

A Potential Role for Substance P in West Nile Virus Neuropathogenesis

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Husbandry Practices, Monitoring for All Animals, and Specimen Collection
2.2. Viruses and Drug Compounds
2.3. Experimental Design and Groups
2.4. Detection of SP and WNV
2.5. MRI
2.6. Analysis
3. Results
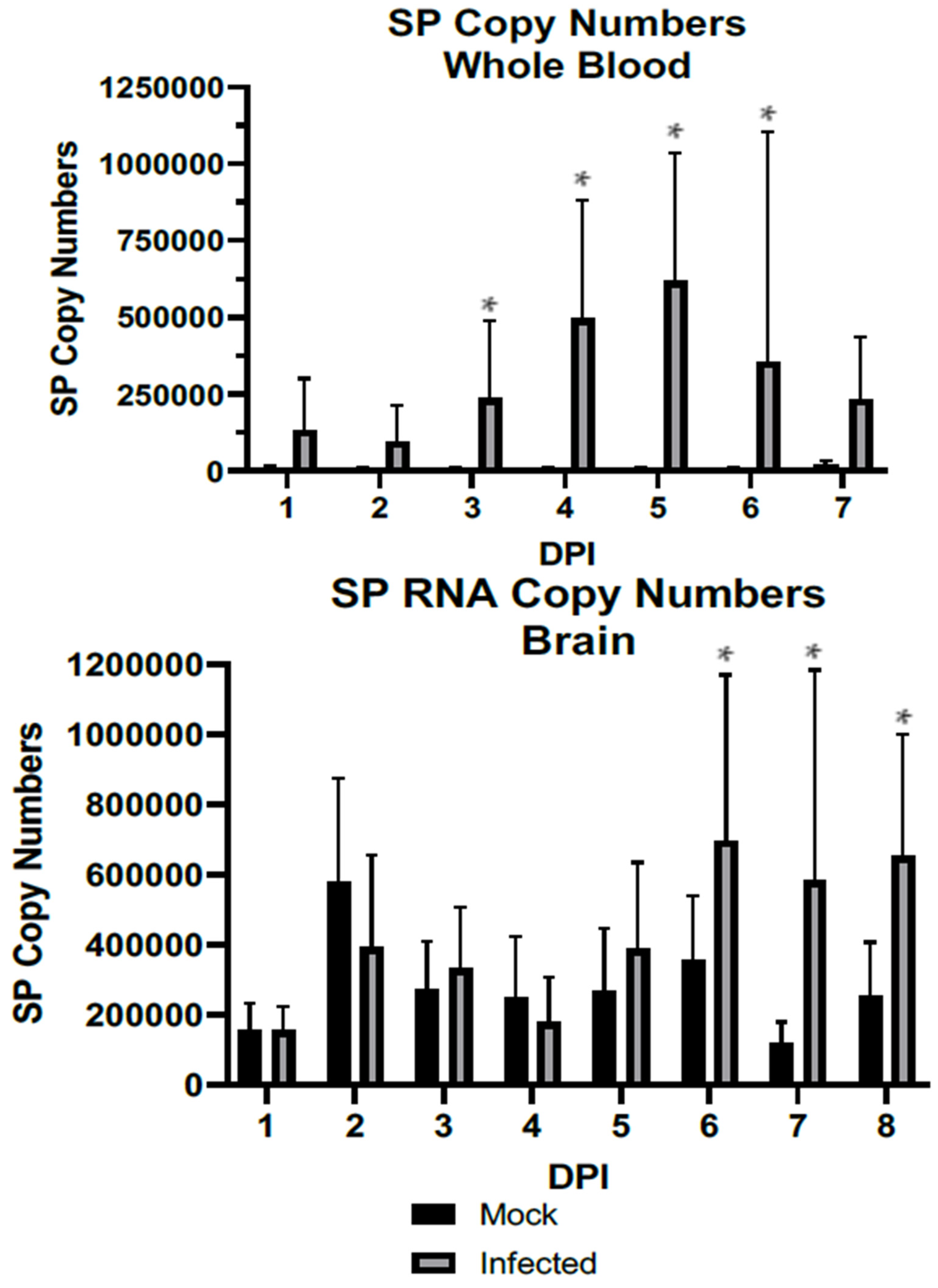
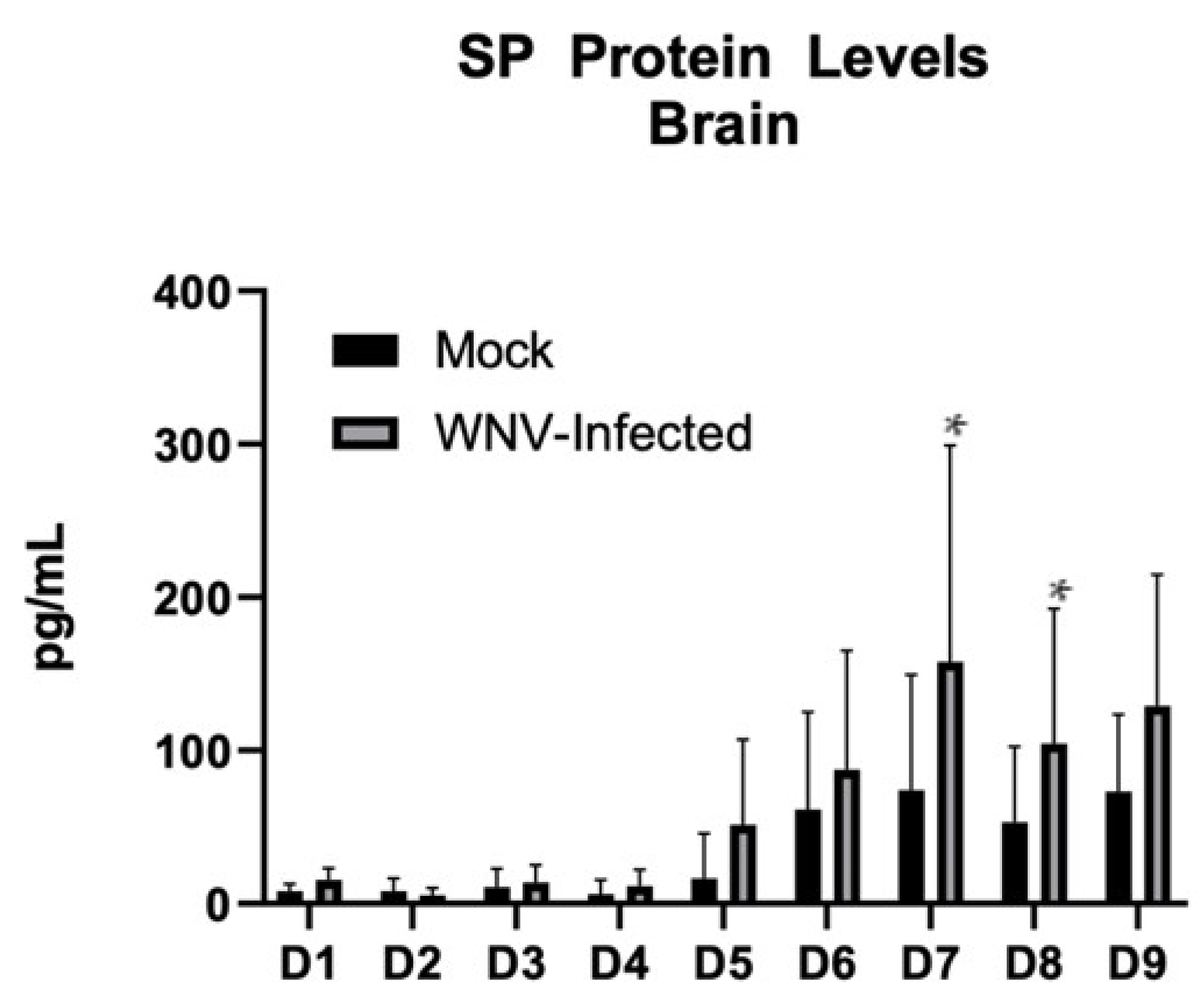
3.1. SP Becomes Significantly Elevated over the Course of WNV Infection
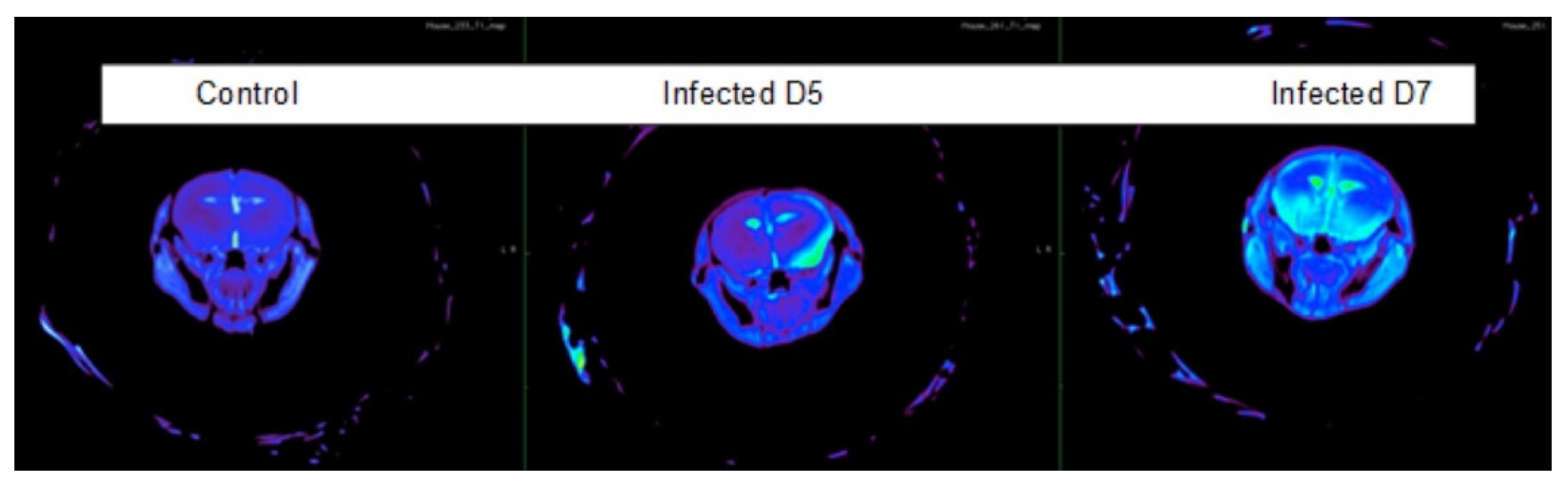
3.2. Blood–Brain Barrier Permeability Is Visualized during Infection
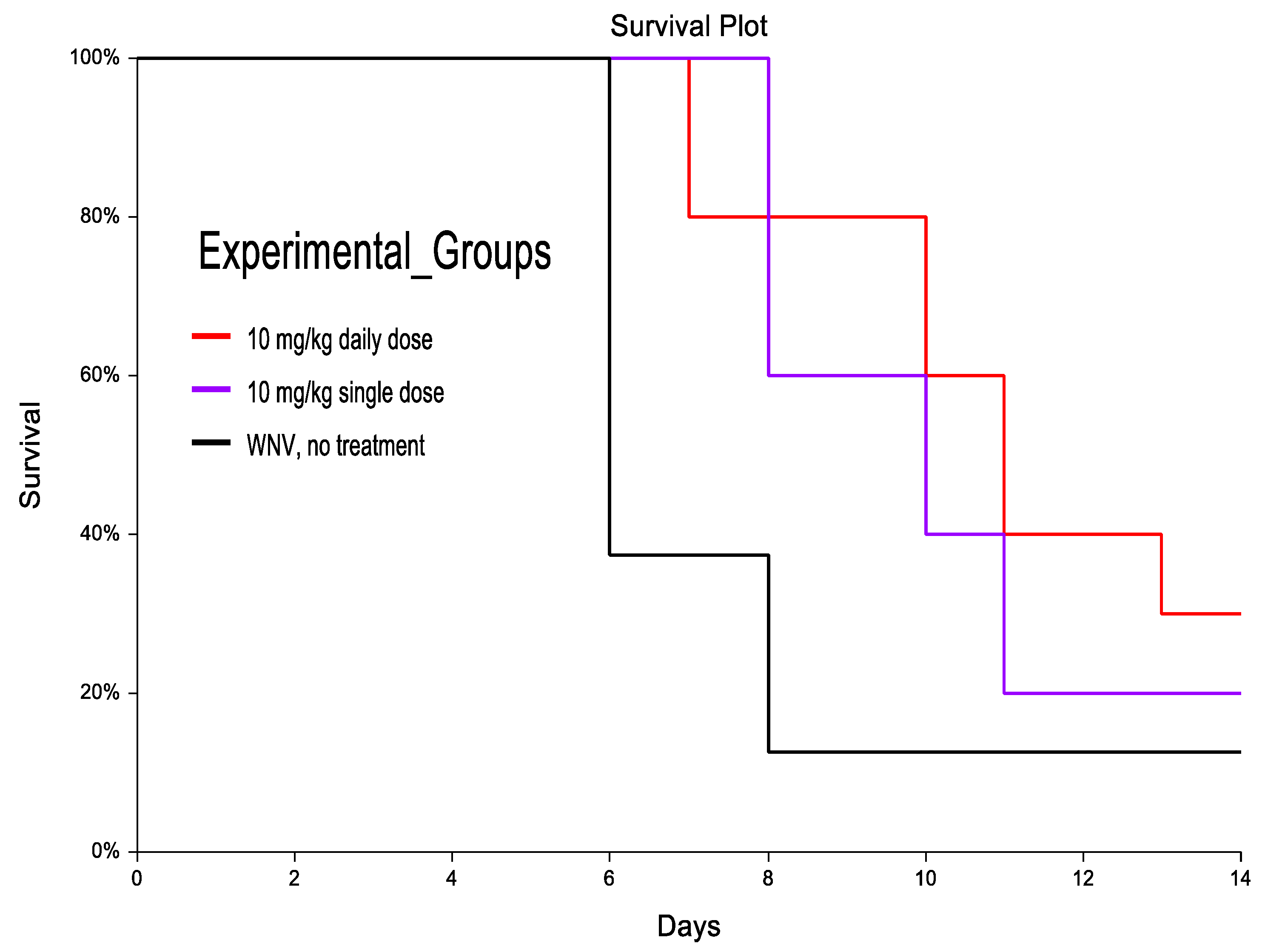
3.3. CH123001 Increased Survival in Treatment Groups and Prolonged Time to Death
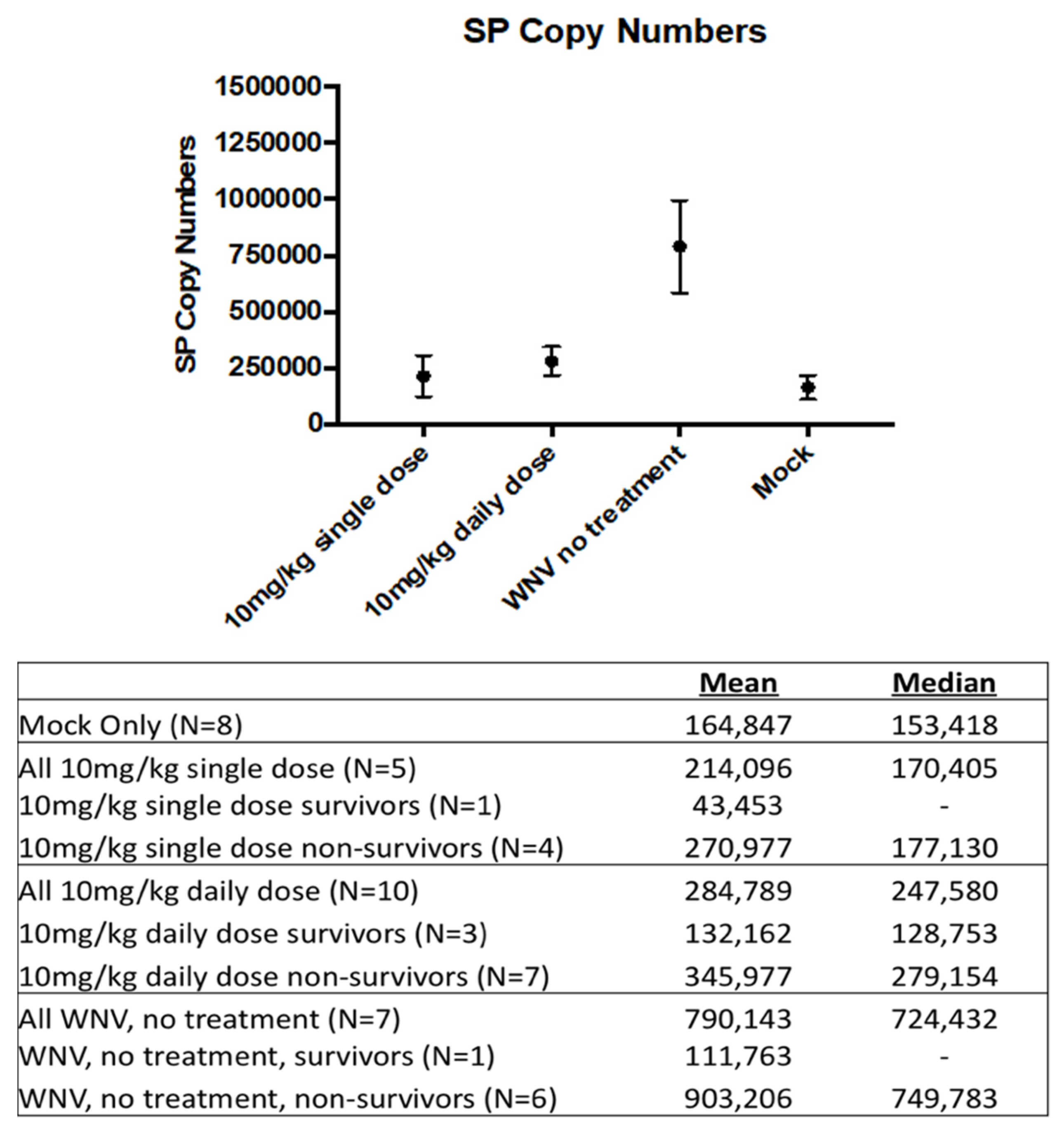
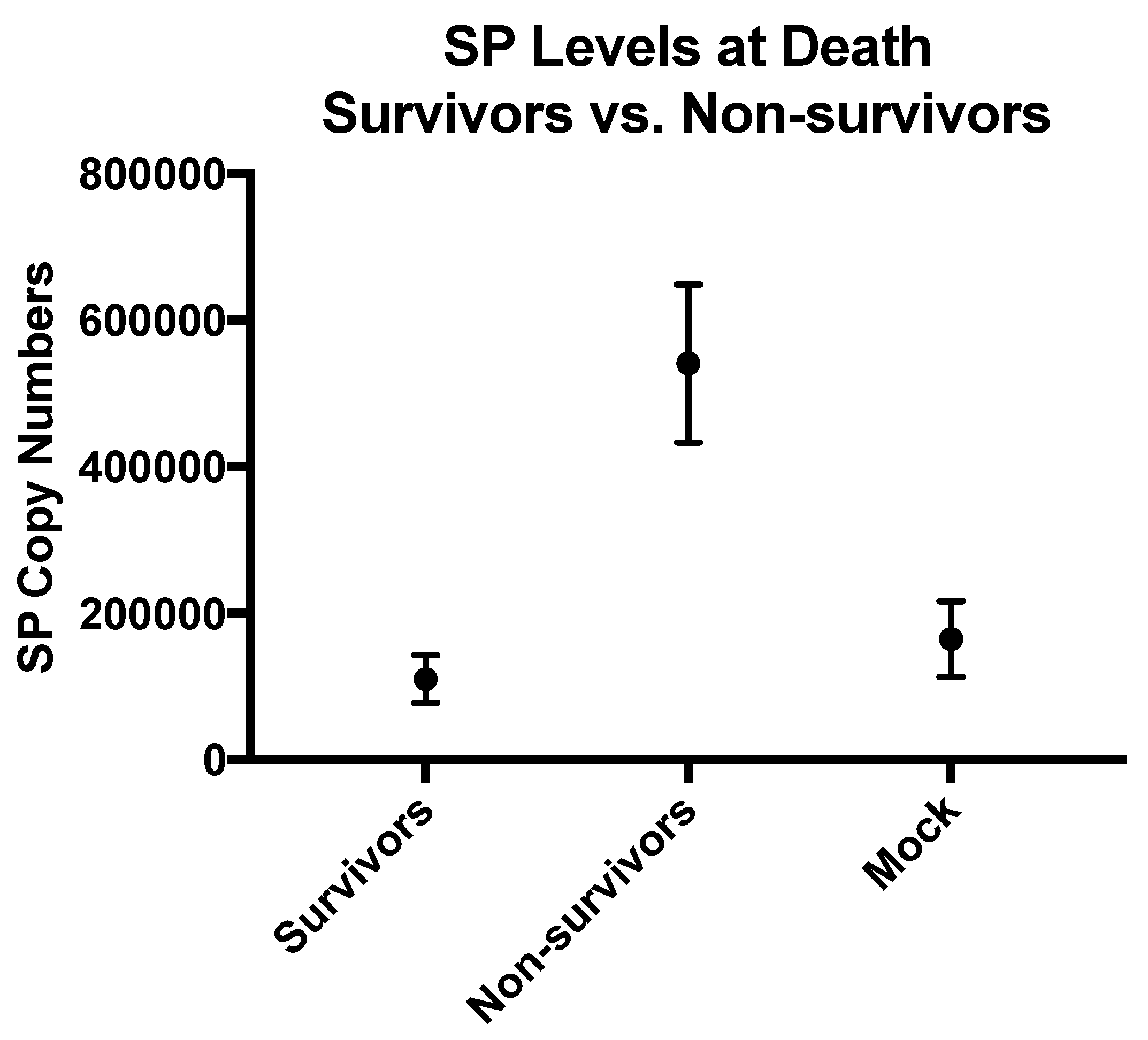
3.4. SP Levels Were Decreased in the Treated Groups and Correlated with Survival
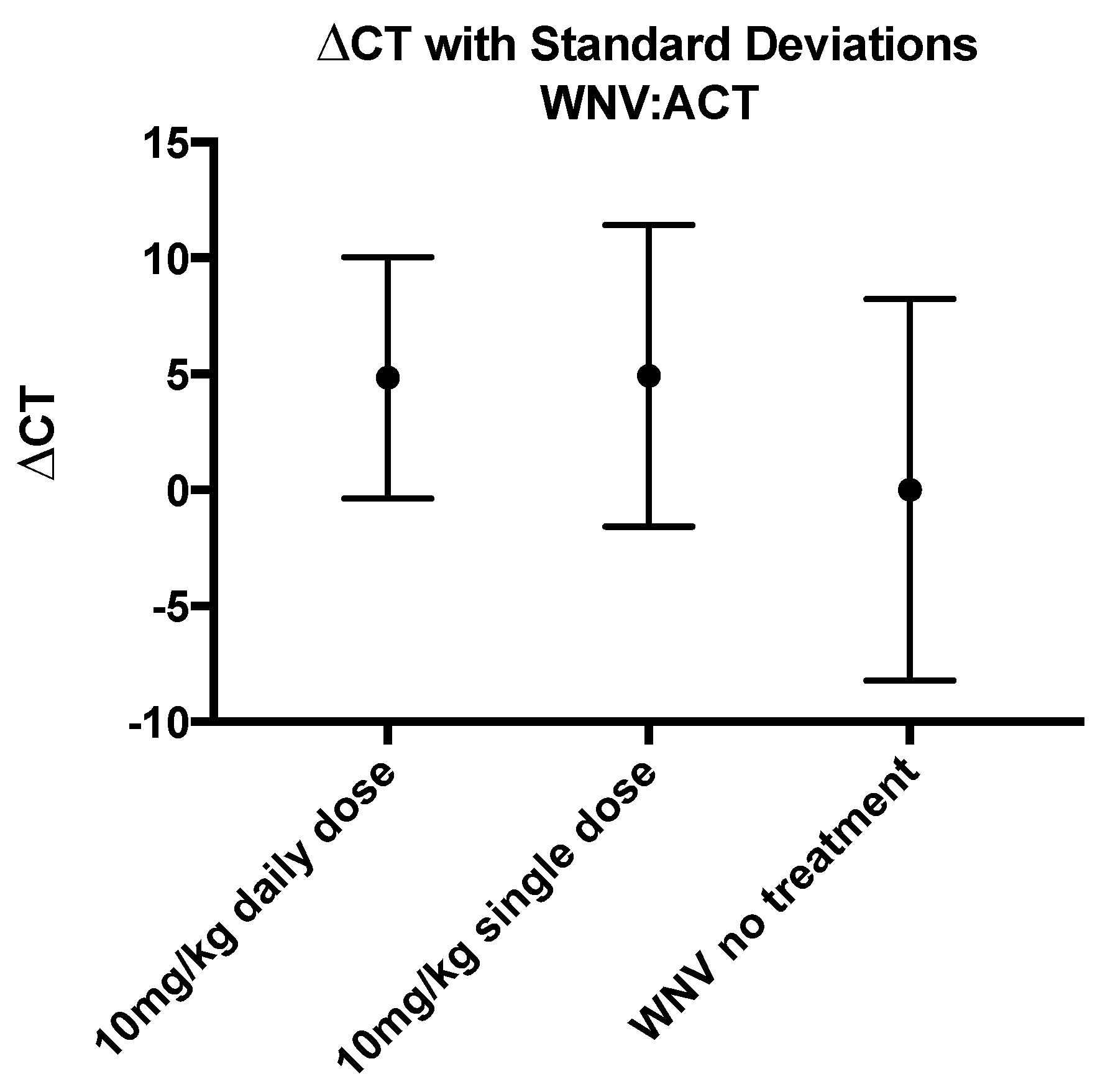
3.5. Levels of WNV RNA Were Not Affected by CH123001 Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ronca, S.E.; Murray, K.O.; Nolan, M.S. Cumulative Incidence of West Nile Virus Infection, Continental United States, 1999-2016. Emerg. Infect. Dis. 2019, 25, 325–327. [Google Scholar] [CrossRef]
- Mostashari, F.; Bunning, M.L.; Kitsutani, P.T.; Singer, D.A.; Nash, D.; Cooper, M.J.; Katz, N.; Liljebjelke, K.A.; Biggerstaff, B.J.; Fine, A.D.; et al. Epidemic West Nile encephalitis, New York, 1999: Results of a household-based seroepidemiological survey. Lancet 2001, 358, 261–264. [Google Scholar] [CrossRef]
- Carson, P.J.; Borchardt, S.M.; Custer, B.; Prince, H.E.; Dunn-Williams, J.; Winkelman, V.; Tobler, L.; Biggerstaff, B.J.; Lanciotti, R.; Petersen, L.R.; et al. Neuroinvasive disease and West Nile virus infection, North Dakota, USA, 1999–2008. Emerg. Infect. Dis. 2012, 18, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Weatherhead, J.E.; Miller, V.E.; Garcia, M.N.; Hasbun, R.; Salazar, L.; Dimachkie, M.M.; Murray, K.O. Long-term neurological outcomes in West Nile virus-infected patients: An observational study. Am. J. Trop Med. Hyg. 2015, 92, 1006–1012. [Google Scholar] [CrossRef] [PubMed]
- Bode, A.V.; Sejvar, J.J.; Pape, W.J.; Campbell, G.L.; Marfin, A.A. West Nile virus disease: A descriptive study of 228 patients hospitalized in a 4-county region of Colorado in 2003. Clin. Infect. Dis. 2006, 42, 1234–1240. [Google Scholar] [CrossRef]
- Sejvar, J.J. Clinical manifestations and outcomes of West Nile virus infection. Viruses 2014, 6, 606–623. [Google Scholar] [CrossRef]
- Erickson, T.A.; Ronca, S.E.; Gunter, S.M.; Brown, E.L.; Hasbun, R.; Murray, K.O. Zoonotic Disease Testing Practices in Pediatric Patients with Meningitis and Encephalitis in a Subtropical Region. Pathogens 2022, 11, 501. [Google Scholar] [CrossRef]
- Vanichanan, J.; Salazar, L.; Wootton, S.H.; Aguilera, E.; Garcia, M.N.; Murray, K.O.; Hasbun, R. Use of Testing for West Nile Virus and Other Arboviruses. Emerg. Infect. Dis. 2016, 22, 1587. [Google Scholar] [CrossRef]
- Sampson, B.A.; Ambrosi, C.; Charlot, A.; Reiber, K.; Veress, J.F.; Armbrustmacher, V. The pathology of human West Nile Virus infection. Hum. Pathol. 2000, 31, 527–531. [Google Scholar] [CrossRef]
- Sampson, B.A.; Armbrustmacher, V. West Nile encephalitis: The neuropathology of four fatalities. Ann. N. Y. Acad. Sci. 2001, 951, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.D.; Leeman, S.E. Neurokinin-1 receptor: Functional significance in the immune system in reference to selected infections and inflammation. Ann. N. Y. Acad. Sci. 2011, 1217, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, P.; Cioni, C.; Santonini, R.; Paccagnini, E. Substance P antagonist blocks leakage and reduces activation of cytokine-stimulated rat brain endothelium. J. Neuroimmunol. 2002, 131, 41–49. [Google Scholar] [CrossRef]
- Corrigan, F.; Vink, R.; Turner, R.J. Inflammation in acute CNS injury: A focus on the role of substance P. Br. J. Pharmacol. 2016, 173, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.M.; Turner, R.J.; Vink, R. Blocking neurogenic inflammation for the treatment of acute disorders of the central nervous system. Int. J. Inflamm. 2013, 2013, 578480. [Google Scholar] [CrossRef] [PubMed]
- Ziebell, J.M.; Morganti-Kossmann, M.C. Involvement of pro- and anti-inflammatory cytokines and chemokines in the pathophysiology of traumatic brain injury. Neurotherapeutics 2010, 7, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Setiawan, T.; Hang, L.; Stoyanoff, K.; Weinstock, J.V. Interleukin-12 (IL-12) and IL-23 induction of substance p synthesis in murine T cells and macrophages is subject to IL-10 and transforming growth factor beta regulation. Infect. Immun. 2008, 76, 3651–3656. [Google Scholar] [CrossRef]
- Donkin, J.J.; Nimmo, A.J.; Cernak, I.; Blumbergs, P.C.; Vink, R. Substance P is associated with the development of brain edema and functional deficits after traumatic brain injury. J. Cereb. Blood Flow Metab. 2009, 29, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Ebner, K.; Singewald, N. The role of substance P in stress and anxiety responses. Amino Acids 2006, 31, 251–272. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.B.; Young, A.D.; Marriott, I. The Therapeutic Potential of Targeting Substance P/NK-1R Interactions in Inflammatory CNS Disorders. Front. Cell. Neurosci. 2016, 10, 296. [Google Scholar] [CrossRef]
- Li, C.; Micci, M.A.; Murthy, K.S.; Pasricha, P.J. Substance P is essential for maintaining gut muscle contractility: A novel role for coneurotransmission revealed by botulinum toxin. Am. J. Physiol. Gastrointest Liver Physiol. 2014, 306, G839–G848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide substance P and the immune response. Cell. Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Ramnath, R.D.; Zhi, L.; Tamizhselvi, R.; Bhatia, M. Substance P enhances NF-kappaB transactivation and chemokine response in murine macrophages via ERK1/2 and p38 MAPK signaling pathways. Am. J. Physiol. Cell. Physiol. 2008, 294, C1586–C1596. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Qu, C.; Lu, X.; Zhang, S. Activation of microglia by histamine and substance P. Cell. Physiol. Biochem. 2014, 34, 768–780. [Google Scholar] [CrossRef]
- Martinez, A.N.; Burmeister, A.R.; Ramesh, G.; Doyle-Meyers, L.; Marriott, I.; Philipp, M.T. Aprepitant limits in vivo neuroinflammatory responses in a rhesus model of Lyme neuroborreliosis. J. Neuroinflamm. 2017, 14, 37. [Google Scholar] [CrossRef]
- Martinez, A.N.; Ramesh, G.; Jacobs, M.B.; Philipp, M.T. Antagonist of the neurokinin-1 receptor curbs neuroinflammation in ex vivo and in vitro models of Lyme neuroborreliosis. J. Neuroinflamm. 2015, 12, 243. [Google Scholar] [CrossRef]
- Vinet-Oliphant, H.; Alvarez, X.; Buza, E.; Borda, J.T.; Mohan, M.; Aye, P.P.; Tuluc, F.; Douglas, S.D.; Lackner, A.A. Neurokinin-1 receptor (NK1-R) expression in the brains of SIV-infected rhesus macaques: Implications for substance P in NK1-R immune cell trafficking into the CNS. Am. J. Pathol. 2010, 177, 1286–1297. [Google Scholar] [CrossRef]
- Hoffmann, T.; Bos, M.; Stadler, H.; Schnider, P.; Hunkeler, W.; Godel, T.; Galley, G.; Ballard, T.M.; Higgins, G.A.; Poli, S.M.; et al. Design and synthesis of a novel, achiral class of highly potent and selective, orally active neurokinin-1 receptor antagonists. Bioorg Med. Chem. Lett. 2006, 16, 1362–1365. [Google Scholar] [CrossRef] [PubMed]
- Piorkowski, G.; Baronti, C.; de Lamballerie, X.; de Fabritus, L.; Bichaud, L.; Pastorino, B.A.; Bessaud, M. Development of generic Taqman PCR and RT-PCR assays for the detection of DNA and mRNA of beta-actin-encoding sequences in a wide range of animal species. J. Virol. Methods 2014, 202, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Narayanaswami, P.; Edwards, L.L.; Hyde, C.; Page, C.; Hastings, N.E. West Nile meningitis/encephalitis: Experience with corticosteroid therapy. Neurology 2004, 62, A404. [Google Scholar]
- Walid, M.S.; Mahmoud, F.A. Successful treatment with intravenous immunoglobulin of acute flaccid paralysis caused by west nile virus. Perm. J. 2009, 13, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Amanna, I.J.; Slifka, M.K. Current trends in West Nile virus vaccine development. Expert Rev. Vaccines 2014, 13, 589–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.; Safriel, Y.; Sohi, J.; Llave, A.; Weathers, S. West Nile virus infection: MR imaging findings in the nervous system. AJNR Am. J. Neuroradiol. 2005, 26, 289–297. [Google Scholar]
- Patel, C.B.; Trikamji, B.; Mathisen, G.; Yim, C.; Zipser, B.; Mishra, S. MRI Ventral Nerve Root Enhancement in Five Patients Presenting With Extremity Weakness Secondary to Neuroinvasive West Nile Virus. J. Clin. Neuromuscul. Dis. 2016, 18, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Puchalski, A.; Liu, A. A Case of West Nile Encephalitis That Develops into a Disease of Deep White Matter on MRI over a Two-Week Span. Case Rep. Neurol. Med. 2016, 2016, 4389691. [Google Scholar] [CrossRef] [PubMed]
- Murray, K.O.; Nolan, M.S.; Ronca, S.E.; Datta, S.; Govindarajan, K.; Narayana, P.A.; Salazar, L.; Woods, S.P.; Hasbun, R. The Neurocognitive and MRI Outcomes of West Nile Virus Infection: Preliminary Analysis Using an External Control Group. Front. Neurol. 2018, 9, 111. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, J.M.; Frick, A.; Ahs, F.; Linnman, C.; Appel, L.; Jonasson, M.; Lubberink, M.; Langstrom, B.; Frans, O.; von Knorring, L.; et al. Association between amygdala neurokinin-1 receptor availability and anxiety-related personality traits. Transl. Psychiatry 2018, 8, 168. [Google Scholar] [CrossRef]
- Ebner, K.; Rupniak, N.M.; Saria, A.; Singewald, N. Substance P in the medial amygdala: Emotional stress-sensitive release and modulation of anxiety-related behavior in rats. Proc. Natl. Acad. Sci. USA 2004, 101, 4280–4285. [Google Scholar] [CrossRef]
- Herpfer, I.; Lieb, K. Substance P receptor antagonists in psychiatry: Rationale for development and therapeutic potential. CNS Drugs 2005, 19, 275–293. [Google Scholar] [CrossRef] [PubMed]
- Kahnau, P.; Habedank, A.; Diederich, K.; Lewejohann, L. Behavioral Methods for Severity Assessment. Animals 2020, 10, 1136. [Google Scholar] [CrossRef]
- Hohlbaum, K.; Bert, B.; Dietze, S.; Palme, R.; Fink, H.; Thone-Reineke, C. Severity classification of repeated isoflurane anesthesia in C57BL/6JRj mice-Assessing the degree of distress. PLoS ONE 2017, 12, e0179588. [Google Scholar] [CrossRef]
- Chirico, M.T.T.; Guedes, M.R.; Vieira, L.G.; Reis, T.O.; Dos Santos, A.M.; Souza, A.B.F.; Ribeiro, I.M.L.; Noronha, S.; Nogueira, K.O.; Oliveira, L.A.M.; et al. Lasting effects of ketamine and isoflurane administration on anxiety- and panic-like behavioral responses in Wistar rats. Life Sci. 2021, 276, 119423. [Google Scholar] [CrossRef] [PubMed]
- Yonezaki, K.; Uchimoto, K.; Miyazaki, T.; Asakura, A.; Kobayashi, A.; Takase, K.; Goto, T. Postanesthetic effects of isoflurane on behavioral phenotypes of adult male C57BL/6J mice. PLoS ONE 2015, 10, e0122118. [Google Scholar] [CrossRef] [Green Version]
- Suvas, S. Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J. Immunol. 2017, 199, 1543–1552. [Google Scholar] [CrossRef]
- Vilisaar, J.; Kawabe, K.; Braitch, M.; Aram, J.; Furtun, Y.; Fahey, A.J.; Chopra, M.; Tanasescu, R.; Tighe, P.J.; Gran, B.; et al. Reciprocal Regulation of Substance P and IL-12/IL-23 and the Associated Cytokines, IFNgamma/IL-17: A Perspective on the Relevance of This Interaction to Multiple Sclerosis. J. Neuroimmune Pharmacol. 2015, 10, 457–467. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronca, S.E.; Gunter, S.M.; Kairis, R.B.; Lino, A.; Romero, J.; Pautler, R.G.; Nimmo, A.; Murray, K.O. A Potential Role for Substance P in West Nile Virus Neuropathogenesis. Viruses 2022, 14, 1961. https://doi.org/10.3390/v14091961
Ronca SE, Gunter SM, Kairis RB, Lino A, Romero J, Pautler RG, Nimmo A, Murray KO. A Potential Role for Substance P in West Nile Virus Neuropathogenesis. Viruses. 2022; 14(9):1961. https://doi.org/10.3390/v14091961
Chicago/Turabian StyleRonca, Shannon E., Sarah M. Gunter, Rebecca Berry Kairis, Allison Lino, Jonathan Romero, Robia G. Pautler, Alan Nimmo, and Kristy O. Murray. 2022. "A Potential Role for Substance P in West Nile Virus Neuropathogenesis" Viruses 14, no. 9: 1961. https://doi.org/10.3390/v14091961
APA StyleRonca, S. E., Gunter, S. M., Kairis, R. B., Lino, A., Romero, J., Pautler, R. G., Nimmo, A., & Murray, K. O. (2022). A Potential Role for Substance P in West Nile Virus Neuropathogenesis. Viruses, 14(9), 1961. https://doi.org/10.3390/v14091961