

Third Early “Booster” Dose Strategy in France of bnt162b2 SARS-CoV-2 Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients Enhances Neutralizing Antibody Responses
 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Measurement of Anti-S-RBD IgG Antibody Levels
2.3. Cell Lines and Viruses
2.4. Fluorescence-Based Neutralization Assay
2.5. Statistical Analyses
3. Results
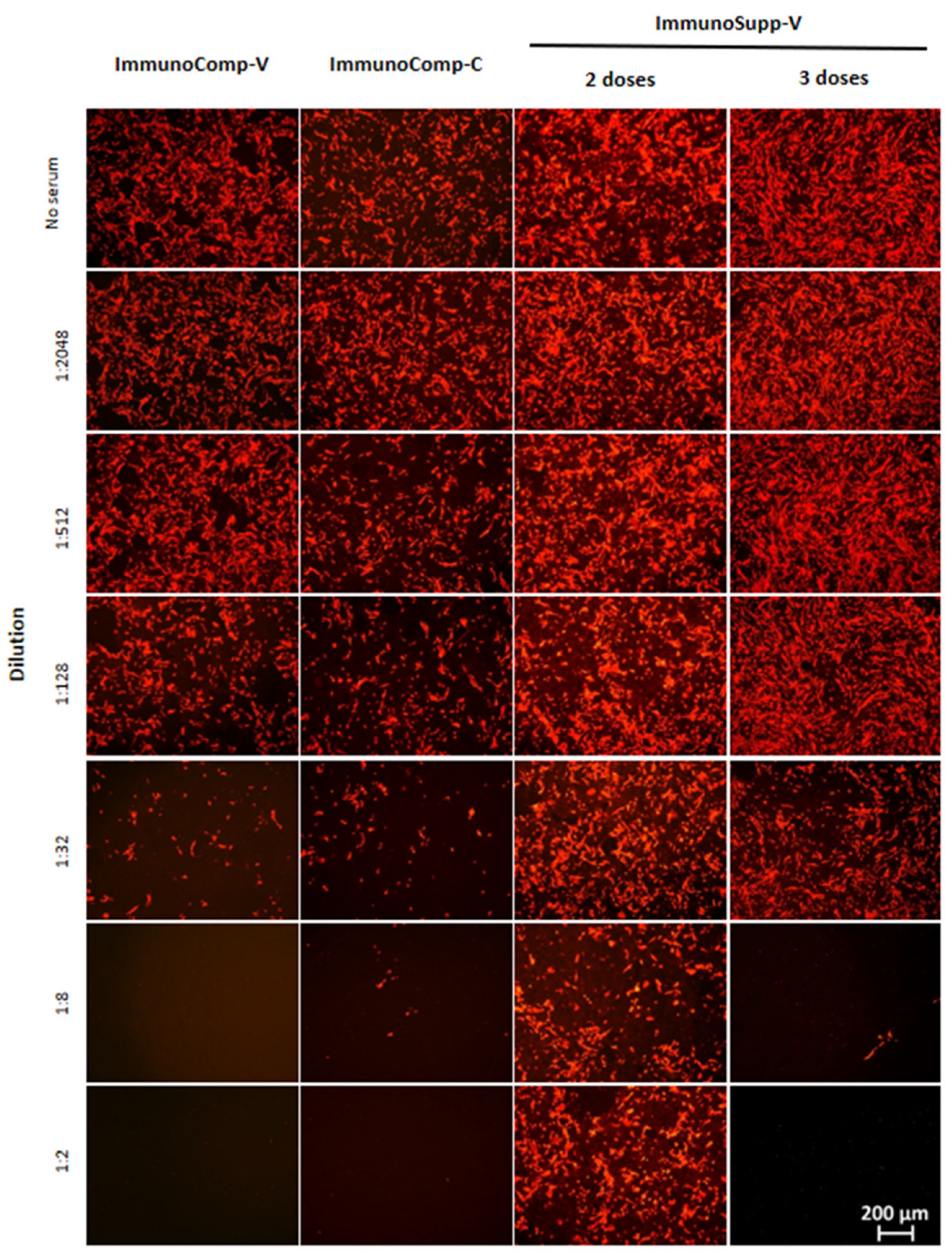
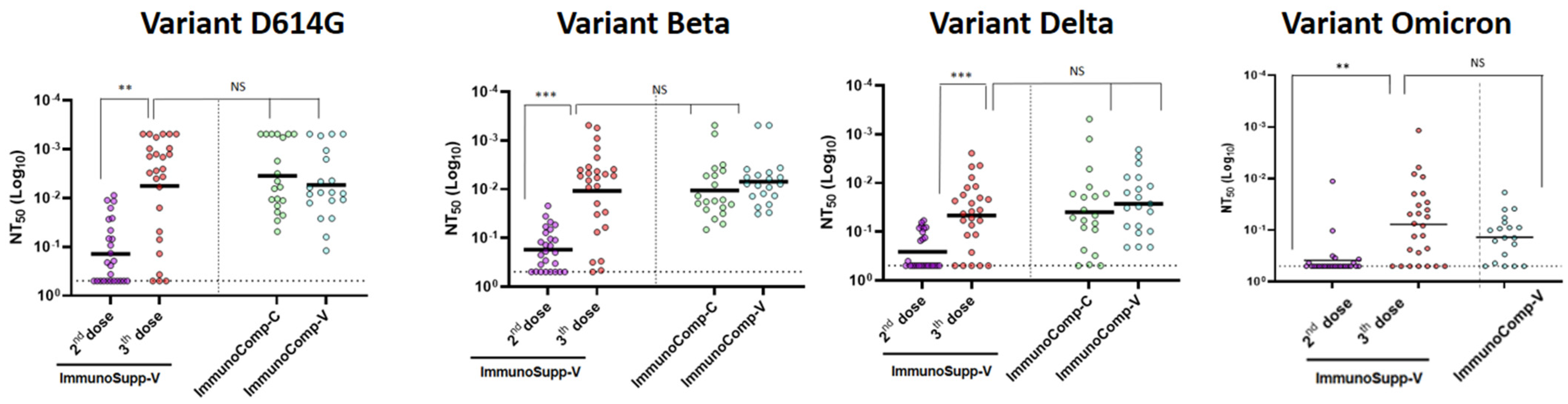
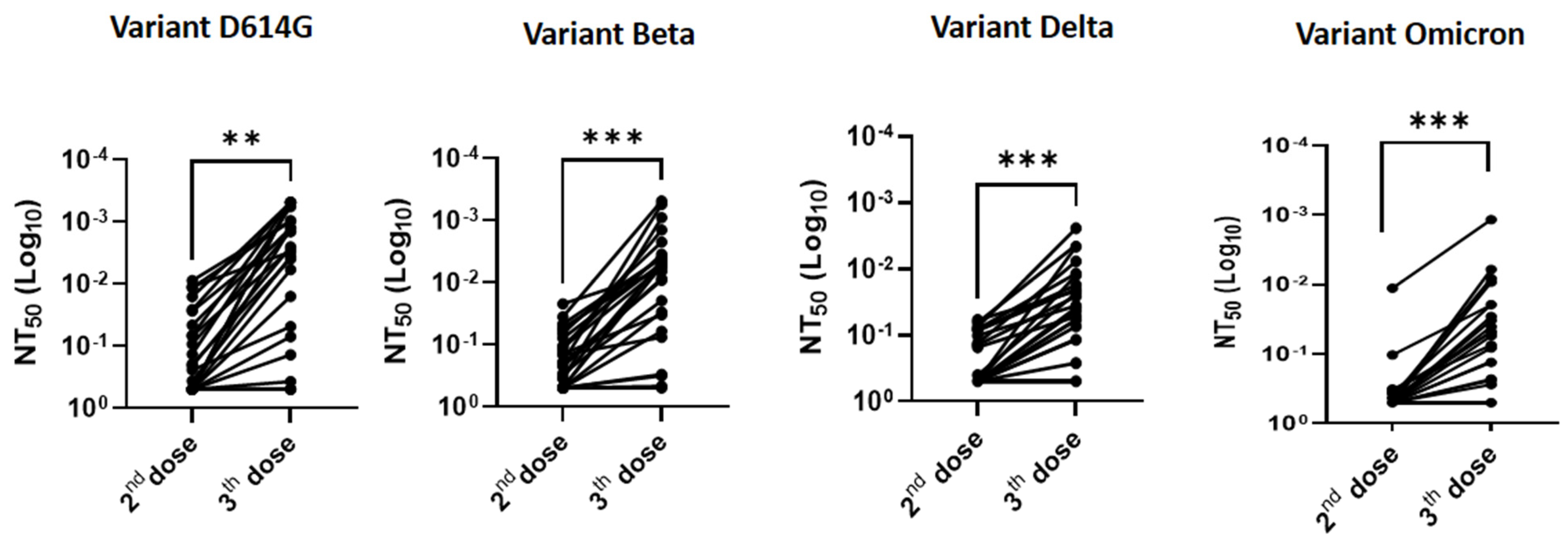
3.1. Effectiveness of a Third “Booster” Vaccine Dose in HSCT Recipients vs. Two Doses Only in Immunocompetent Patients
3.2. Relationship between Anti-S-RBD Antibody Responses and Neutralization Titers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Shrotri, M.; Navaratnam, A.M.; Nguyen, V.; Byrne, T.; Geismar, C.; Fragaszy, E.; Beale, S.; Fong, W.L.E.; Patel, P.; Kovar, J.; et al. Spike-antibody waning after second dose of BNT162b2 or ChAdOx1. Lancet 2021, 398, 385–387. [Google Scholar] [CrossRef]
- Israel, A.; Merzon, E.; Schäffer, A.A.; Shenhar, Y.; Green, I.; Golan-Cohen, A.; Ruppin, E.; Magen, E.; Vinker, S. Elapsed time since BNT162b2 vaccine and risk of SARS-CoV-2 infection: Test negative design study. BMJ 2021, 375, e067873. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.; Haas, E.J.; Milo, R.; Alroy-Preis, S.; Ash, N.; Huppert, A. Waning Immunity after the BNT162b2 Vaccine in Israel. N. Engl. J. Med. 2021, 385, e85. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.G.; Lustig, Y.; Cohen, C.; Fluss, R.; Indenbaum, V.; Amit, S.; Doolman, R.; Asraf, K.; Mendelson, E.; Ziv, A.; et al. Waning Immune Humoral Response to BNT162b2 COVID-19 Vaccine over 6 Months. N. Engl. J. Med. 2021, 385, e84. [Google Scholar] [CrossRef] [PubMed]
- Munro, A.P.; Janani, L.; Cornelius, V.; Aley, P.K.; Babbage, G.; Baxter, D.; Bula, M.; Cathie, K.; Chatterjee, K.; Dodd, K.; et al. Safety and immunogenicity of seven COVID-19 vaccines as a third dose (booster) following two doses of ChAdOx1 nCov-19 or BNT162b2 in the UK (COV-BOOST): A blinded, multicentre, randomised, controlled, phase 2 trial. Lancet 2021, 398, 2258–2276. [Google Scholar] [CrossRef]
- Falsey, A.R.; Frenck, R.W., Jr.; Walsh, E.E.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Bailey, R.; Swanson, K.A.; Xu, X.; et al. SARS-CoV-2 Neutralization with BNT162b2 Vaccine Dose 3. N. Engl. J. Med. 2021, 385, 1627–1629. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Goldberg, Y.; Mandel, M.; Bodenheimer, O.; Freedman, L.; Kalkstein, N.; Mizrahi, B.; Alroy-Preis, S.; Ash, N.; Milo, R.; et al. Protection of BNT162b2 Vaccine Booster against COVID-19 in Israel. N. Engl. J. Med. 2021, 385, 1393–1400. [Google Scholar] [CrossRef]
- Barda, N.; Dagan, N.; Cohen, C.; Hernán, M.A.; Lipsitch, M.; Kohane, I.S.; Reis, B.Y.; Balicer, R.D. Effectiveness of a third dose of the BNT162b2 mRNA COVID-19 vaccine for preventing severe outcomes in Israel: An observational study. Lancet 2021, 398, 2093–2100. [Google Scholar] [CrossRef]
- Redjoul, R.; Le Bouter, A.; Parinet, V.; Fourati, S.; Maury, S. Antibody response after third BNT162b2 dose in recipients of allogeneic HSCT. Lancet Haematol. 2021, 8, e681–e683. [Google Scholar] [CrossRef]
- Kamar, N.; Abravanel, F.; Marion, O.; Couat, C.; Izopet, J.; Del Bello, A. Three Doses of an mRNA COVID-19 Vaccine in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2021, 385, 661–662. [Google Scholar] [CrossRef]
- Del Bello, A.; Abravanel, F.; Marion, O.; Couat, C.; Esposito, L.; Lavayssière, L.; Izopet, J.; Kamar, N. Efficiency of a boost with a third dose of anti-SARS-CoV-2 messenger RNA-based vaccines in solid organ transplant recipients. Am. J. Transplant. 2021, 22, 322. [Google Scholar] [CrossRef]
- Planas, D.; Bruel, T.; Grzelak, L.; Guivel-Benhassine, F.; Staropoli, I.; Porrot, F.; Planchais, C.; Buchrieser, J.; Rajah, M.M.; Bishop, E.; et al. Sensitivity of infectious SARS-CoV-2 B.1.1.7 and B.1.351 variants to neutralizing antibodies. Nat. Med. 2021, 27, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, Y.; Xia, H.; Zou, J.; Weaver, S.C.; Swanson, K.A.; Cai, H.; Cutler, M.; Cooper, D.; Muik, A.; et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature 2021, 596, 273–275. [Google Scholar] [CrossRef] [PubMed]
- Mlcochova, P.; Kemp, S.A.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S.; et al. SARS-CoV-2 B.1.617.2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J.; et al. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Mamez, A.C.; Pradier, A.; Giannotti, F.; Petitpas, A.; Urdiola, M.F.; Vu, D.L.; Masouridi-Levrat, S.; Morin, S.; Dantin, S.; Clerc-Renaud, D.; et al. Antibody responses to SARS-CoV2 vaccination in allogeneic hematopoietic stem cell transplant recipients. Bone Marrow Transplant. 2021, 56, 3094–3096. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Saunders, N.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.-H.; Porrot, F.; Staropoli, I.; Lemoine, F.; et al. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| “ImmunoSupp-V” Allogeneic HSCT Recipients (n = 26) | “ImmunoComp-V” Immunocompetent Vaccinated Individuals (n = 22) | “ImmunoComp-C” Immunocompetent Convalescent Patients (n = 20) | |
|---|---|---|---|
| Median age (min-max), years | 61 (31–75) | 40 (21–56) | 71 (26–94) |
| % male gender (n/N) | 73.1% (19/26) | 40.9% (9/22) | 40.0% (8/20) |
| Wards of origin | |||
| Hematology unit [% (n/N)] | 100.0% (26/26) | - | - |
| Geriatric wards [% (n/N)] | - | - | 45.0% (9/20) |
| Medical wards [% (n/N)] | - | - | 10.0% (2/20) |
| Outpatients [% (n/N)] | - | - | 20.0% (4/20) |
| Healthcare workers [% (n/N)] | - | 100.0% (22/22) | 25.0% (5/20) |
| Median time between HSCT and initiation of vaccination, (min-max), months | 14 (3–100) | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed-Belkacem, A.; Redjoul, R.; Brillet, R.; Ahnou, N.; Leclerc, M.; López-Molina, D.S.; Soulier, A.; Gourgeon, A.; Rodriguez, C.; Maury, S.; et al. Third Early “Booster” Dose Strategy in France of bnt162b2 SARS-CoV-2 Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients Enhances Neutralizing Antibody Responses. Viruses 2022, 14, 1928. https://doi.org/10.3390/v14091928
Ahmed-Belkacem A, Redjoul R, Brillet R, Ahnou N, Leclerc M, López-Molina DS, Soulier A, Gourgeon A, Rodriguez C, Maury S, et al. Third Early “Booster” Dose Strategy in France of bnt162b2 SARS-CoV-2 Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients Enhances Neutralizing Antibody Responses. Viruses. 2022; 14(9):1928. https://doi.org/10.3390/v14091928
Chicago/Turabian StyleAhmed-Belkacem, Abdelhakim, Rabah Redjoul, Rozenn Brillet, Nazim Ahnou, Mathieu Leclerc, Dennis Salomón López-Molina, Alexandre Soulier, Aurélie Gourgeon, Christophe Rodriguez, Sébastien Maury, and et al. 2022. "Third Early “Booster” Dose Strategy in France of bnt162b2 SARS-CoV-2 Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients Enhances Neutralizing Antibody Responses" Viruses 14, no. 9: 1928. https://doi.org/10.3390/v14091928
APA StyleAhmed-Belkacem, A., Redjoul, R., Brillet, R., Ahnou, N., Leclerc, M., López-Molina, D. S., Soulier, A., Gourgeon, A., Rodriguez, C., Maury, S., Pawlotsky, J.-M., & Fourati, S. (2022). Third Early “Booster” Dose Strategy in France of bnt162b2 SARS-CoV-2 Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients Enhances Neutralizing Antibody Responses. Viruses, 14(9), 1928. https://doi.org/10.3390/v14091928