

Genomic and Epidemiological Features of COVID-19 in the Novosibirsk Region during the Beginning of the Pandemic

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collecting Samples
2.2. Detection of SARS-CoV-2 RNA
2.3. SARS-CoV-2 Genomes
3. Results
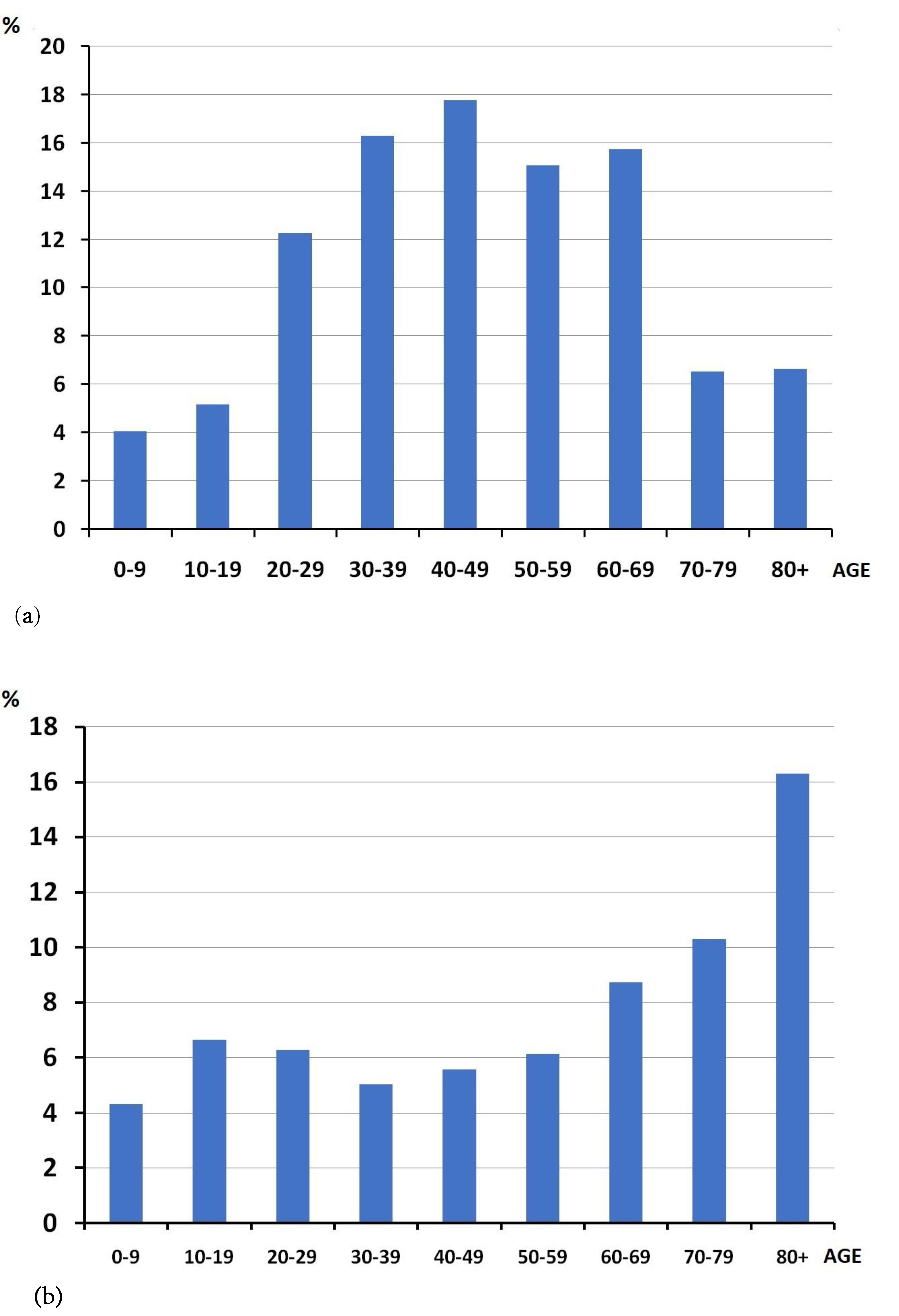
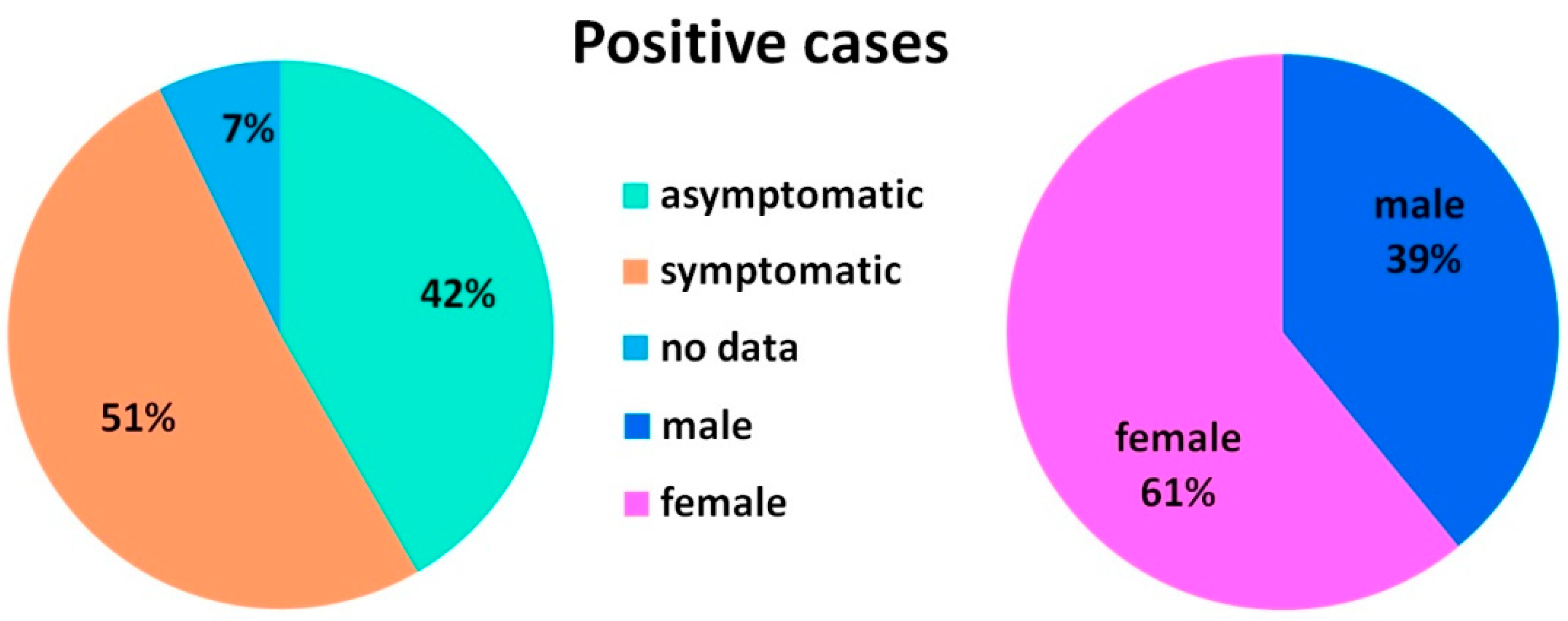
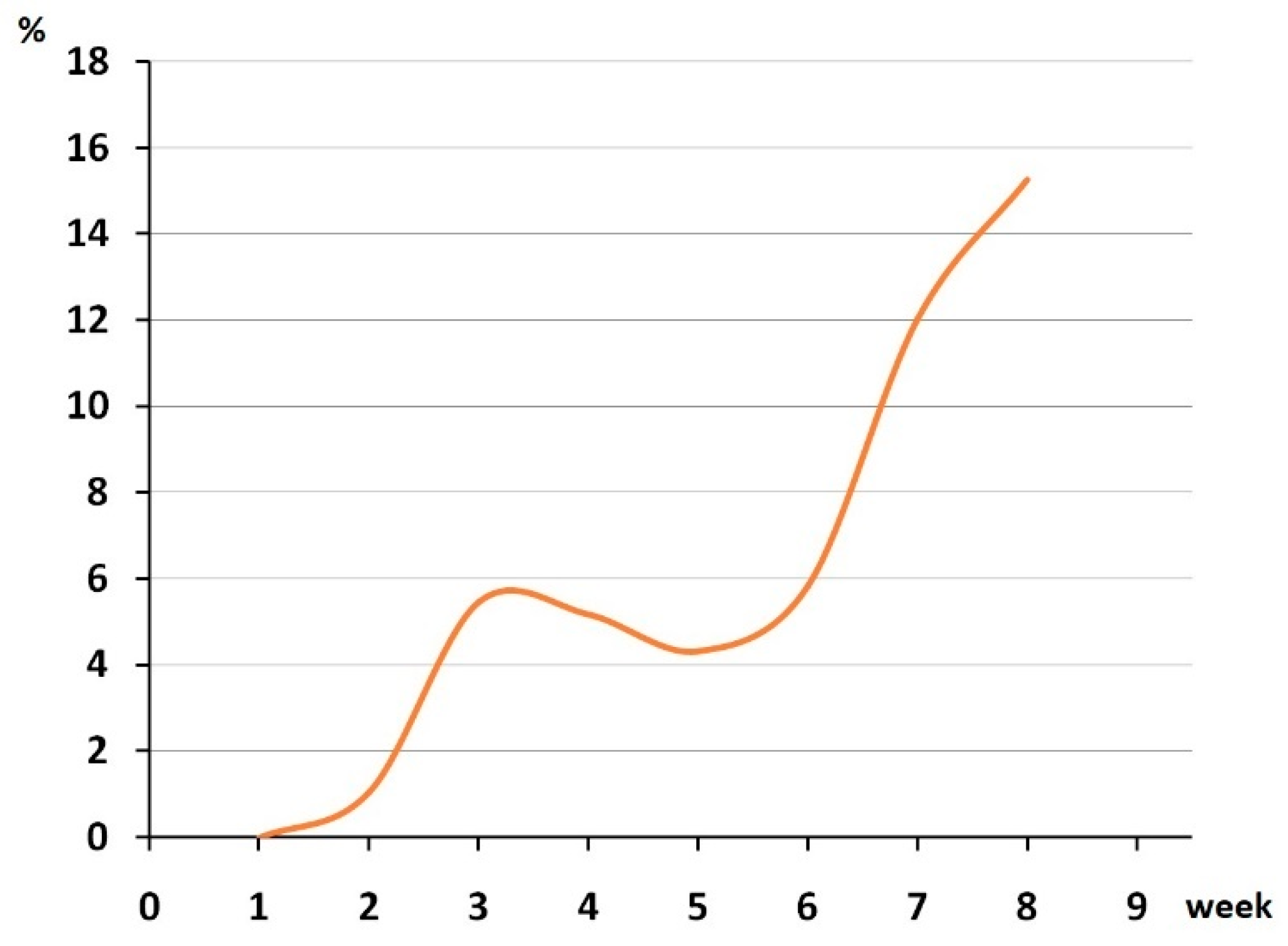
3.1. Epidemiological Analysis
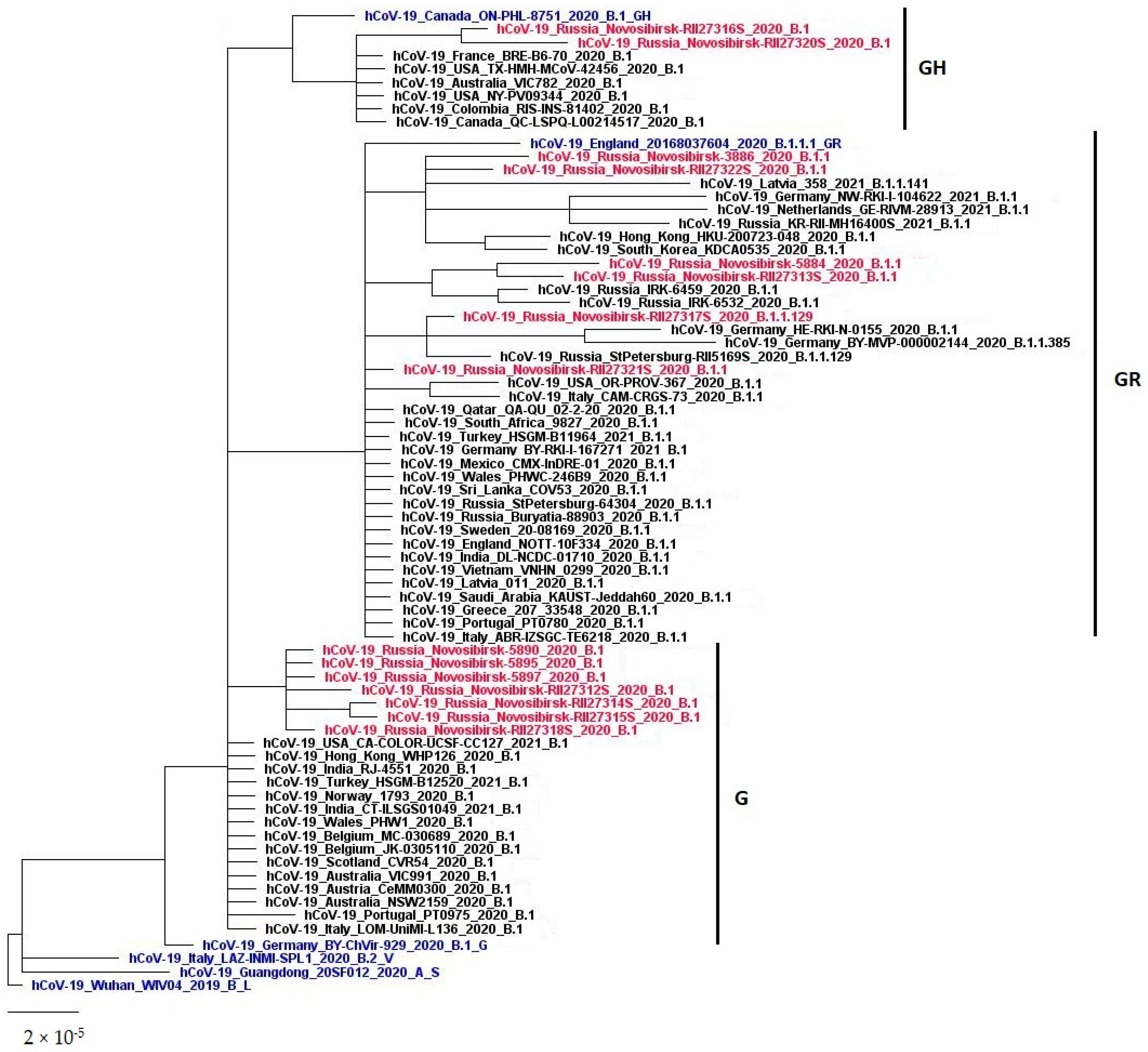
3.2. The Genetic Diversity of SARS-CoV-2 in the Novosibirsk Region at the Beginning of the Pandemic
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salzberger, B.; Buder, F.; Lampl, B.; Ehrenstein, B.; Hitzenbichler, F.; Holzmann, T.; Schmidt, B.; Hanses, F. Epidemiology of SARS-CoV-2. Infection 2021, 49, 233–239. [Google Scholar] [CrossRef]
- Bulut, C.; Kato, Y. Epidemiology of COVID-19. Turk.J.Med. Sci. 2020, 50, 563–570. [Google Scholar] [CrossRef]
- Poorolajal, J. The global pandemics are getting more frequent and severe. J. Res. Health Sci. 2021, 21, 00502. [Google Scholar] [CrossRef]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blomberg, W.R.; Meigs, D.D.; Hasan, M.; Patel, M.; et al. The natural history, pathobiology, and clinical manifestations of SARS-CoV-2 infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Scientific Brief: SARS-CoV-2 Transmission. Available online: https://www.cdc.gov/coronavirus/2019-ncov/science/science-briefs/sars-cov-2-transmission.html (accessed on 1 September 2022).
- Lee, S.; Meyler, P.; Mozel, M.; Tauh, T.; Merchant, R. Asymptomatic carriage and transmission of SARS-CoV-2: What do we know? Can. J. Anaesth. 2020, 67, 1424–1430. [Google Scholar] [CrossRef]
- Attwood, S.W.; Hill, S.C.; Aanensen, D.M.; Connor, T.R.; Pybus, O.G. Phylogenetic and phylodynamic approaches to understanding and combating the early SARS-CoV-2 pandemic. Nat. Rev. Genet. 2022, 23, 547–562. [Google Scholar] [CrossRef]
- Awadasseid, A.; Wu, Y.; Tanaka, Y.; Zhang, W. SARS-CoV-2 variants evolved during the early stage of the pandemic and effects of mutations on adaptation in Wuhan populations. Int. J. Biol. Sci. 2021, 17, 97–106. [Google Scholar] [CrossRef]
- Komissarov, A.B.; Safina, K.R.; Garushyants, S.K.; Fadeev, A.V.; Sergeeva, M.V.; Ivanova, A.A.; Danilenko, D.M.; Lioznov, D.; Shneider, O.V.; Shvyrev, N.; et al. Genomic epidemiology of the early stages of the SARS-CoV-2 outbreak in Russia. Nat. Commun. 2021, 12, 649. [Google Scholar] [CrossRef]
- Popova, A.Y.; Smirnov, V.S.; Ezhlova, E.B.; Mel’nikova, A.A.; Samoilova, L.V.; Lyalina, L.V.; Semenova, E.V.; Gurskiy, M.A.; Aksenova, E.A.; Arbuzova, T.B.; et al. Herd immunity to SARS-CoV-2 in the Novosibirsk Region population amid the COVID-19 pandemic. Probl.Virol. 2021, 66, 299–309. (In Russian) [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Di Giallonardo, F.; Duchene, S.; Puglia, I.; Curini, V.; Profeta, F.; Cammà, C.; Marcacci, M.; Calistri, P.; Holmes, E.C.; Lorusso, A. Genomic epidemiology of the first wave of SARS-CoV-2 in Italy. Viruses 2020, 12, 1438. [Google Scholar] [CrossRef]
- D’Ardes, D.; Boccatonda, A.; Rossi, I.; Pontolillo, M.; Cocco, G.; Schiavone, C.; Santilli, F.; Guagnano, M.T.; Bucci, M.; Cipollone, F. Long-term positivity to SARS-CoV-2: A clinical case of COVID-19 with persistent evidence of infection. Eur. J. Case Rep.Intern. Med. 2020, 7, 001707. [Google Scholar] [CrossRef]
- Tan, J.; Liu, S.; Zhuang, L.; Chen, L.; Dong, M.; Zhang, J.; Xin, Y. Transmission and clinical characteristics of asymptomatic patients with SARS-CoV-2 infection. Future Virol. 2020, 15, 373–380. [Google Scholar] [CrossRef]
- Nishiura, H.; Kobayashi, T.; Miyama, T.; Suzuki, A.; Jung, S.-M.; Hayashi, K.; Kinoshita, R.; Yang, Y.; Yuan, B.; Akhmetzhanov, A.R.; et al. Estimation of the asymptomatic ratio of novel coronavirus infections (COVID-19). Int. J. Infect. Dis. 2020, 94, 154–155. [Google Scholar] [CrossRef]
- Hu, Z.; Song, C.; Xu, C.; Jin, G.; Chen, Y.; Xu, X.; Ma, H.; Chen, W.; Lin, Y.; Zheng, Y.; et al. Clinical characteristics of 24 asymptomatic infections with COVID-19 screened among close contacts in Nanjing, China. Sci. China Life Sci. 2020, 63, 706–711. [Google Scholar] [CrossRef]
- B.1.1.129 Lineage Report. Available online: https://outbreak.info/situation-reports?pango=B.1.1.129 (accessed on 31 August 2022).
- Borisova, A.A.; Popova, N.D.; Kychkina, A.I.; Rozhina, A.A.; Fedulova, A.G. Epidemiological characteristics of the dynamics of a new coronavirus infection (COVID-19) in the republic of Sakha (Yakutia). Int. Res. J. 2022, 1, 84–87. (In Russian) [Google Scholar] [CrossRef]
- Ivanenko, A.V.; Soloviev, D.V.; Volkova, N.A.; Glinenko, V.M.; Smirnova, O.A.; Shvedova, I.S.; Bakhmutskaya, E.V.; Melnikov, D.A. Epidemiological features of the incidence of COVID-19 in Moscow in the period from March 1 to August 31, 2020. Public Health Life Environ.—PH&LE 2021, 3, 57–62. (In Russian) [Google Scholar] [CrossRef]
- Ochkasova, Y.V.; Korotkov, V.V.; Saveliev, S.I.; Zubchonok, N.V.; Shchukina, I.A.; Yarkovskaya, I.V.; Khodyakova, I.A.; Bondarev, V.A. Epidemiological features of the novel coronavirus disease in the Lipetsk region in 2020. Public Health Life Environ.—PH&LE 2021, 29, 63–68. (In Russian) [Google Scholar] [CrossRef]
- Popova, A.Y.; Ezhlova, E.B.; Melnikova, A.A.; Smirnov, V.S.; Lyalina, L.V.; Ermakov, A.V.; Solomashchenko, N.I.; Kovalchuk, I.V.; Vasilenko, E.A.; Romanenko, E.N.; et al. Characteristic of herd immunity among the population of Stavropol region amid the COVID-19 epidemic. J. Infectol. 2021, 13, 79–89. (In Russian) [Google Scholar] [CrossRef]
- COVID-19 Dashboard by the Center for Systems Science and Engineering (CSSE) at Johns Hopkins University (JHU). ArcGIS. Johns Hopkins University. Available online: https://coronavirus.jhu.edu/map.html (accessed on 1 September 2022).
- Lai, A.; Bergna, A.; Caucci, S.; Clementi, N.; Vicenti, I.; Dragoni, F.; Cattelan, A.M.; Menzo, S.; Pan, A.; Callegaro, A.; et al. Molecular tracing of SARS-CoV-2 in Italy in the first three months of the epidemic. Viruses 2020, 12, 798. [Google Scholar] [CrossRef]
- Sekizuka, T.; Itokawa, K.; Hashino, M.; Kawano-Sugaya, T.; Tanaka, R.; Yatsu, K.; Ohnishi, A.; Goto, K.; Tsukagoshi, H.; Ehara, H.; et al. A genome epidemiological study of SARS-CoV-2 introduction into Japan. mSphere 2020, 5, e00786-20. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.D.S.; Mishra, S.; Crispim, M.A.E.; Sales, F.C.S.; Hawryluk, I.; McCrone, J.T.; et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Tikhonov, D.G. Specifics of COVID-19 in Siberia. Sib. Res. 2021, 1, 38–41. [Google Scholar] [CrossRef]
- Bertakis, K.D.; Azari, R.; Helms, L.J.; Callahan, E.J.; Robbins, J.A. Gender differences in the utilization of health care services. J. Fam.Pract. 2000, 49, 147–152. [Google Scholar]
- Population of the Russian Federation by Gender and Age. Available online: https://rosstat.gov.ru/storage/mediabank/Bul_chislen_nasel-pv_01-01-2022.pdf (accessed on 1 September 2022).
- Russo, C.; Morello, G.; Malaguarnera, R.; Piro, S.; Furno, D.L.; Malaguarnera, L. Candidate genes of SARS-CoV-2 gender susceptibility. Sci. Rep. 2021, 11, 21968. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2021, 19, 141–154. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (Years) | Number of Tests | Positive |
|---|---|---|
| 0–6 | 544 | 21 |
| 0–9 | 833 | 36 |
| 10–19 | 693 | 46 |
| 20–29 | 1738 | 109 |
| 30–39 | 2884 | 145 |
| 40–49 | 2833 | 158 |
| 50–59 | 2189 | 134 |
| 60–69 | 1604 | 140 |
| 70–79 | 563 | 58 |
| 80+ | 362 | 59 |
| Total | 13,699 | 889 |
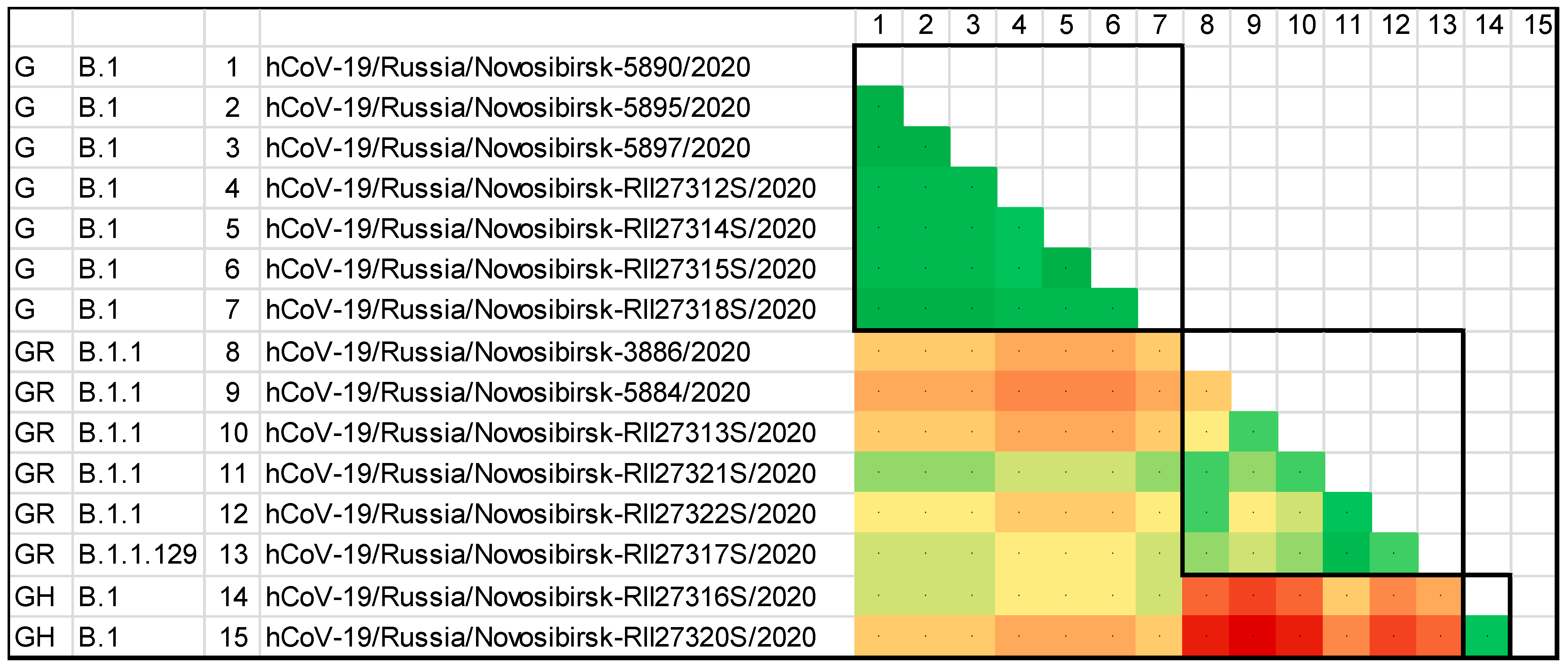
| Name | Pango | GISAID | Date | Sex | Age | Symptoms | Contact with an Infected Person | |
|---|---|---|---|---|---|---|---|---|
| 1 | hCoV-19/Russia/Novosibirsk-3886/2020 | B.1.1 | GR | 14 April 2020 | Female | 47 | Yes | |
| 2 | hCoV-19/Russia/Novosibirsk-5897/2020 | B.1 | G | 25 April 2020 | Male | 77 | Yes | |
| 3 | hCoV-19/Russia/Novosibirsk-5895/2020 | B.1 | G | 25 April 2020 | Female | 57 | Yes | |
| 4 | hCoV-19/Russia/Novosibirsk-5890/2020 | B.1 | G | 25 April 2020 | Female | 64 | Yes | |
| 5 | hCoV-19/Russia/Novosibirsk-5884/2020 | B.1.1 | GR | 26 April 2020 | Female | 46 | Yes | |
| 6 | hCoV-19/Russia/Novosibirsk-RII27318S/2020 | B.1 | G | 29 April 2020 | Female | 59 | No | Yes |
| 7 | hCoV-19/Russia/Novosibirsk-RII27315S/2020 | B.1 | G | 29 April 2020 | Male | 53 | Yes | |
| 8 | hCoV-19/Russia/Novosibirsk-RII27314S/2020 | B.1 | G | 29 April 2020 | Female | 47 | Yes | |
| 9 | hCoV-19/Russia/Novosibirsk-RII27313S/2020 | B.1.1 | GR | 29 April 2020 | Female | 68 | No | Yes |
| 10 | hCoV-19/Russia/Novosibirsk-RII27312S/2020 | B.1 | G | 29 April 2020 | Female | 70 | No | No 1 |
| 11 | hCoV-19/Russia/Novosibirsk-RII27317S/2020 | B.1.1.129 | GR | 30 April 2020 | Female | 66 | Yes | |
| 12 | hCoV-19/Russia/Novosibirsk-RII27316S/2020 | B.1 | GH | 4 April 2020 | Male | 47 | Yes | |
| 13 | hCoV-19/Russia/Novosibirsk-RII27322S/2020 | B.1.1 | GR | 7 April 2020 | Female | 41 | Yes | |
| 14 | hCoV-19/Russia/Novosibirsk-RII27321S/2020 | B.1.1 | GR | 7 May 2020 | Female | 60 | No | Yes |
| 15 | hCoV-19/Russia/Novosibirsk-RII27320S/2020 | B.1 | GH | 7 May 2020 | Female | 47 | No | No 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palyanova, N.; Sobolev, I.; Alekseev, A.; Glushenko, A.; Kazachkova, E.; Markhaev, A.; Kononova, Y.; Gulyaeva, M.; Adamenko, L.; Kurskaya, O.; et al. Genomic and Epidemiological Features of COVID-19 in the Novosibirsk Region during the Beginning of the Pandemic. Viruses 2022, 14, 2036. https://doi.org/10.3390/v14092036
Palyanova N, Sobolev I, Alekseev A, Glushenko A, Kazachkova E, Markhaev A, Kononova Y, Gulyaeva M, Adamenko L, Kurskaya O, et al. Genomic and Epidemiological Features of COVID-19 in the Novosibirsk Region during the Beginning of the Pandemic. Viruses. 2022; 14(9):2036. https://doi.org/10.3390/v14092036
Chicago/Turabian StylePalyanova, Natalia, Ivan Sobolev, Alexander Alekseev, Alexandra Glushenko, Evgeniya Kazachkova, Alexander Markhaev, Yulia Kononova, Marina Gulyaeva, Lubov Adamenko, Olga Kurskaya, and et al. 2022. "Genomic and Epidemiological Features of COVID-19 in the Novosibirsk Region during the Beginning of the Pandemic" Viruses 14, no. 9: 2036. https://doi.org/10.3390/v14092036
APA StylePalyanova, N., Sobolev, I., Alekseev, A., Glushenko, A., Kazachkova, E., Markhaev, A., Kononova, Y., Gulyaeva, M., Adamenko, L., Kurskaya, O., Bi, Y., Xin, Y., Sharshov, K., & Shestopalov, A. (2022). Genomic and Epidemiological Features of COVID-19 in the Novosibirsk Region during the Beginning of the Pandemic. Viruses, 14(9), 2036. https://doi.org/10.3390/v14092036