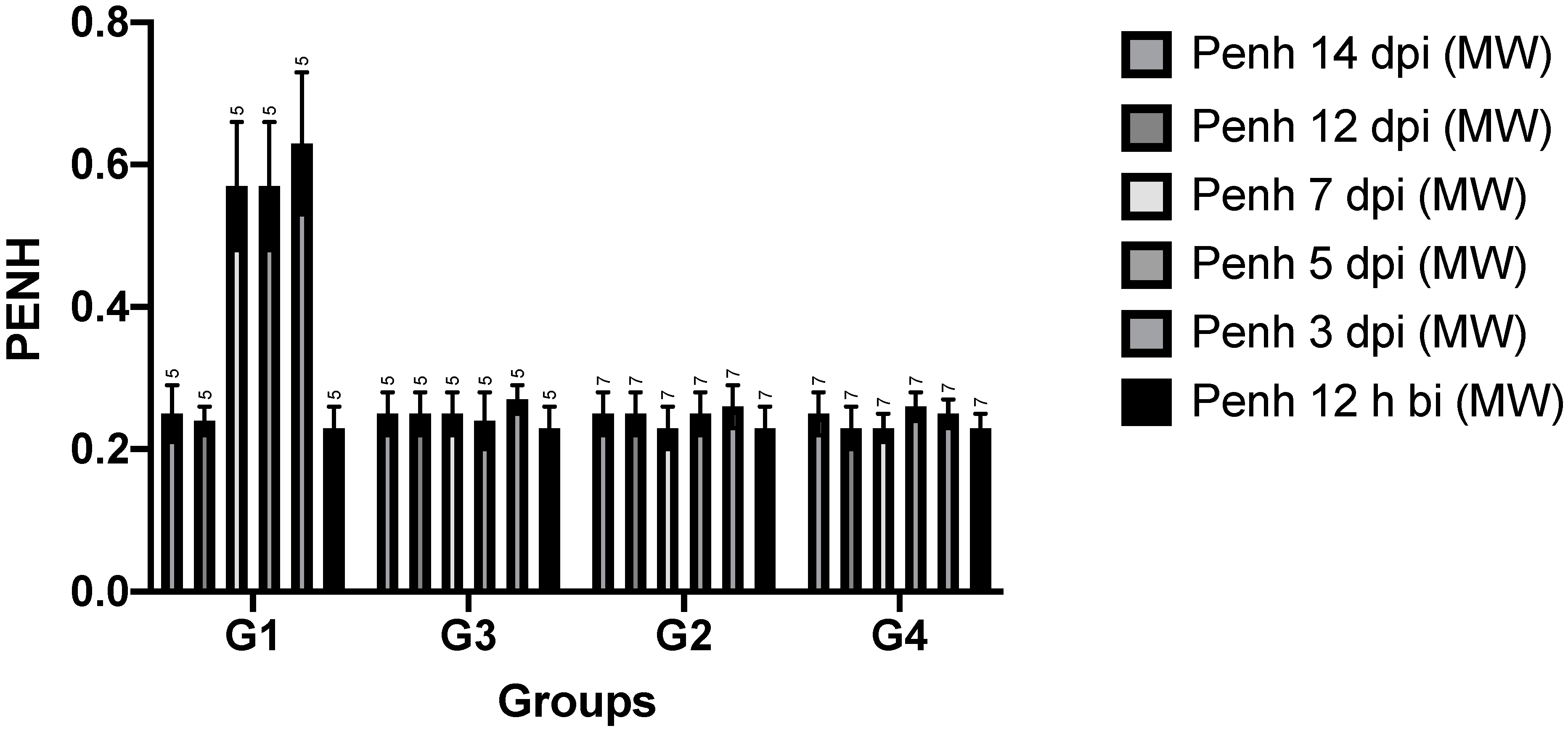
3.1.3. Whole-Body Plethysmography

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| G1-12 h Before Intransal Infection | G2-12 h Before Intransasal Re-Infection | G3-12 h Before Epiocular Infection | G4-12 h Before Intranasal Re-Infection | |
|---|---|---|---|---|
| D3pi | p 0.0001 | ns | p 0.0201 | ns |
| D5pi | p 0.0001 | ns | Ns | p 0.0102 |
| D7pi | p 0.0001 | ns | ns | ns |
| D12pi | ns | ns | ns | ns |
3.2. Time Points
3.3. Volumes, Mean and Peak Flow Rates
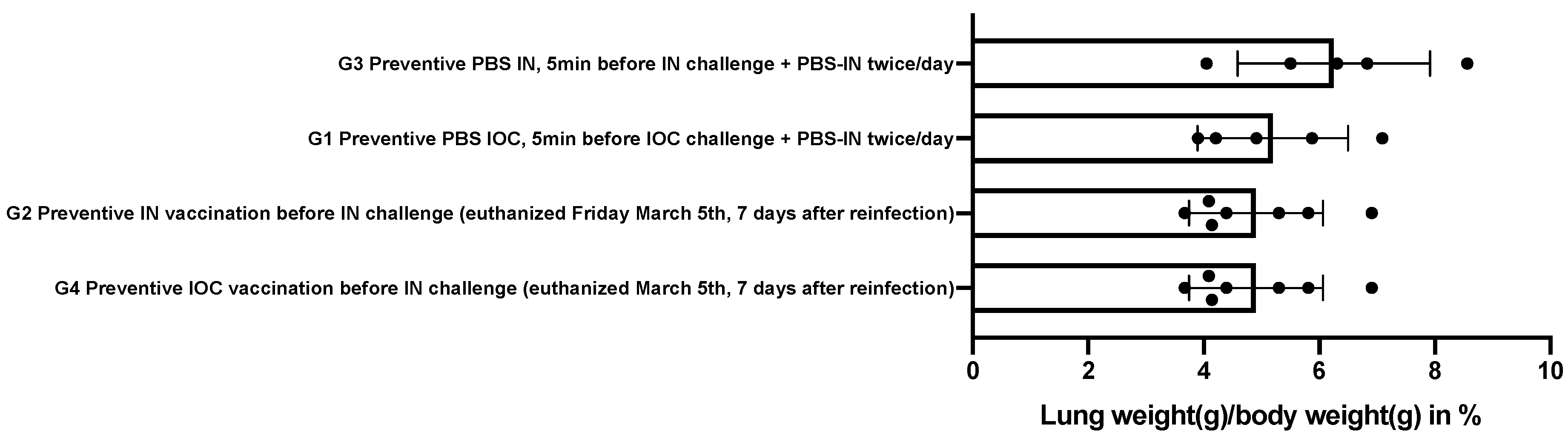
3.4. Autopsy
3.5. Histology
3.6. Viral Load in the Lungs
3.7. Neutralizing Titers
4. Discussion
- if no vaccine can be found, the limited infection of one eye will yield a weak disease with full immunity, comparable to recovery.
- if a targeted vaccination via the eyes proves successful, the amount of vaccine administered can be reduced. In contrast to current vaccines administered by intramuscular injection, we expect to achieve not only humoral but also mucosal immunization [9].
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/Overview (accessed on 5 June 2022).
- da Silva Torres, M.K.; Bichara, C.D.A.; Vallinoto, M.C.; Queiroz, M.A.F.; Vallinoto, I.M.V.C.; Dos Santos, E.J.M.; de Carvalho, C.A.M.; Vallinoto, A.C.R. The complexity of SARS-CoV-2 infection and the COVID-19 pandemic. Front. Microbiol. 2022, 13, 789882. [Google Scholar] [CrossRef]
- Richardson, J.R.; Götz, R.; Mayr, V.; Lohse, M.J.; Holthoff, H.-P.; Ungerer, M. SARS-CoV2 wild type and mutant specific humoral and T cell immunity is superior after vaccination than after natural infection. PLoS ONE 2022, 17, e0266701. [Google Scholar] [CrossRef] [PubMed]
- Gebert, A.; Pabst, R. M cells at locations outside the gut. Semin. Immunol. 1999, 11, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Jonas, R.A.; Ung, L.; Rajaiya, J.; Chodosh, J. Mystery eye: Human adenovirus and the enigma of epidemic keratoconjunctivitis. Prog. Retin. Eye Res. 2020, 76, 100826. [Google Scholar] [CrossRef] [PubMed]
- Alrawashdeh, H.M.; Al Zubi, K.; Abdulmannan, D.M.; Al-Habahbeh, O.; Abu-Ismail, L. Conjunctivitis as the only sign and symptom of COVID-19: A case report and review of literature. Qatar Med. J. 2021, 2021, 31. [Google Scholar]
- Sirakaya, E.; Sahiner, M.; Sirakaya, H.A. A patient with bilateral conjunctivitis positive for SARS-CoV-2 RNA in a conjunctival sample. Cornea 2021, 40, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Zerti, D.; Genitti, G.; Vicentini, M.T.; Baccante, M.; Flati, V.; Maccarone, R. Characterization of SARS-CoV-2 Entry Factors’ Expression in Corneal and Limbal Tissues of Adult Human Donors Aged from 58 to 85. J. Ocul. Pharmacol. Ther. 2022, 38, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.Y.; Han, S.J.; Cha, H.-R.; Seo, S.-U.; Song, J.-H.; Chung, S.-H.; Kweon, M.-N. Eye mucosa: An efficient vaccine delivery route for inducing protective immunity. J. Immunol. 2010, 185, 3610–3619. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-D.; Han, S.J.; Byun, Y.-H.; Yoon, S.C.; Choi, K.S.; Seong, B.L.; Seo, K.Y. Inactivated eyedrop influenza vaccine adjuvanted with poly (I:C) Is safe and effective for inducing protective systemic and mucosal immunity. PLoS ONE 2015, 10, e0137608. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Bao, L.; Gao, H.; Xiang, Z.; Qu, Y.; Song, Z.; Gong, S.; Liu, J.; Liu, J.; Yu, P.; et al. Ocular conjunctival inoculation of SARS-CoV-2 can cause mild COVID-19 in rhesus macaques. Nat. Commun. 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Selvaraj, P.; Lien, C.Z.; Liu, S.; Stauft, C.B.; Nunez, I.A.; Hernandez, M.; Nimako, E.; Ortega, M.A.; Starost, M.F.; Dennis, J.U.; et al. SARS-CoV-2 infection induces protective immunity and limits transmission in Syrian hamsters. Life Sci. Alliance 2021, 4, e202000886. [Google Scholar] [CrossRef] [PubMed]
- Misset, B.; Hoste, E.; Donneau, A.-F.; Grimaldi, D.; Meyfroidt, G.; Moutschen, M.; Compernolle, V.; Gothot, A.; Desmecht, D.; Garigliany, M.; et al. A multicenter randomized trial to assess the efficacy of CONvalescent plasma therapy in patients with Invasive COVID-19 and acute respiratory failure treated with mechanical ventilation: The CONFIDENT trial protocol. BMC Pulm. Med. 2020, 20, 317. [Google Scholar] [CrossRef] [PubMed]
- Hamelmann, E.; Schwarze, J.; Takeda, K.; Oshiba, A.; Larsen, G.L.; Irvin, C.G.; Gelfand, E.W. Noninvasive Measurement of Airway Responsiveness in Allergic Mice Using Barometric Plethysmography. Am. J. Respir. Crit. Care Med. 1997, 156, 766–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarze, J.; Hamelmann, E.; Bradley, K.L.; Takeda, K.; Gelfand, E.W. Respiratory syncytial virus infection results in airway hyperresponsiveness and enhanced airway sensitization to allergen. J. Clin. Investig. 1997, 100, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, I.A.; Lien, C.Z.; Selvaraj, P.; Stauft, C.B.; Liu, S.; Starost, M.F.; Wang, T.T. SARS-CoV-2 B.1.1.7 Infection of Syrian Hamster Does Not Cause More Severe Disease, and Naturally Acquired Immunity Confers Protection. mSphere 2021, 6, e0050721. [Google Scholar] [CrossRef] [PubMed]
- Salz, A.; Acharya, M.; Hofmann, N.; Wittmershaus, I.; Sangwan, V.; Börgel, M.; Mathur, U. Risk of SARS-CoV-2 virus transmission from donor corneal tissue: A review. Indian J. Ophthalmol. 2021, 69, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- Brdovčak, M.C.; Materljan, J.; Šustić, M.; Ravlić, S.; Ružić, T.; Lisnić, B.; Miklić, K.; Brizić, I.; Matešić, M.P.; Lisnić, V.J.; et al. ChAdOx1-S adenoviral vector vaccine applied intranasally elicits superior mucosal immunity compared to the intramuscular route of vaccination. Eur. J. Immunol. 2022, 52, 936–945. [Google Scholar] [CrossRef] [PubMed]





| Group/Way of Infection IN = Intranasal IOC = Epiocular | G4 | G2 | G3 | G1 |
|---|---|---|---|---|
| G4 Preventive /IOC infection | X | ns | p 0.0013 | p 0.0013 |
| G2 Preventive /IN infection | X | p 0.0051 | p 0.0025 | |
| G3 preventive PBS /IOC | X | ns | ||
| G1 Preventive PBS /IN | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schrage, N.; Blomet, J.; Holzer, F.; Tromme, A.; Ectors, F.; Desmecht, D. Eye Infection with SARS-CoV-2 as a Route to Systemic Immunization? Viruses 2022, 14, 1447. https://doi.org/10.3390/v14071447
Schrage N, Blomet J, Holzer F, Tromme A, Ectors F, Desmecht D. Eye Infection with SARS-CoV-2 as a Route to Systemic Immunization? Viruses. 2022; 14(7):1447. https://doi.org/10.3390/v14071447
Chicago/Turabian StyleSchrage, Norbert, Joel Blomet, Frank Holzer, A. Tromme, F. Ectors, and Daniel Desmecht. 2022. "Eye Infection with SARS-CoV-2 as a Route to Systemic Immunization?" Viruses 14, no. 7: 1447. https://doi.org/10.3390/v14071447
APA StyleSchrage, N., Blomet, J., Holzer, F., Tromme, A., Ectors, F., & Desmecht, D. (2022). Eye Infection with SARS-CoV-2 as a Route to Systemic Immunization? Viruses, 14(7), 1447. https://doi.org/10.3390/v14071447