
Using Self-Assembling ADDomer Platform to Display B and T Epitopes of Type O Foot-and-Mouth Disease Virus

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Amino Acid Sequence Alignment, 3D Structure Displaying, and Simulation
2.2. Construction of Recombinant Baculoviruses
2.3. Generation of VLPs
2.4. Identification of the Recombinant Proteins by Western Blotting and QE Mass Spectrometry
2.5. Characterization of VLPs Using Transmission Electron Microscopy (TEM)
2.6. Animal Immunization and Sample Collection
2.7. Evaluation of FMDV-Specific Antibody by Indirect ELISA
2.8. Detection of T Lymphocyte Level by Flow Cytometry
2.9. IL-2, IL-4, and IFN-γ Release Assays
2.10. Statistical Analysis
3. Results
3.1. Three-Dimensional Structural Homology Modeling of Recombinant Proteins
3.2. Expression and Characterization of Recombinant Proteins
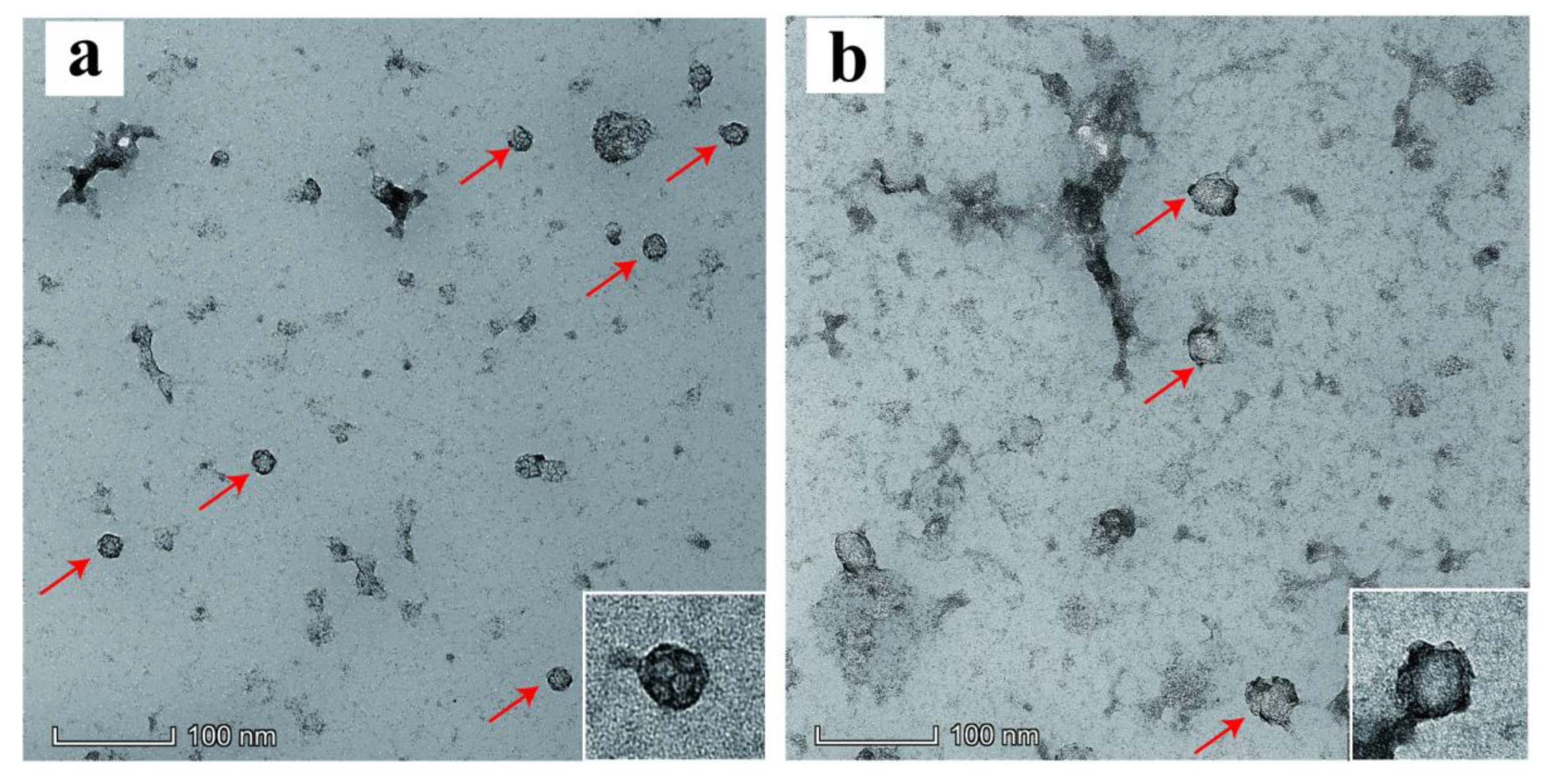
3.3. Transmission Electron Microscopy Analysis
3.4. FMDV-Specific Humoral Immune Responses
3.5. T Lymphocyte Level Detected by Flow Cytometry
3.6. The Levels of Cytokines in Sera from Mice Detected by ELISA
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gu, Y.X.; Gao, Z.L.; Zhou, J.H.; Zhang, J.; Liu, Y.S. Establishment and evaluation of stable cell lines inhibiting foot-and-mouth disease virus by RNA interference. BioMed Res. Int. 2014, 2014, 109428. [Google Scholar] [CrossRef] [PubMed]
- Horsington, J.; Nfon, C.; Bittner, H.; Durr, P.A.; Singanallur, N.; Alexandersen, S.; Vosloo, W. The protective capacity of high payload FMDV A22 IRQ vaccine in sheep against direct-contact challenge with a heterologous, contemporary FMDV A strain from South East Asia. PLoS ONE 2018, 13, e195302. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, Z.; Du, X.; Cao, W.; Yang, F.; Zhang, X.; Feng, H.; Li, D.; Zhang, K.; Liu, X.; et al. Foot-and-mouth disease virus induces lysosomal degradation of host protein kinase PKR by 3C proteinase to facilitate virus replication. Virology 2017, 509, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Jones, K.; Mahapatra, M.; Upadhyaya, S.; Paton, D.J.; Babu, A.; Hutchings, G.; Parida, S. Genetic and antigenic characterization of serotype O FMD viruses from East Africa for the selection of suitable vaccine strain. Vaccine 2017, 35, 6842–6849. [Google Scholar] [CrossRef]
- Liu, X.; Lv, J.; Fang, Y.; Zhou, P.; Lu, Y.; Pan, L.; Zhang, Z.; Ma, J.; Zhang, Y.; Wang, Y. Expression and Immunogenicity of Two Recombinant Fusion Proteins Comprising Foot-and-Mouth Disease Virus Structural Protein VP1 and DC-SIGN-Binding Glycoproteins. BioMed Res. Int. 2017, 2017, 7658970. [Google Scholar] [CrossRef]
- Chen, T.H.; Hu, C.C.; Lee, C.W.; Feng, Y.M.; Lin, N.S.; Hsu, Y.H. Stable Display of Artificially Long Foreign Antigens on Chimeric Bamboo mosaic virus Particles. Viruses 2021, 13, 572. [Google Scholar] [CrossRef]
- Lei, Y.; Shao, J.; Zhao, F.; Li, Y.; Lei, C.; Ma, F.; Chang, H.; Zhang, Y. Artificially designed hepatitis B virus core particles composed of multiple epitopes of type A and O foot-and-mouth disease virus as a bivalent vaccine candidate. J. Med. Virol. 2019, 91, 2142–2152. [Google Scholar] [CrossRef]
- Fang, M.; Li, J.; Wang, H.; Yang, M.; Zhang, Y.; Zhou, L.; Wei, H.; Yang, G.; Yu, Y.; Wei, X.; et al. Correlation between efficacy and structure of recombinant epitope vaccines against bovine type O foot and mouth disease virus. Biotechnol. Lett. 2012, 34, 839–847. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, K.N.; Kim, S.M.; Lee, H.S.; Ko, Y.J.; Tark, D.S.; Shin, Y.K.; Seo, M.G.; Kim, B. Reemergence of foot-and-mouth disease, South Korea, 2000–2011. Emerg. Infect. Dis. 2014, 20, 2158–2161. [Google Scholar] [CrossRef]
- Singh, R.K.; Sharma, G.K.; Mahajan, S.; Dhama, K.; Basagoudanavar, S.H.; Hosamani, M.; Sreenivasa, B.P.; Chaicumpa, W.; Gupta, V.K.; Sanyal, A. Foot-and-Mouth Disease Virus: Immunobiology, Advances in Vaccines and Vaccination Strategies Addressing Vaccine Failures-An Indian Perspective. Vaccines 2019, 7, 90. [Google Scholar] [CrossRef]
- Cao, Y.; Lu, Z.; Liu, Z. Foot-and-mouth disease vaccines: Progress and problems. Expert Rev. Vaccines 2016, 15, 783–789. [Google Scholar] [CrossRef]
- Ruhl, J.; Leung, C.S.; Munz, C. Vaccination against the Epstein-Barr virus. Cell Mol. Life Sci. 2020, 77, 4315–4324. [Google Scholar] [CrossRef]
- Chen, Q. Plant-made vaccines against West Nile virus are potent, safe, and economically feasible. Biotechnol. J. 2015, 10, 671–680. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, S.H.; Kim, A.R.; Quan, F.S. Virus-Like Nanoparticle Vaccine Confers Protection against Toxoplasma gondii. PLoS ONE 2016, 11, e161231. [Google Scholar] [CrossRef]
- Brune, K.D.; Howarth, M. New Routes and Opportunities for Modular Construction of Particulate Vaccines: Stick, Click, and Glue. Front. Immunol. 2018, 9, 1432. [Google Scholar] [CrossRef]
- Vragniau, C.; Bufton, J.C.; Garzoni, F.; Stermann, E.; Rabi, F.; Terrat, C.; Guidetti, M.; Josserand, V.; Williams, M.; Woods, C.J.; et al. Synthetic self-assembling ADDomer platform for highly efficient vaccination by genetically encoded multiepitope display. Sci. Adv. 2019, 5, w2853. [Google Scholar] [CrossRef]
- Chevillard, C.; Amen, A.; Besson, S.; Hannani, D.; Bally, I.; Dettling, V.; Gout, E.; Moreau, C.J.; Buisson, M.; Gallet, S.; et al. Elicitation of potent SARS-CoV-2 neutralizing antibody responses through immunization with a versatile adenovirus-inspired multimerization platform. Mol. Ther. 2022, 30, 1913–1925. [Google Scholar] [CrossRef]
- Jung, H.C.; Lee, S.H.; Lee, S.M.; An, Y.J.; Lee, J.H.; Lee, H.S.; Kang, S.G. Adaptive evolution of a hyperthermophilic archaeon pinpoints a formate transporter as a critical factor for the growth enhancement on formate. Sci. Rep. 2017, 7, 6124. [Google Scholar] [CrossRef]
- Nguyen, Q.T.; Yang, J.; Byun, J.W.; Pyo, H.M.; Park, M.Y.; Ku, B.K.; Nah, J.; Ryoo, S.; Wee, S.H.; Choi, K.S.; et al. Development of Monoclonal Antibody Specific to Foot-and-Mouth Disease Virus Type A for Serodiagnosis. Pathogens 2019, 8, 301. [Google Scholar] [CrossRef]
- Alexandersen, S.; Zhang, Z.; Donaldson, A.I. Aspects of the persistence of foot-and-mouth disease virus in animals-the carrier problem. Microbes Infect. 2002, 4, 1099–1110. [Google Scholar] [CrossRef]
- Parida, S. Vaccination against foot-and-mouth disease virus: Strategies and effectiveness. Expert Rev. Vaccines 2009, 8, 347–365. [Google Scholar] [CrossRef]
- Palatnik-de-Sousa, C.B.; Soares, I.S.; Rosa, D.S. Editorial: Epitope Discovery and Synthetic Vaccine Design. Front. Immunol. 2018, 9, 826. [Google Scholar] [CrossRef]
- Soria, I.; Quattrocchi, V.; Langellotti, C.; Perez-Filgueira, M.; Pega, J.; Gnazzo, V.; Romera, S.; Schammas, J.; Bucafusco, D.; Di Giacomo, S.; et al. Immune Response and Partial Protection against Heterologous Foot-and-Mouth Disease Virus Induced by Dendrimer Peptides in Cattle. J. Immunol. Res. 2018, 2018, 3497401. [Google Scholar] [CrossRef]
- Lee, H.S.; Park, S.Y.; Kim, A.Y.; Lee, S.O.; Kim, J.S.; Kim, H.; Youn, H.J.; Ko, Y.J. Recombinant vesicular stomatitis virus glycoprotein carrying a foot-and-mouth disease virus epitope as a vaccine candidate. J. Vet. Med. Sci. 2020, 82, 1155–1159. [Google Scholar] [CrossRef]
- Pan, Q.; Wang, H.; Ouyang, W.; Wang, X.; Bi, Z.; Xia, X.; Wang, Y.; He, K. Immunogenicity of adenovirus-derived porcine parvovirus-like particles displaying B and T cell epitopes of foot-and-mouth disease. Vaccine 2016, 34, 578–585. [Google Scholar] [CrossRef]
- Martinez-Solis, M.; Herrero, S.; Targovnik, A.M. Engineering of the baculovirus expression system for optimized protein production. Appl. Microbiol. Biotechnol. 2019, 103, 113–123. [Google Scholar] [CrossRef]
- Sharpe, H.R.; Gilbride, C.; Allen, E.; Belij-Rammerstorfer, S.; Bissett, C.; Ewer, K.; Lambe, T. The early landscape of coronavirus disease 2019 vaccine development in the UK and rest of the world. Immunology 2020, 160, 223–232. [Google Scholar] [CrossRef]
- Zhang, B.; Chao, C.W.; Tsybovsky, Y.; Abiona, O.M.; Hutchinson, G.B.; Moliva, J.I.; Olia, A.S.; Pegu, A.; Phung, E.; Stewart-Jones, G.; et al. A platform incorporating trimeric antigens into self-assembling nanoparticles reveals SARS-CoV-2-spike nanoparticles to elicit substantially higher neutralizing responses than spike alone. Sci. Rep. 2020, 10, 18149. [Google Scholar] [CrossRef]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, e1800376. [Google Scholar] [CrossRef]
- Lai, C.; Duan, S.; Ye, F.; Hou, X.; Li, X.; Zhao, J.; Yu, X.; Hu, Z.; Tang, Z.; Mo, F.; et al. The enhanced antitumor-specific immune response with mannose- and CpG-ODN-coated liposomes delivering TRP2 peptide. Theranostics 2018, 8, 1723–1739. [Google Scholar] [CrossRef]
- Terhuja, M.; Saravanan, P.; Tamilselvan, R.P. Comparative efficacy of virus like particle (VLP) vaccine of foot-and-mouth-disease virus (FMDV) type O adjuvanted with poly I:C or CpG in guinea pigs. Biologicals 2015, 43, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Beyer, T.; Herrmann, M.; Reiser, C.; Bertling, W.; Hess, J. Bacterial carriers and virus-like-particles as antigen delivery devices: Role of dendritic cells in antigen presentation. Curr. Drug Targets Infect. Disord. 2001, 1, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Nair-Gupta, P.; Blander, J.M. An updated view of the intracellular mechanisms regulating cross-presentation. Front. Immunol. 2013, 4, 401. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.G.; Jeong, J.; Hong, S.; Jung, W.H. Current Issues and Clinical Evidence in Tumor-Infiltrating Lymphocytes in Breast Cancer. J. Pathol. Transl. Med. 2015, 49, 355–363. [Google Scholar] [CrossRef]
- Suzuki, D.; Furukawa, K.; Kimura, F.; Shimizu, H.; Yoshidome, H.; Ohtsuka, M.; Kato, A.; Yoshitomi, H.; Miyazaki, M. Effects of perioperative immunonutrition on cell-mediated immunity, T helper type 1 (Th1)/Th2 differentiation, and Th17 response after pancreaticoduodenectomy. Surgery 2010, 148, 573–581. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Trypsin |
|---|---|
| Database | zjk_1_20201125 |
| Fixed modifications | Carbamidomethyl (C) |
| Variable modifications | Oxidation (M) |
| Missed cleavage | 2 |
| Peptide mass tolerance | 20 ppm |
| Fragment mass tolerance | 0.1 Da |
| Filter by score ≥ 20 |
| Groups | Number of Mice | Recombinant Proteins | Type and Composition | Immunization | Number of Injection | Immune Dose |
|---|---|---|---|---|---|---|
| 1 | 5 | ADDomer | W/O/W | SC | 2 | 200 µL |
| ISA 201 VG | ||||||
| 2 | 5 | ADDomer-RBT | W/O/W | SC | 2 | 200 µL |
| ISA 201 VG | ||||||
| 3 | 5 | ADDomer | ISA 201 VG + NAA | SC | 2 | 200 µL |
| 4 | 5 | ADDomer-RBT | ISA 201 VG + NAA | SC | 2 | 200 µL |
| 5 | 5 | Inactive vaccine | Commercial vaccine | SC | 2 | 200 µL |
| 6 | 5 | PBS | PBS | SC | 2 | 200 µL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, C.; Yan, Q.; Huang, J.; Liu, J.; Li, Y.; Wu, K.; Li, B.; Zhao, M.; Fan, S.; Ding, H.; et al. Using Self-Assembling ADDomer Platform to Display B and T Epitopes of Type O Foot-and-Mouth Disease Virus. Viruses 2022, 14, 1810. https://doi.org/10.3390/v14081810
Luo C, Yan Q, Huang J, Liu J, Li Y, Wu K, Li B, Zhao M, Fan S, Ding H, et al. Using Self-Assembling ADDomer Platform to Display B and T Epitopes of Type O Foot-and-Mouth Disease Virus. Viruses. 2022; 14(8):1810. https://doi.org/10.3390/v14081810
Chicago/Turabian StyleLuo, Chaowei, Quanhui Yan, Juncong Huang, Jiameng Liu, Yuwan Li, Keke Wu, Bingke Li, Mingqiu Zhao, Shuangqi Fan, Hongxing Ding, and et al. 2022. "Using Self-Assembling ADDomer Platform to Display B and T Epitopes of Type O Foot-and-Mouth Disease Virus" Viruses 14, no. 8: 1810. https://doi.org/10.3390/v14081810