
An Adenovirus-Based Recombinant Herpes Simplex Virus 2 (HSV-2) Therapeutic Vaccine Is Highly Protective against Acute and Recurrent HSV-2 Disease in a Guinea Pig Model
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. FI-HSV-2 Preparation
2.2. Amplification of Recombinant Adenovirus Vaccine
2.3. Expression of Recombinant Virus
2.4. Guinea Pig Viral Challenge and Immunization
2.5. Direct ELISA for gD-Specific IgG
2.6. Neutralizing Antibody Assay
2.7. Cytokine Assay
2.8. Virus Shedding
2.9. Statistical Analysis
3. Results
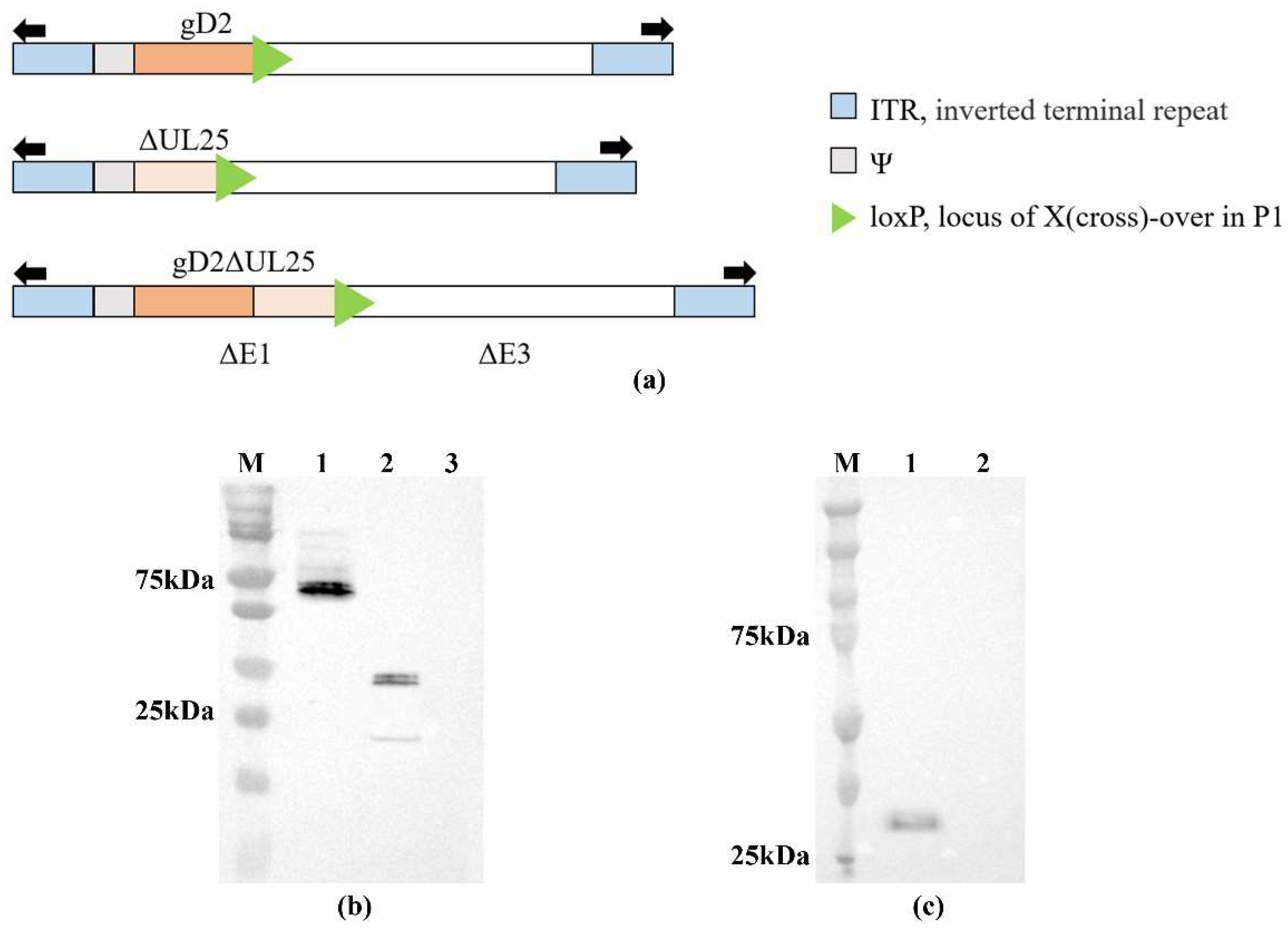
3.1. Expression of Recombinant Viruses and Proteins
3.2. Survival Rate of Guinea Pigs against LD50 HSV-2 Challenge
3.3. Disease Scores
3.4. Quantitation of Viral Shedding
3.5. Antibody Response in HSV-2 Infected Guinea Pigs
3.6. Cellular Response in HSV-2 Infected Guinea Pigs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Straface, G.; Selmin, A.; Zanardo, V.; De Santis, M.; Ercoli, A.; Scambia, G. Herpes Simplex Virus Infection in Pregnancy. Infect. Dis. Obstet. Gynecol. 2012, 2012, 385697. [Google Scholar] [CrossRef]
- WHO. 2020. Available online: https://www.who.int/news/item/01-05-2020-billions-worldwide-living-with-herpes (accessed on 12 January 2023).
- Freeman, E.E.; White, R.G.; Bakker, R.; Orroth, K.K.; Weiss, H.A.; Buve, A.; Hayes, R.J.; Glynn, J.R. Population-level effect of potential HSV2 prophylactic vaccines on HIV incidence in sub-Saharan Africa. Vaccine 2009, 27, 940–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straus, S.; Corey, L.; Burke, R.; Savarese, B.; Barnum, G.; Krause, P.; Kost, R.; Meier, J.; Sekulovich, R.; Adair, S. Placebo-controlled trial of vaccination with recombinant glycoprotein D of herpes simplex virus type 2 for immunotherapy of genital herpes. Lancet 1994, 343, 1460–1463. [Google Scholar] [CrossRef]
- Casanova, G.; Cancela, R.; Alonzo, L.; Benuto, R.; Magana, M.C.; Hurley, D.; Fishbein, E.; Lara, C.; Gonzalez, T.; Ponce, R.; et al. A double-blind study of the efficacy and safety of the ICP10deltaPK vaccine against recurrent genital HSV-2 infections. Cutis 2002, 70, 235–239. [Google Scholar] [PubMed]
- Kutinová, L.; Benda, R.; Kalos, Z.; Dbalý, V.; Votruba, T.; Kvícalová, E.; Petrovská, P.; Doutlík, S.; Kamínková, J.; Domorázková, E. Placebo-controlled study with subunit herpes simplex virus vaccine in subjects suffering from frequent herpetic recurrences. Vaccine 1988, 6, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Kimberlin, D.W.; Whitley, R.J.; Wan, W.; Powell, D.A.; Storch, G.; Ahmed, A.; Palmer, A.; Sanchez, P.J.; Jacobs, R.F.; Bradley, J.S.; et al. Oral acyclovir suppression and neurodevelopment after neonatal herpes. N. Engl. J. Med. 2011, 365, 1284–1292. [Google Scholar] [CrossRef] [Green Version]
- Koelle, D.M.; Corey, L. Herpes simplex: Insights on pathogenesis and possible vaccines. Annu. Rev. Med. 2008, 59, 381–395. [Google Scholar] [CrossRef]
- Connolly, S.A.; Landsburg, D.J.; Carfi, A.; Wiley, D.C.; Eisenberg, R.J.; Cohen, G.H. Structure-Based Analysis of the Herpes Simplex Virus Glycoprotein D Binding Site Present on Herpesvirus Entry Mediator HveA (HVEM). J. Virol. 2002, 76, 10894–10904. [Google Scholar] [CrossRef] [Green Version]
- Mettenleiter, T.C.; Klupp, B.G.; Granzow, H. Herpesvirus assembly: A tale of two membranes. Curr. Opin. Microbiol. 2006, 9, 423–429. [Google Scholar] [CrossRef]
- Ali, M.A.; Forghani, B.; Cantin, E.M. Characterization of an essential HSV-1 protein encoded by the UL25 gene reported to be involved in virus penetration and capsid assembly. Virology 1996, 216, 278–283. [Google Scholar] [CrossRef]
- Hosken, N.; McGowan, P.; Meier, A.; Koelle, D.M.; Sleath, P.; Wagener, F.; Elliott, M.; Grabstein, K.; Posavad, C.; Corey, L. Diversity of the CD8+ T-cell response to herpes simplex virus type 2 proteins among persons with genital herpes. J. Virol. 2006, 80, 5509–5515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumida, S.; Truitt, D.; Kishko, M.; Arthur, J.; Jackson, S.; Gorgone, D.; Lifton, M.; Koudstaal, W.; Pau, M.; Kostense, S.; et al. Neutralizing antibodies and CD8+ T lymphocytes both contribute to immunity to adenovirus serotype 5 vaccine vectors. J. Virol. 2004, 78, 2666–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhou, Y.; Wang, Z.; Zhang, Z.; Wang, Q.; Su, W.; Chen, Y.; Zhang, Y.; Gao, F.; Jiang, C.; et al. Evaluation of recombinant adenovirus vaccines based on glycoprotein D and truncated UL25 against herpes simplex virus type 2 in mice. Microbiol. Immunol. 2017, 61, 176–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morello, C.S.; Levinson, M.S.; Kraynyak, K.A.; Spector, D.H. Immunization with herpes simplex virus 2 (HSV-2) genes plus inactivated HSV-2 is highly protective against acute and recurrent HSV-2 disease. J. Virol. 2011, 85, 3461–3472. [Google Scholar] [CrossRef] [Green Version]
- Dib, A.; Rdc, A.; Fjb, A.; Th, B.; Dap, A.; Gc, C.; Vb, B.; Af, B. Intranasal nanoemulsion-adjuvanted HSV-2 subunit vaccine is effective as a prophylactic and therapeutic vaccine using the guinea pig model of genital herpes. Vaccine 2019, 37, 6470–6477. [Google Scholar]
- Odegard, J.M.; Flynn, P.A.; Campbell, D.J.; Robbins, S.H.; Dong, L.; Wang, K.; Ter Meulen, J.; Cohen, J.I.; Koelle, D.M. A novel HSV-2 subunit vaccine induces GLA-dependent CD4 and CD8 T cell responses and protective immunity in mice and guinea pigs. Vaccine 2016, 34, 101–109. [Google Scholar] [CrossRef]
- Skoberne, M.; Cardin, R.; Lee, A.; Kazimirova, A.; Zielinski, V.; Garvie, D.; Lundberg, A.; Larson, S.; Bravo, F.J.; Bernstein, D.I.; et al. An adjuvanted herpes simplex virus 2 subunit vaccine elicits a T cell response in mice and is an effective therapeutic vaccine in Guinea pigs. J. Virol. 2013, 87, 3930–3942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, D.; Pullum, D.; Cardin, R.; Bravo, F.; Dixon, D.; Kousoulas, K. The HSV-1 live attenuated VC2 vaccine provides protection against HSV-2 genital infection in the guinea pig model of genital herpes. Vaccine 2019, 37, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, Z.; Xu, Y.; Zhang, Z.; Hua, R.; Liu, W.; Jiang, C.; Chen, Y.; Yang, W.; Kong, W. Optimized DNA Vaccine Enhanced by Adjuvant IL28B Induces Protective Immune Responses Against Herpes Simplex Virus Type 2 in Mice. Viral Immunol. 2017, 30, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Looker, K.J.; Garnett, G.P.; Schmid, G.P. An estimate of the global prevalence and incidence of herpes simplex virus type 2 infection. Bull. World Health Organ 2008, 86, 805–812A. [Google Scholar] [CrossRef]
- Mertz, G.J. Asymptomatic Shedding of Herpes Simplex Virus 1 and 2: Implications for Prevention of Transmission. J. Infect. Dis. 2008, 198, 1098–1100. [Google Scholar] [CrossRef]
- Langenberg, A.G.; Corey, L.; Ashley, R.L.; Leong, W.P.; Straus, S.E. A prospective study of new infections with herpes simplex virus type 1 and type 2. Chiron HSV Vaccine Study Group. N. Engl. J. Med. 1999, 341, 1432–1438. [Google Scholar] [CrossRef] [PubMed]
- Tavares, F.; Cheuvart, B.; Heineman, T.; Arellano, F.; Dubin, G. Meta-analysis of pregnancy outcomes in pooled randomized trials on a prophylactic adjuvanted glycoprotein D subunit herpes simplex virus vaccine. Vaccine 2013, 31, 1759–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanberry, L.R.; Spruance, S.L.; Cunningham, A.L.; Bernstein, D.I.; Mindel, A.; Sacks, S.; Tyring, S.; Aoki, F.Y.; Slaoui, M.; Denis, M.; et al. Glycoprotein-D-adjuvant vaccine to prevent genital herpes. N. Engl. J. Med. 2002, 347, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Danko, J.; Kochel, T.; Teneza-Mora, N.; Luke, T.; Raviprakash, K.; Sun, P.; Simmons, M.; Moon, J.; De La Barrera, R.; Martinez, L.; et al. Safety and Immunogenicity of a Tetravalent Dengue DNA Vaccine Administered with a Cationic Lipid-Based Adjuvant in a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2018, 98, 849–856. [Google Scholar] [CrossRef] [PubMed]
- Mahant, A.; Guerguis, S.; Blevins, T.; Cheshenko, N.; Gao, W.; Anastos, K.; Belshe, R.; Herold, B. Herpes Simplex Virus Glycoprotein D Antibodies Fail to Elicit Antibody-Dependent Cell-Mediated Cytotoxicity: Implications for Future Vaccines. J. Infect. Dis. 2022, 226, 1489–1498. [Google Scholar] [CrossRef]
- Endresz, V.; Berencsi, K.; Gönczöl, E. An adenovirus-herpes simplex virus glycoprotein B recombinant (Ad-HSV.gB) protects mice against a vaccinia HSV.gB and HSV challenge. Acta Microbiol. Immunol. Hung. 1995, 42, 247–254. [Google Scholar]
- Johnson, D. Adenovirus vectors as potential vaccines against herpes simplex virus. Rev. Infect. Dis. 1991, 13, S912–S916. [Google Scholar] [CrossRef]
- McDermott, M.; Graham, F.; Hanke, T.; Johnson, D. Protection of mice against lethal challenge with herpes simplex virus by vaccination with an adenovirus vector expressing HSV glycoprotein B. Virology 1989, 169, 244–247. [Google Scholar] [CrossRef]
- Gallichan, W.; Johnson, D.; Graham, F.; Rosenthal, K. Mucosal immunity and protection after intranasal immunization with recombinant adenovirus expressing herpes simplex virus glycoprotein B. J. Infect. Dis. 1993, 168, 622–629. [Google Scholar] [CrossRef]
- Rollier, C.S.; Reyes-Sandoval, A.; Cottingham, M.G.; Ewer, K.; Hill, A.V. Viral vectors as vaccine platforms: Deployment in sight. Curr. Opin. Immunol. 2011, 23, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Dervillez, X.; Chentoufi, A.; Badakhshan, T.; Bettahi, I.; Benmohamed, L. Targeting the genital tract mucosa with a lipopeptide/recombinant adenovirus prime/boost vaccine induces potent and long-lasting CD8+ T cell immunity against herpes: Importance of MyD88. J. Immunol. 2012, 189, 4496–4509. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, S.; Mahairas, G.; Shaw, C.; Huang, M.; Koelle, D.; Posavad, C.; Corey, L.; Friedman, H. A Dual-Modality Herpes Simplex Virus 2 Vaccine for Preventing Genital Herpes by Using Glycoprotein C and D Subunit Antigens To Induce Potent Antibody Responses and Adenovirus Vectors Containing Capsid and Tegument Proteins as T Cell Immunogens. J. Virol. 2015, 89, 8497–8509. [Google Scholar] [CrossRef] [Green Version]
- Parr, M.B.; Parr, E.L. The role of gamma interferon in immune resistance to vaginal infection by herpes simplex virus type 2 in mice. Virology 1999, 258, 282–294. [Google Scholar] [CrossRef] [Green Version]
- Cantin, E.; Hinton, D.; Chen, J.; Openshaw, H. Gamma interferon expression during acute and latent nervous system infection by herpes simplex virus type 1. J. Virol. 1995, 69, 4898–4905. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, A.; Ruby, J.; Ramshaw, I. A case for cytokines as effector molecules in the resolution of virus infection. Immunol. Today 1993, 14, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Betts, M.R.; Casazza, J.P.; Patterson, B.A.; Waldrop, S.; Trigona, W.; Fu, T.M.; Kern, F.; Picker, L.J.; Koup, R.A. Putative Immunodominant Human Immunodeficiency Virus-Specific CD8+ T-Cell Responses Cannot Be Predicted by Major Histocompatibility Complex Class I Haplotype. J. Virol. 2000, 74, 9144–9151. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, R.; Roy, S.; Coulon, P.; Vahed, H.; Prakash, S.; Dhanushkodi, N.; Kim, G.; Fouladi, M.; Campo, J.; Teng, A.; et al. Therapeutic Mucosal Vaccination of Herpes Simplex Virus 2-Infected Guinea Pigs with Ribonucleotide Reductase 2 (RR2) Protein Boosts Antiviral Neutralizing Antibodies and Local Tissue-Resident CD4 and CD8 T Cells Associated with Protection against Recurrent Genital Herpes. J. Virol. 2019, 93, e02309-18. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, D.I. Use of the Guinea pig model of genital herpes to evaluate vaccines and antivirals: Review. Antivir. Res. 2020, 180, 104821. [Google Scholar] [CrossRef]
- Wu, Y.; Zhu, R.; Xu, L.; Li, Y.; Li, S.; Yu, H.; Li, S.; Zhu, H.; Cheng, T.; Xia, N. A novel combined vaccine based on monochimeric VLP co-displaying multiple conserved epitopes against enterovirus 71 and varicella-zoster virus. Vaccine 2017, 35, 2728–2735. [Google Scholar] [CrossRef]
- Jacquet, A.; Haumont, M.; Massaer, M.; Garcia, L.; Mazzu, P.; Daminet, V.; Gregoire, D.; Jacobs, P.; Bollen, A. Immunogenicity of a recombinant varicella-zoster virus gE-IE63 fusion protein, a putative vaccine candidate against primary infection and zoster reactivation. Vaccine 2002, 20, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Bonsignori, M.; Pollara, J.; Moody, M.; Alpert, M.; Chen, X.; Hwang, K.; Gilbert, P.; Huang, Y.; Gurley, T.; Kozink, D.; et al. Antibody-dependent cellular cytotoxicity-mediating antibodies from an HIV-1 vaccine efficacy trial target multiple epitopes and preferentially use the VH1 gene family. J. Virol. 2012, 86, 11521–11532. [Google Scholar] [CrossRef] [PubMed]
- Florese, R.; Demberg, T.; Xiao, P.; Kuller, L.; Larsen, K.; Summers, L.; Venzon, D.; Cafaro, A.; Ensoli, B.; Robert-Guroff, M. Contribution of nonneutralizing vaccine-elicited antibody activities to improved protective efficacy in rhesus macaques immunized with Tat/Env compared with multigenic vaccines. J. Immunol. 2009, 182, 3718–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialuk, I.; Whitney, S.; Andresen, V.; Florese, R.H.; Franchini, G. Vaccine induced antibodies to the first variable loop of human immunodeficiency virus type 1 gp120, mediate antibody-dependent virus inhibition in macaques. Vaccine 2012, 30, 78–94. [Google Scholar] [CrossRef] [Green Version]
- Petro, C.; González, P.; Cheshenko, N.; Jandl, T.; Khajoueinejad, N.; Bénard, A.; Sengupta, M.; Herold, B.; Jacobs, W. Herpes simplex type 2 virus deleted in glycoprotein D protects against vaginal, skin and neural disease. eLife 2015, 4, e06054. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T cell and antibody responses induced by a single dose of ChAdOx1 nCoV-19 (AZD1222) vaccine in a phase 1/2 clinical trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Liu, J.; Fang, E.; Liu, X.; Peng, Q.; Zhang, Z.; Li, M.; Liu, X.; Wu, X.; et al. Combining intramuscular and intranasal homologous prime-boost with a chimpanzee adenovirus-based COVID-19 vaccine elicits potent humoral and cellular immune responses in mice. Emerg. Microbes Infect. 2022, 11, 1890–1899. [Google Scholar] [CrossRef]
- Stanfield, B.; Kousoulas, K.G. Herpes Simplex Vaccines: Prospects of Live-Attenuated HSV Vaccines to Combat Genital and Ocular Infections. Curr. Clin. Microbiol. Rep. 2015, 2, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Halford, W. Antigenic breadth: A missing ingredient in HSV-2 subunit vaccines? Expert Rev. Vaccines 2014, 13, 691–710. [Google Scholar] [CrossRef]
- Zhang, P.; Narayanan, E.; Liu, Q.; Tsybovsky, Y.; Boswell, K.; Ding, S.; Hu, Z.; Follmann, D.; Lin, Y.; Miao, H.; et al. A multiclade env-gag VLP mRNA vaccine elicits tier-2 HIV-1-neutralizing antibodies and reduces the risk of heterologous SHIV infection in macaques. Nat. Med. 2021, 27, 2234–2245. [Google Scholar] [CrossRef]
- Laing, K.J.; Magaret, A.S.; Mueller, D.E.; Zhao, L.; Johnston, C.; De Rosa, S.C.; Koelle, D.M.; Wald, A.; Corey, L. Diversity in CD8(+) T cell function and epitope breadth among persons with genital herpes. J. Clin. Immunol. 2010, 30, 703–722. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immunogen | Immunization Mode | Immunization Dose |
|---|---|---|
| rAd-gD2ΔUL25 | i.m. | 1 × 108 PFU |
| rAd-gD2 + rAd-ΔUL25 | i.m. | 1 × 108 PFU + 1 × 108 PFU |
| rAd-gD2 | i.m. | 1 × 108 PFU |
| FI-HSV-2 | i.m. | 12.5 μg |
| Acyclovir | p.o. | 15 mg/kg |
| PBS | i.m. | 100 μL |
| Groups | 14–27 dpi | 28–41 dpi | 42–56 dpi | |||
|---|---|---|---|---|---|---|
| Recurrent Mean Lesions | Recurrent Mean Lesions | Recurrent Mean Lesions | ||||
| Score | % Reduction a | Score | % Reduction a | Score | % Reduction a | |
| PBS | 12.0 ± 3.1 | 11.0 ± 1.2 | 11.0 ± 2.2 | |||
| Acyclovir | 10.6 ± 1.4 | 11.6% | 9.3 ± 2.9 | 15.5% | 7.3 ± 1.2 | 33.6% |
| FI-HSV-2 | 11.3 ± 1.5 | 5.8% | 10.2 ± 1.6 | 0.7% | 7.7 ± 4.2 | 30.0% |
| rAd-gD2 | 10.2 ± 1.7 | 15.0% b | 6.8 ± 1.5 | 38.2% b | 3.0 ± 2.4 | 72.7% c |
| rAd-gD2 + rAd-ΔUL25 | 4.8 ± 3.8 | 60.0% c | 1.5 ± 1.1 | 86.4% d | 1.3 ± 0.5 | 88.2% c |
| rAd-gD2ΔUL25 | 3.5 ± 3.2 | 70.8% d | 1.5 ± 1.2 | 86.4% d | 1.6 ± 0.9 | 85.5% c |
| Recurrent Viral Shedding | ||||||||
|---|---|---|---|---|---|---|---|---|
| Groups | 14–27 dpi | 28–41 dpi | 42–56 dpi | Total | ||||
| Days a | Quantity b | Days | Quantity | Days | Quantity | Days | Quantity | |
| PBS | 14 | 2.5 ± 1.2 | 14 | 2.5 ± 0.7 | 12 | 3.5 ± 1.5 | 40 | 2.8 ± 1.1 |
| Acyclovir | 10 | 1.7 ± 1.0 | 5 | 1.3 ± 0.4 | 10 | 2.0 ± 0.9 | 25 | 1.7 ± 1.3 |
| FI-HSV-2 | 10 | 0.6 ± 0.4 | 9 | 1.6 ± 0.6 | 13 | 1.5 ± 0.5 | 32 | 1.2 ± 0.6 |
| rAd-gD2 | 9 | 0.9 ± 0.2 | 4 | 1.8 ± 1.3 | 13 | 1.7 ± 1.0 | 26 | 1.4 ± 1.1 |
| rAd-gD2 + rAd-ΔUL25 | 4 | 1.0 ± 0.6 e | 1 | 1.0 ± 0.5 e | 6 | 1.0 ± 0.5 d | 11 | 1.0 ± 0.6 f |
| rAd-gD2ΔUL25 | 6 | 1.2 ± 0.5 d | 1 | 1.5 ± 0.5 | 5 | 1.5 ± 1.3 c | 12 | 1.4 ± 0.7 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, M.; Yang, X.; Sun, J.; Ding, X.; Chen, Z.; Su, W.; Cai, L.; Hou, A.; Sun, B.; Gao, F.; et al. An Adenovirus-Based Recombinant Herpes Simplex Virus 2 (HSV-2) Therapeutic Vaccine Is Highly Protective against Acute and Recurrent HSV-2 Disease in a Guinea Pig Model. Viruses 2023, 15, 219. https://doi.org/10.3390/v15010219
Wan M, Yang X, Sun J, Ding X, Chen Z, Su W, Cai L, Hou A, Sun B, Gao F, et al. An Adenovirus-Based Recombinant Herpes Simplex Virus 2 (HSV-2) Therapeutic Vaccine Is Highly Protective against Acute and Recurrent HSV-2 Disease in a Guinea Pig Model. Viruses. 2023; 15(1):219. https://doi.org/10.3390/v15010219
Chicago/Turabian StyleWan, Mingming, Xiao Yang, Jie Sun, Xue Ding, Zhijun Chen, Weiheng Su, Linjun Cai, Ali Hou, Bo Sun, Feng Gao, and et al. 2023. "An Adenovirus-Based Recombinant Herpes Simplex Virus 2 (HSV-2) Therapeutic Vaccine Is Highly Protective against Acute and Recurrent HSV-2 Disease in a Guinea Pig Model" Viruses 15, no. 1: 219. https://doi.org/10.3390/v15010219