
Rapid Quantification of Infectious Cucumber green mottle mosaic virus in Watermelon Tissues by PMA Coupled with RT-qPCR

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and RNA Extraction
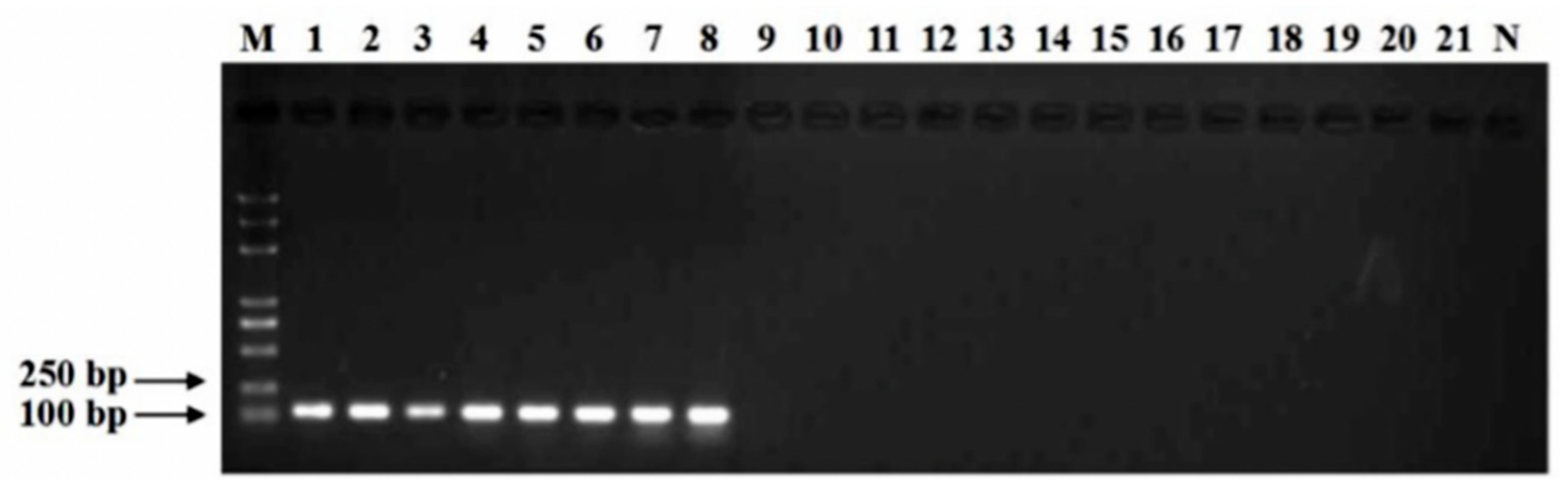
2.2. Primer Design and RT-PCR Amplificati on
2.3. RT-qPCR Conditions and Quantification Standards
2.4. Preparation of Virus Suspensions
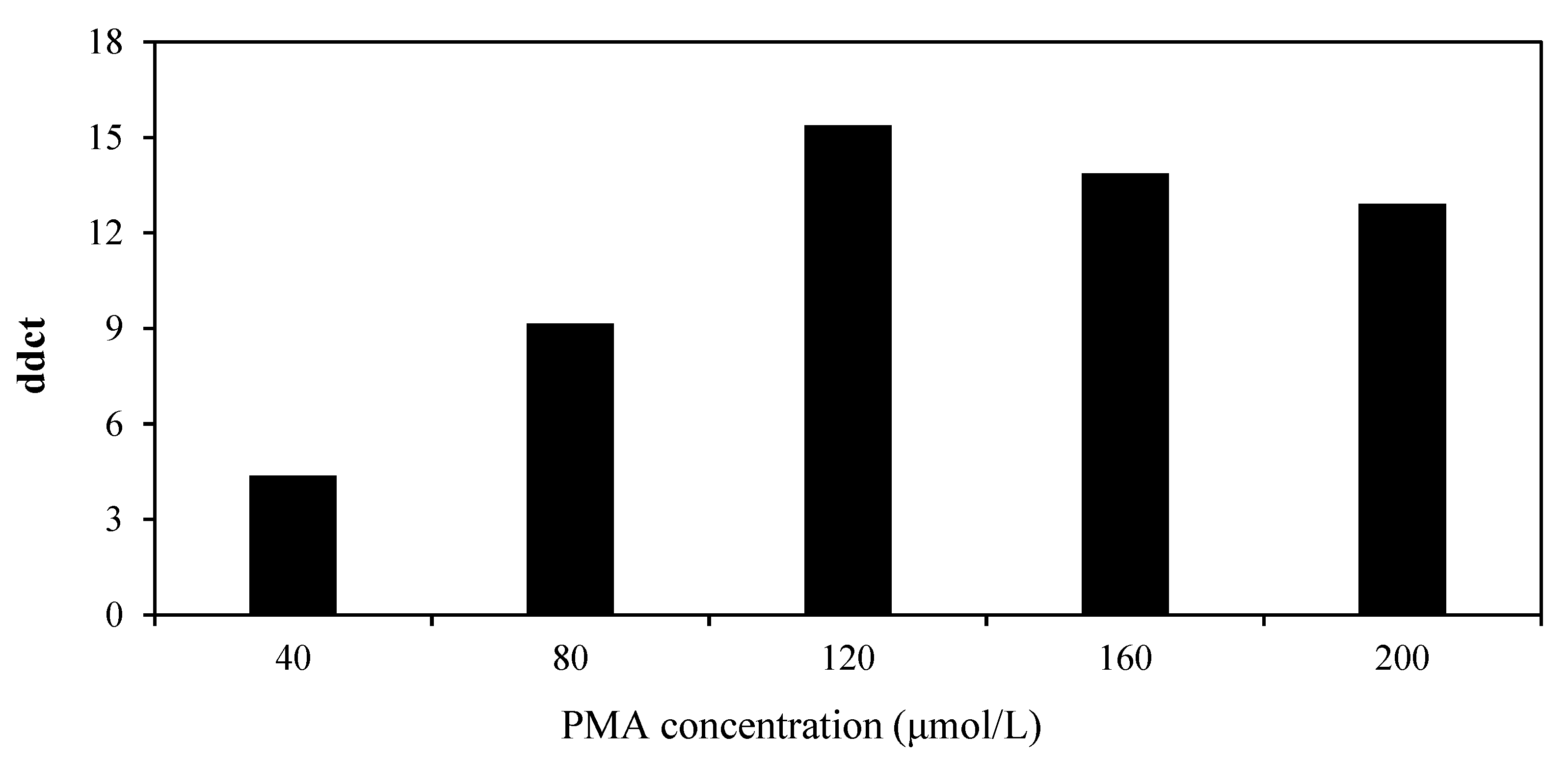
2.5. Optimization of PMA Concentration
2.6. Detection of the Defined Ratio of Mixed Virus Suspension
2.7. Sampling of Naturally Infected Watermelon Tissues
2.8. Detection of CGMMV from Naturally Infected Watermelon Tissues
2.9. Statistical Analysis
3. Results
3.1. Specificity of Primers
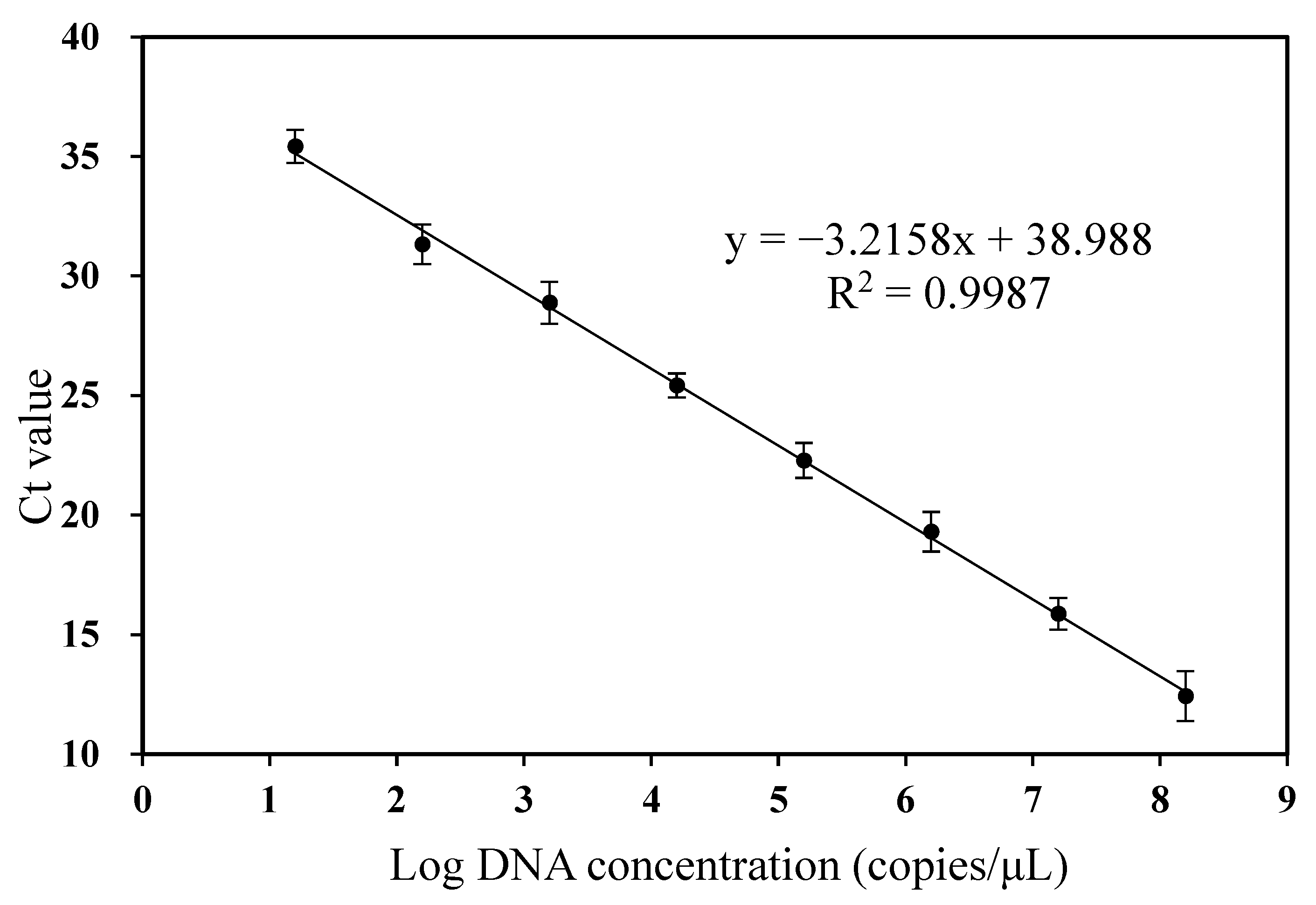
3.2. Standard Curve and Sensitivity of RT-qPCR
3.3. Optimization of the PMA-RT-qPCR System
3.4. Verification and Application of the PMA System
3.5. Application of PMA-RT-qPCR to Naturally Infected Watermelon Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Celix, A. First report of Cucumber green mottle mosaic tobamovirus infecting greenhouse-grown cucumber in Spain. Plant Dis. 1996, 80, 1303. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, G.F.; Li, M.F. Occurrence of Cucumber green mottle mosaic virus on Cucurbitaceous plants in China. Plant Dis. 2009, 93, 200. [Google Scholar] [CrossRef] [PubMed]
- Kim, O.; Mizutani, T.; Natsuaki, K.; Lee, K.; Soe, K. First report and the genetic variability of Cucumber green mottle mosaic virus occurring on bottle gourd in Myanmar. J. PhytoPathol. 2010, 158, 572–575. [Google Scholar] [CrossRef]
- Antignus, Y.; Pearlsman, M.; Rachel, B.Y.; Cohen, S. Occurrence of a variant of Cucumber green mottle mosaic virus in Israel. Phytoparasitica 1990, 18, 50–56. [Google Scholar] [CrossRef]
- Ainsworth, G.C. Mosaic disease of cucumber. Ann. Appl. Biol. 1935, 22, 55–67. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.A.; Wang, X.F.; Zhou, G.H. Molecular characterization and distribution of Cucumber green mottle mosaic virus in China. J. PhytoPathol. 2009, 157, 393–399. [Google Scholar] [CrossRef]
- Ling, K.S.; Li, R.; Zhang, W. First report of Cucumber green mottle mosaic virus infecting greenhouse cucumber in Canada. Plant Dis. 2014, 98, 701. [Google Scholar] [CrossRef]
- Sharma, P.; Verma, R.K.; Mishra, R.; Sahu, A.K.; Choudhary, D.K.; Gaur, R.K. First report of Cucumber green mottle mosaic virus association with the leaf green mosaic disease of a vegetable crop, Luffa acutangula. Acta Virol. 2014, 58, 299–300. [Google Scholar] [CrossRef]
- Tian, T.; Posis, K.; Maroon-Lango, C.J.; Mavrodieva, V.; Falk, B.W. First report of Cucumber green mottle mosaic virus on melon in the United States. Plant Dis. 2014, 98, 1163. [Google Scholar] [CrossRef]
- Cho, S.Y.; Kim, Y.S.; Yong, H.J. First report of Cucumber green mottle mosaic virus infecting Heracleum moellendorffii in Korea. Plant Dis. 2015, 99, 897. [Google Scholar] [CrossRef]
- Li, R.; Zheng, Y.; Fei, Z.; Ling, K.S. First complete genome sequence of an emerging Cucumber green mottle mosaic virus isolates in North America. Genome Announc. 2015, 3, e00452-15. [Google Scholar] [CrossRef] [PubMed]
- Borodynsko-Filas, N.; Minicka, J.; Hasiow-Jaroszewska, B. The occurrence of Cucumber green mottle mosaic virus infecting greenhouse in Poland. Plant Dis. 2017, 101, 1336. [Google Scholar] [CrossRef]
- Kehoe, M.A.; Jones, R.; Coutts, B.A. First complete genome sequence of Cucumber green mottle mosaic virus isolated from Australia. Genome Announc. 2017, 5, e00036-17. [Google Scholar] [CrossRef] [PubMed]
- Li, J.X.; Liu, S.S.; Gu, Q.S. Transmission efficiency of Cucumber green mottle mosaic virus via seeds, soil, pruning and irrigation water. J. PhytoPathol. 2016, 164, 300–309. [Google Scholar] [CrossRef]
- Darzi, E.; Smith, E.; Shargil, D.; Lachman, O.; Dombrovsky, A. The honeybee Apis mellifera contributes to Cucumber green mottle mosaic virus spread via pollination. Plant Pathol. 2017, 67, 244–251. [Google Scholar] [CrossRef]
- Reingold, V.; Lachman, O.; Belausov, E.; Koren, A.; Mor, N.; Dombrovsky, A. Epidemiological study of Cucumber green mottle mosaic virus in greenhouses enables reduction of disease damage in cucurbit production. Ann. Appl. Biol. 2016, 168, 29–40. [Google Scholar] [CrossRef]
- Dombrovsky, A.; Tran-Nguyen, L.T.T.; Jones, R.A.C. Cucumber green mottle mosaic virus: Rapidly increasing global distribution, etiology, epidemiology, and management. Annu. Rev. PhytoPathol. 2017, 55, 231–256. [Google Scholar] [CrossRef]
- Wu, H.J.; Qin, B.X.; Chen, H.Y.; Peng, B.; Cai, J.H.; Gu, Q.S. The rate of seed contamination and transmission of Cucumber green mottle mosaic virus in watermelon and melon. Sci. Agric. Sin. 2011, 44, 1527–1532. [Google Scholar]
- Sui, X.; Li, R.; Shamimuzzaman, M.; Wu, Z.; Ling, K.S. Understanding the transmissibility of Cucumber green mottle mosaic virus in watermelon seeds and seed health assays. Plant Dis. 2019, 103, 1126–1131. [Google Scholar] [CrossRef]
- Reingold, V.; Lachman, O.; Blaosov, E.; Dombrovsky, A. Seed disinfection treatments do not sufficiently eliminate the infectivity of Cucumber green mottle mosaic virus (CGMMV) on cucurbit seeds. Plant Pathol. 2015, 64, 245–255. [Google Scholar] [CrossRef]
- Smith, E.; Luria, N.; Reingold, V.; Frenkel, O.; Dombrovsky, A. Aspects in tobamovirus management in modern agriculture: Cucumber green mottle mosaic virus. Acta Hortic. 2019, 1257, 1–8. [Google Scholar] [CrossRef]
- Lee, H.I.; Kim, J.H.; Yea, M.C. Immunocapture RT-PCR for detection of seed-borne viruses on Cucurbitaceae crops. Adv. Mat. Res. 2010, 16, 121–124. [Google Scholar]
- Shang, H.; Xie, Y.; Zhou, X.; Qian, Y.; Wu, J. Monoclonal antibody-based serological methods for detection of Cucumber green mottle mosaic virus. Virol. J. 2011, 8, 228. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Wei, Q.W.; Liu, Y.; Tan, X.Q.; Zhang, W.N.; Wu, J.Y.; Charimbu, M.K.; Hu, B.S.; Cheng, Z.B.; Yu, C.; et al. One-step reverse transcription loop-mediated isothermal amplification for the rapid detection of Cucumber green mottle mosaic virus. J. Virol. Methods 2013, 193, 583–588. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Hong, J.S.; Kim, M.J.; Choi, S.H.; Min, B.E.; Song, E.G.; Kim, H.H.; Ryu, K.H. Simultaneous multiplex PCR detection of seven cucurbit-infecting viruses. J. Virol. Methods 2014, 206, 133–139. [Google Scholar] [CrossRef]
- Lee, S.; Min, B.D.; Lee, J.Y.; Shin, Y.G.; Lee, S.H.; Jeong, W.H. Development of diagnostic PCR system for three seed transmitted quarantine viruses associated with Cucurbitaceae. Microbiol. Biotechnol. Lett. 2015, 43, 79–83. [Google Scholar] [CrossRef]
- Hasiow-Jaroszewska, B.; Budzynska, D.; Borodynko-Filas, N. Genetic diversity of the Cucumber green mottle mosaic virus and the development of RT-LAMP assay for its detection. Plant Protect. Sci. 2019, 55, 1–7. [Google Scholar]
- Kwon, S.J.; Cho, Y.E.; Kim, M.H.; Seo, J.K. A one-step reverse-transcription loop-mediated isothermal amplification assay optimized for the direct detection of Cucumber green mottle mosaic virus in cucurbit seeds. Mol. Cell. Probes 2021, 60, 101775. [Google Scholar] [CrossRef]
- Yin, X.Y.; Li, X.; Yang, L.L.; Zheng, Q.Y.; Piao, Y.Z.; Cao, J.J. Detection of Cucumber green mottle mosaic virus in low-concentration virus-infected seeds by improved one-step pre-amplification RT-qPCR. Plant Methods 2022, 18, 70. [Google Scholar]
- Chen, H.; Zhao, W.; Gu, Q.; Chen, Q.; Lin, S.; Zhu, S. Real-time TaqMan RT-PCR assay for the detection of Cucumber green mottle mosaic virus. J. Virol. Methods 2008, 149, 326–329. [Google Scholar]
- Torre, C.; Agüero, J.; Gómez-Aix, C.; Aranda, M.A. Comparison of DAS-ELISA and qRT-PCR for the detection of cucurbit viruses in seeds. Ann. Appl. Biol. 2020, 176, 158–169. [Google Scholar] [CrossRef]
- Golpayegani, A.; Douraghi, M.; Rezaei, F.; Alimohammadi, M.; Nodehi, R.N. Propidium monoazide-quantitative polymerase chain reaction (PMA-qPCR) assay for rapid detection of viable and viable but non-culturable (VBNC) Pseudomonas aeruginosa in swimming pools. J. Environ. Health. Sci. 2019, 17, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Elizaquível, P.; Aznar, R.; Sánchez, G. Recent developments in the use of viability dyes and quantitative PCR in the food microbiology field. J. Appl. MicroBiol. 2013, 116, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mariana, F.; Nancy, J.R.; Francesc, C.; Bárbara, A.; Gustavo, A.; Jordi, M. Discrimination of infectious bacteriophage T4 virus by propidium monoazide real-time PCR. J. Virol. Methods 2010, 168, 228–232. [Google Scholar]
- Parshionikar, S.; Laseke, I.; Fout, G.S. Use of propidium monoazide in reverse transcriptase PCR to distinguish between infectious and noninfectious enteric viruses in water samples. Appl. Environ. Microb. 2010, 76, 4318–4326. [Google Scholar] [CrossRef]
- Sánchez, G.; Elizaquível, P.; Aznar, R. Discrimination of infectious hepatitis a viruses by propidium monoazide real-time RT-PCR. Food Environ. Virol. 2012, 4, 21–25. [Google Scholar] [CrossRef]
- Karim, M.R.; Fout, G.S.; Johnson, C.H.; White, K.M.; Parshionikar, S.U. Propidium monoazide reverse transcriptase PCR and RT-QPCR for detecting infectious enterovirus and norovirus. J. Virol. Methods 2015, 219, 51–61. [Google Scholar] [CrossRef]
- Lee, M.; Seo, D.J.; Seo, J.; Oh, H.; Jeon, S.B.; Ha, S.D. Detection of viable murine norovirus using the plaque assay and propidium-monoazide-combined real-time reverse transcription-polymerase chain reaction. J. Virol. Methods 2015, 221, 57–61. [Google Scholar] [CrossRef]
- Fongaro, G.; Hernández, M.; García-González, M.; Barardi, C.; Rodríguez-Lázaro, D. Propidium monoazide coupled with PCR predicts infectivity of enteric viruses in swine manure and biofertilized soil. Food Environ. Virol. 2016, 8, 79–85. [Google Scholar] [CrossRef]
- Hong, W.; Xiong, J.; Nyaruaba, R.; Li, J.; Muturi, E.; Liu, H.; Yu, J.; Yang, H.; Wei, H. Rapid determination of infectious SARS-CoV-2 in PCR-positive samples by SDS-PMA assisted RT-qPCR. Sci. Total Env. 2021, 797, 149085. [Google Scholar] [CrossRef]
- Terio, V.; Lorusso, P.; Mottola, A.; Buonavoglia, C.; Pinto, A.D. Norovirus detection in ready-to-eat salads by propidium monoazide real time RT-PCR assay. Appl. Sci. 2020, 10, 5176. [Google Scholar] [CrossRef]
- Fronhoffs, S.; Totzke, G.; Stier, S.; Werner, N.; Rothe, M.; Bruning, T.; Koch, B.; Sachinidis, A.; Vetter, H.; Ko, Y. A method for the rapid construction of cRNA standard curves in quantitative real-time reverse transcription polymerase chain reaction. Mol. Cell Probes. 2002, 16, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.L.; Qi, X.H.; Han, Z.Y.; Guo, Y.B.; Wang, Y.N.; Hu, T.L.; Wang, L.M.; Cao, K.Q.; Wang, S.T. Latent infection of Valsa mali in the seeds, seedlings and twigs of crabapple and cpple trees is a potential inoculum source of valsa canker. Sci. Rep. 2019, 9, 7738. [Google Scholar] [CrossRef] [PubMed]
- Chai, A.L.; Ben, H.Y.; Guo, W.T.; Shi, Y.X.; Xie, X.W.; Li, L.; Li, B.J. Quantification of viable cells of Pseudomonas syringae pv. tomato in tomato seed using propidium monoazide and a real-time PCR assay. Plant Dis. 2020, 104, 2225–2232. [Google Scholar] [CrossRef]
- Jones, P.; Angood, S.B.; Carpenter, J.M. Melon rugose mosaic virus, the cause of a disease of watermelon and sweet melon. Ann. Appl. Biol. 1986, 108, 303–307. [Google Scholar] [CrossRef]
- Zhang, Y.B.; Wang, Y.J.; Xie, Z.K.; Wang, R.Y.; Guo, Z.H.; Werf, W.; Wang, L. Purification and immuno-gold labeling of lily mottle virus from lily leaves. J. Virol. Methods 2016, 232, 33–38. [Google Scholar] [CrossRef]
- Han, S.N.; Jiang, N.; Lv, Y.Q.; Kan, Y.M.; Hao, J.J.; Li, J.Q.; Luo, L.X. Detection of Clavibacter michiganensis subsp. michiganensis in viable but nonculturable state from tomato seed using improved qPCR. PLoS ONE 2018, 13, e0196525. [Google Scholar] [CrossRef]
- Chang, K.S.; Han, K.S.; Lee, J.H.; Bae, D.W.; Kim, D.K.; Kim, H.K. Isolation and characterization of watermelon isolate of Cucumber green mottle mosaic virus (CGMMV-HY1) from watermelon plants with severe mottle mosaic symptoms. Plant Pathol. J. 2005, 21, 167–171. [Google Scholar]
- Quijada, N.M.; Gislaine, F.; Barardi, C.; Marta, H.; David, R.L. Propidium monoazide integrated with qPCR enables the detection and enumeration of infectious enteric RNA and DNA viruses in clam and fermented sausages. Front. MicroBiol. 2016, 7, 2008. [Google Scholar] [CrossRef]
- Puente, H.; Randazzo, W.; Falcó, I.; Carvajal, A.; Sánchez, G. Rapid selective detection of potentially infectious porcine epidemic diarrhea coronavirus exposed to heat treatments using viability RT-qPCR. Front. MicroBiol. 2020, 11, 1911. [Google Scholar] [CrossRef]
- Coudray-Meunier, C.; Fraisse, A.; Martin-Latil, S.; Guillier, L.; Perelle, S. Discrimination of infectious hepatitis a virus and rotavirus by combining dyes and surfactants with RT-qPCR. BMC MicroBiol. 2013, 13, 216. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Liu, H.; Meng, L.; Xing, M.; Wang, J.; Wang, C.; Chen, H.; Zheng, N. Quantitative PCR coupled with sodium dodecyl sulfate and propidium monoazide for detection of viable Staphylococcus aureus inmilk. J. Dairy Sci. 2018, 101, 4936–4943. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, H.; Liu, H.; Cai, J.; Meng, L.; Dong, L.; Zheng, N.; Wang, J.; Wang, C. Quantitative polymerase chain reaction coupled with sodium dodecyl sulfate and propidium monoazide for detection of viable Streptococcus agalactiaein milk. Front. MicroBiol. 2019, 10, 661. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Viruses | Isolate Code | Host | Tissues | Geographic Origin | PCR a |
|---|---|---|---|---|---|
| Cucumber green mottle mosaic virus (CGMMV) | TG20091901 | Watermelon | Leaves | Zhejiang Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG20091902 | Watermelon | Leaves | Zhejiang Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG20081101 | Watermelon | Leaves | Hainan Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG20081102 | Watermelon | Leaves | Hainan Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG21061411 | Watermelon | Leaves | Liaoning Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG21061412 | Watermelon | Leaves | Liaoning Province | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG21121234 | Watermelon | Leaves | Ningxia Hui Autonomous Region | + |
| Cucumber green mottle mosaic virus (CGMMV) | TG21121235 | Watermelon | Leaves | Ningxia Hui Autonomous Region | + |
| Watermelon mosaic virus (WMV) | HG16092601 | Cucumber | Leaves | Shandong province | − |
| Cucumber mosaic virus (CMV) | TG18041704 | Melon | Leaves | Shandong province | − |
| Melon mosaic virus (MMV) | TG19011201 | Melon | Leaves | Shandong province | − |
| Tobacco ringspot virus (TRSV) | HG18092107 | Cucumber | Leaves | Shandong Province | − |
| Cucurbit chlorotic yellows virus (CCYV) | HG18111501 | Cucumber | Leaves | Henan Province | − |
| Zucchini yellow mosaic virus (ZYMV) | NG17071124 | Pumpkin | Leaves | Beijing | − |
| Squash mosaic virus (SqMV) | SQ20041046 | Melon | Leaves | Beijing | − |
| Melon yellow spot virus (MYSV) | MY20075501 | Melon | Leaves | Guangxi Zhuang Autonomous Region | − |
| Melon necrotic spot virus (MNSV) | MN21060102 | Melon | Leaves | Shandong Province | − |
| Prunus necrotic ringspot virus (PNRSV) | PNR20070108 | Melon | Leaves | Shanxi Province | − |
| Papaya ringspot virus (PRSV) | PR20040101 | Papaya | Leaves | Guangdong Province | − |
| Tobacco mosaic virus (TMV) | FQ18062901 | Tomato | Leaves | Inner Mongolia Autonomous Region | − |
| Potato Y virus (PVY) | MLS18080202 | Potato | Leaves | Hebei Province | − |
| PMA Concentration (μmol·L−1) | Ct (Infectious Virus) | Ct (Inactive Virus) | ddCt 1 |
|---|---|---|---|
| 0 | 16.01 ± 0.36 c 2 | 16.38 ± 0.33 d | - |
| 40 | 16.11 ± 0.26 c | 20.40 ± 0.40 c | 4.37 |
| 80 | 16.33 ± 0.13 c | 25.41 ± 0.39 b | 9.15 |
| 120 | 16.12 ± 0.44 c | 30.43 ± 0.41 a | 15.39 |
| 160 | 17.66 ± 0.14 b | 31.46 ± 0.39 a | 13.87 |
| 200 | 19.40 ± 0.25 a | 32.24 ± 0.41 a | 12.91 |
| Sample No. | Collection Sites | Sampling Year | Tissues | RT-qPCR | PMA–RT-qPCR | Bioassay 1 DI (%) 2 | ||
|---|---|---|---|---|---|---|---|---|
| Ct Value | Total Virus (Copies/g) | Ct Value | Infectious Virus (Copies/g) | |||||
| 1 | Ningbo, Zhejiang | 2021 | Leaf | 19.97 ± 0.33 | 7.30 × 105 | 21.37 ± 0.82 | 2.68 × 105 | / |
| 2 | Ningbo, Zhejiang | 2020 | Leaf | 20.82 ± 0.19 | 3.99 × 105 | 23.27 ± 0.83 | 6.89 × 104 | / |
| 3 | Ningbo, Zhejiang | 2021 | Leaf | 27.01 ± 0.34 | 4.72 × 103 | 29.67 ± 0.80 | 7.04 × 102 | / |
| 4 | Ningbo, Zhejiang | 2020 | Leaf | 23.52 ± 0.30 | 5.75 × 104 | 28.01 ± 1.40 | 2.30 × 103 | / |
| 5 | Yingkou, Liaoning | 2020 | Leaf | 24.41 ± 1.10 | 3.05 × 104 | 27.75 ± 0.49 | 2.78 × 103 | / |
| 6 | Sanya, Hainan | 2021 | Leaf | 18.93 ± 0.66 | 1.54 × 106 | 20.48 ± 0.47 | 5.08 × 105 | / |
| 7 | Sanya, Hainan | 2020 | Leaf | 29.27 ± 0.31 | 9.37 × 102 | 30.08 ± 0.43 | 5.24 × 102 | / |
| 8 | Zhongwei, Ningxia | 2020 | Leaf | 20.85 ± 1.34 | 3.90 × 105 | 25.44 ± 2.37 | 1.46 × 104 | / |
| 9 | Ningbo, Zhejiang | 2021 | Fruit | 22.99 ± 0.72 | 8.41 × 104 | 24.99 ± 1.79 | 2.00 × 104 | / |
| 10 | Ningbo, Zhejiang | 2021 | Fruit | 28.15 ± 0.66 | 2.09 × 103 | 29.51 ± 0.41 | 7.89 × 102 | / |
| 11 | Zhongwei, Ningxia | 2020 | Fruit | 23.01 ± 1.12 | 8.27 × 104 | 27.29 ± 1.61 | 3.87 × 103 | / |
| 12 | Ningbo, Zhejiang | 2021 | Seed | 29.31 ± 0.18 | 9.09 × 102 | 31.37 ± 0.42 | 2.08 × 102 | 21.67 ± 0.07 |
| 13 | Ningbo, Zhejiang | 2021 | Seed | 24.12 ± 0.48 | 3.74 × 104 | 28.50 ± 0.46 | 1.63 × 103 | 31.11 ± 0.11 |
| The other 9 samples | >32 | 0 | >32 | 0 | / | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, A.; Wang, Q.; Kang, H.; Yan, L.; Huang, Y.; Shi, Y.; Xie, X.; Li, L.; Fan, T.; Wang, Y.; et al. Rapid Quantification of Infectious Cucumber green mottle mosaic virus in Watermelon Tissues by PMA Coupled with RT-qPCR. Viruses 2022, 14, 2046. https://doi.org/10.3390/v14092046
Chai A, Wang Q, Kang H, Yan L, Huang Y, Shi Y, Xie X, Li L, Fan T, Wang Y, et al. Rapid Quantification of Infectious Cucumber green mottle mosaic virus in Watermelon Tissues by PMA Coupled with RT-qPCR. Viruses. 2022; 14(9):2046. https://doi.org/10.3390/v14092046
Chicago/Turabian StyleChai, Ali, Quancheng Wang, Huajun Kang, Leiyan Yan, Yunping Huang, Yanxia Shi, Xuewen Xie, Lei Li, Tengfei Fan, Yuhong Wang, and et al. 2022. "Rapid Quantification of Infectious Cucumber green mottle mosaic virus in Watermelon Tissues by PMA Coupled with RT-qPCR" Viruses 14, no. 9: 2046. https://doi.org/10.3390/v14092046
APA StyleChai, A., Wang, Q., Kang, H., Yan, L., Huang, Y., Shi, Y., Xie, X., Li, L., Fan, T., Wang, Y., & Li, B. (2022). Rapid Quantification of Infectious Cucumber green mottle mosaic virus in Watermelon Tissues by PMA Coupled with RT-qPCR. Viruses, 14(9), 2046. https://doi.org/10.3390/v14092046