
Umbilical Cord Mesenchymal-Stem-Cell-Derived Exosomes Exhibit Anti-Oxidant and Antiviral Effects as Cell-Free Therapies
 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Antibodies and Reagents
2.3. Exosome Isolation and Purification
2.4. NanoSight Tracking Analysis
2.5. Transmission Electron Microscopy (TEM)
2.6. Oxidative Damage and Exosome Treatment
2.7. Western Blotting
2.8. Cell Proliferation Assay
2.9. Lactate Dehydrogenase (LDH) Assay
2.10. Senescence-β-Galactosidase (SA-β-Gal) Staining
2.11. qPCR Assay
2.12. Plaque Assays
2.13. Statistical Analysis
3. Results
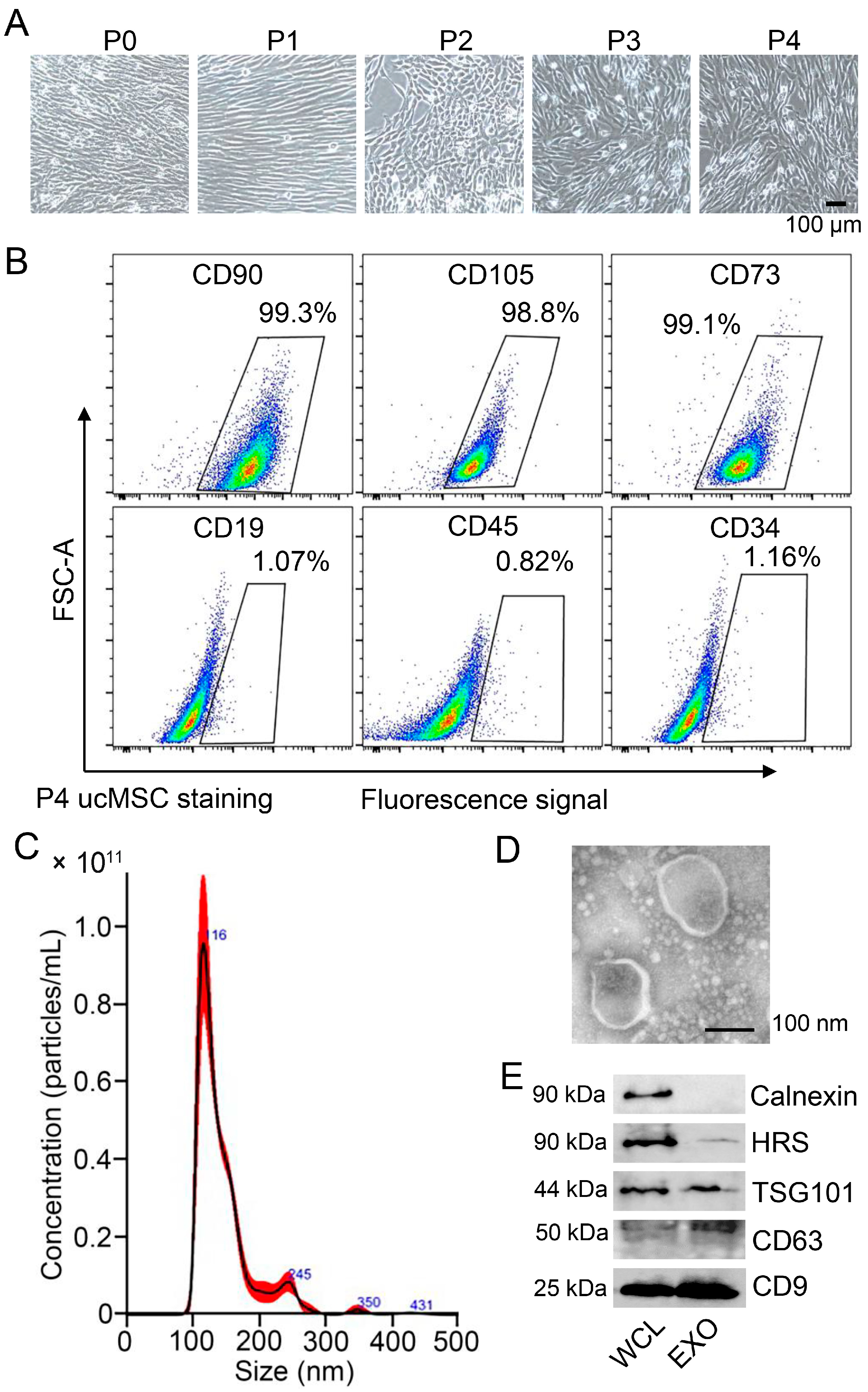
3.1. Characterization of ucMSCs and Identification of ucMSC-EXO
3.2. ucMSC-EXO Attenuate the H2O2-Induced Oxidative Damage of HSFs
3.3. ucMSC-EXO Specific Therapeutic Effects on Oxidative Damage as Indicated by Multiple Signaling Pathways
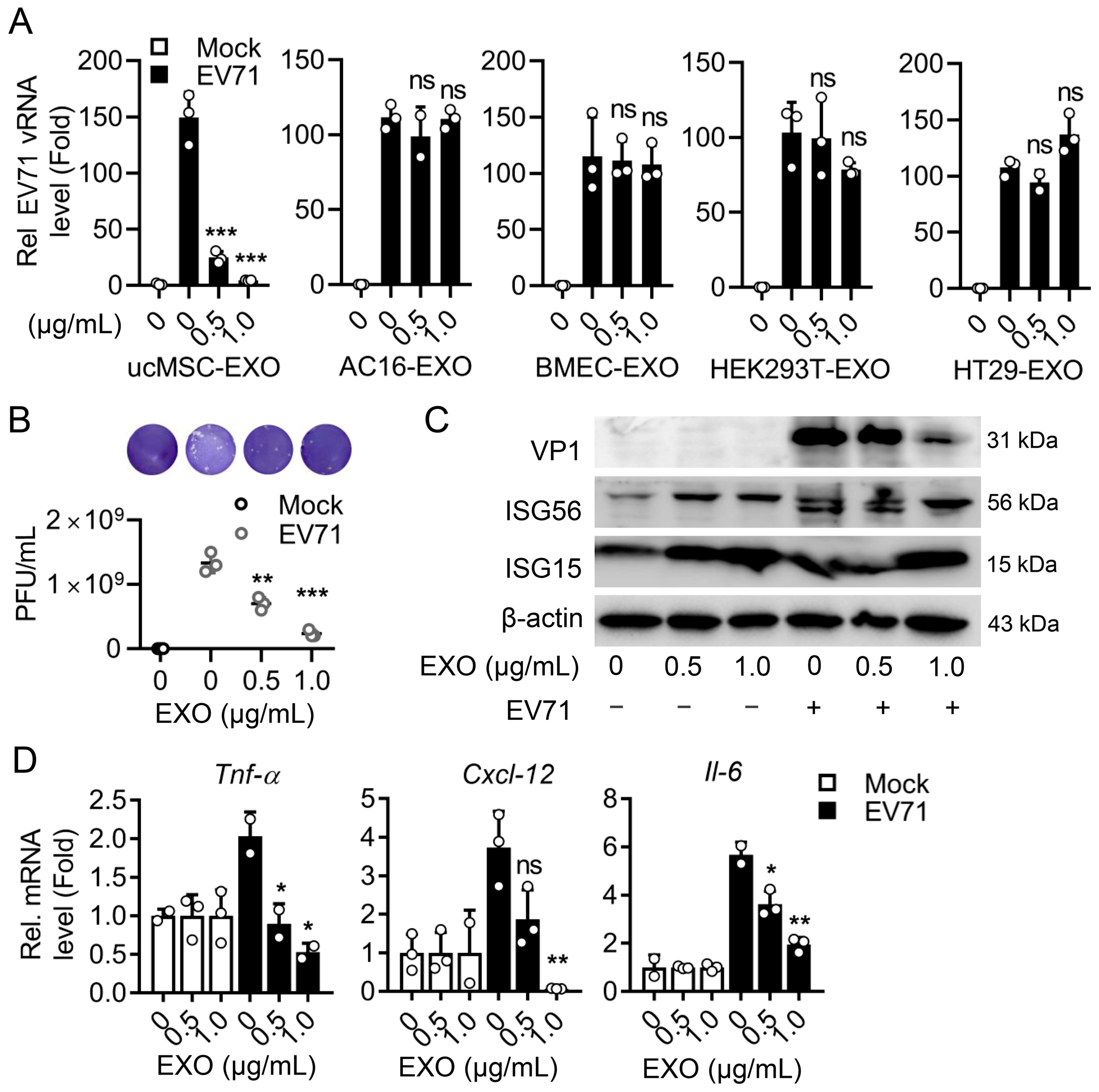
3.4. The Cell-Specific ucMSC-EXO Exert Antiviral Activity upon Enteroviral Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ROS | Reactive oxygen species |
| ucMSC | Umbilical cord mesenchymal stem cells |
| ucMSC-EXO | Exosomes derived from ucMSCs |
| HSFs | Skin fibroblasts |
| MAPK | Mitogen-activated protein kinases |
| JNK | c-Jun N-terminal kinase |
| NF-κB | Nuclear factor kappa-B |
| EV71 | Enterovirus 71 |
| CVB3 | Coxsackievirus B3 |
| ISG15 | Interferon-stimulated gene 15 |
| DDR | DNA damage response |
| SASP | Senescence-associated secretory phenotype |
| p38 MAPK | P38 mitogen-activated protein kinases |
| HFMD | Hand-foot-and-mouth disease |
| EVs | Extracellular vesicles |
| HRS | Hepatocyte growth factor receptor tyrosine kinase substrate |
| TSG101 | Tumor susceptibility gene 101 |
| Nrf2 | Nuclear factor erythroid 2-like |
| AP-1 | Activating protein 1 |
| IBD | Inflammatory bowel disease |
| SARS-CoV-2 | Severe acute respiratory syndrome coronavirus 2 |
| COVID-19 | Coronarius disease 2019 |
| PAMPs | Pathogen-associated molecular patterns |
| IFIT | IFN-induced protein with tetratricopeptide repeats |
| hBMEC | Human brain microvascular endothelial cell |
| TEM | Transmission electron microscopy |
| PVDF | Polyvinylidene fluoride |
| LDH | Lactate dehydrogenase |
| SA-β-Gal | Senescence-β-Galactosidase |
| CCK8 | Cell Counting Kit-8 |
References
- Patel, R.; Kevin Heard, L.; Chen, X.; Bollag, W.B. Aquaporins in the Skin. Adv. Exp. Med. Biol. 2017, 969, 173–191. [Google Scholar] [CrossRef]
- Feng, Y.L.; Yin, Y.X.; Ding, J.; Yuan, H.; Yang, L.; Xu, J.J.; Hu, L.Q. Alpha-1-antitrypsin suppresses oxidative stress in preeclampsia by inhibiting the p38MAPK signaling pathway: An in vivo and in vitro study. PLoS ONE 2017, 12, e0173711. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Shen, Y.; Zhu, R.; Shan, W.; Li, Y.; Yan, M.; Zhang, Y. Benzo[a]pyrene exposure promotes RIP1-mediated necroptotic death of osteocytes and the JNK/IL-18 pathway activation via generation of reactive oxygen species. Toxicology 2022, 476, 153244. [Google Scholar] [CrossRef] [PubMed]
- Yanagisawa, O.; Homma, T.; Okuwaki, T.; Shimao, D.; Takahashi, H. Effects of cooling on human skin and skeletal muscle. Eur. J. Appl. Physiol. 2007, 100, 737–745. [Google Scholar] [CrossRef]
- Douche-Aourik, F.; Berlier, W.; Feasson, L.; Bourlet, T.; Harrath, R.; Omar, S.; Grattard, F.; Denis, C.; Pozzetto, B. Detection of enterovirus in human skeletal muscle from patients with chronic inflammatory muscle disease or fibromyalgia and healthy subjects. J. Med. Virol. 2003, 71, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.X.; Zou, Y.; Liang, Q.H.; Wang, Y.E.; Liu, Y.L.; Xu, G.Q.; Fan, H.D.; Xu, Q.M.; Yang, S.L.; Yu, D. Anemoside B4 inhibits enterovirus 71 propagation in mice through upregulating 14-3-3 expression and type I interferon responses. Acta Pharmacol. Sin. 2022, 43, 977–991. [Google Scholar] [CrossRef]
- Si, Y.L.; Zhao, Y.L.; Hao, H.J.; Fu, X.B.; Han, W.D. MSCs: Biological characteristics, clinical applications and their outstanding concerns. Ageing Res. Rev. 2011, 10, 93–103. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Cao, W.; Shi, Y. Plasticity of mesenchymal stem cells in immunomodulation: Pathological and therapeutic implications. Nat. Immunol. 2014, 15, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Yin, S.; Ji, C.; Wu, P.; Jin, C.; Qian, H. Human umbilical cord mesenchymal stem cells and exosomes: Bioactive ways of tissue injury repair. Am. J. Transl. Res. 2019, 11, 1230–1240. [Google Scholar]
- Keshtkar, S.; Azarpira, N.; Ghahremani, M.H. Mesenchymal stem cell-derived extracellular vesicles: Novel frontiers in regenerative medicine. Stem Cell Res. Ther. 2018, 9, 63. [Google Scholar] [CrossRef]
- Rani, S.; Ritter, T. The Exosome—A Naturally Secreted Nanoparticle and its Application to Wound Healing. Adv. Mater. 2016, 28, 5542–5552. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Shi, H.; Yin, S.; Ji, C.; Zhang, X.; Zhang, B.; Wu, P.; Shi, Y.; Mao, F.; Yan, Y.; et al. Human Mesenchymal Stem Cell Derived Exosomes Alleviate Type 2 Diabetes Mellitus by Reversing Peripheral Insulin Resistance and Relieving beta-Cell Destruction. ACS Nano 2018, 12, 7613–7628. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Bai, L.; Li, D.; Li, J.; Luo, Z.; Yu, S.; Cao, S.; Shen, L.; Zuo, Z.; Ma, X. Bioactive molecules derived from umbilical cord mesenchymal stem cells. Acta Histochem. 2016, 118, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.; Liang, Y.; Chen, Z.; Yin, J.; Li, C.; Pan, P.; Zhang, Q.; Wu, J.; Luo, Z. Enterovirus 71 non-structural protein 3A hijacks vacuolar protein sorting 25 to boost exosome biogenesis to facilitate viral replication. Front. Microbiol. 2022, 13, 1024899. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, T.; Li, L.; Guo, Y.; Xiao, J.; Wang, J.; Du, Z.; Gao, H.; Tang, W.; Yang, L.; et al. Scrapie-Responsive Gene 1 Promotes Chondrogenic Differentiation of Umbilical Cord Mesenchymal Stem Cells via Wnt5a. Stem Cells Int. 2022, 2022, 9124277. [Google Scholar] [CrossRef]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-associated beta-galactosidase is lysosomal beta-galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef]
- Han, Y.M.; Bedarida, T.; Ding, Y.; Somba, B.K.; Lu, Q.; Wang, Q.; Song, P.; Zou, M.H. Beta-Hydroxybutyrate Prevents Vascular Senescence through hnRNP A1-Mediated Upregulation of Oct4. Mol. Cell 2018, 71, 1064–1078.e5. [Google Scholar] [CrossRef]
- Tan, D.Q.; Suda, T. Reactive Oxygen Species and Mitochondrial Homeostasis as Regulators of Stem Cell Fate and Function. Antioxid. Redox Signal 2018, 29, 149–168. [Google Scholar] [CrossRef]
- Damiani, E.; Ullrich, S.E. Understanding the connection between platelet-activating factor, a UV-induced lipid mediator of inflammation, immune suppression and skin cancer. Prog. Lipid Res. 2016, 63, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; van der Vliet, A.; Schock, B.C.; Okamoto, T.; Obermuller-Jevic, U.; Cross, C.E.; Packer, L. Ozone exposure activates oxidative stress responses in murine skin. Toxicology 2002, 179, 163–170. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Mazzoni, L.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Gonzalez-Paramas, A.M.; Santos-Buelga, C.; Quiles, J.L.; Bompadre, S.; Mezzetti, B.; et al. An anthocyanin-rich strawberry extract protects against oxidative stress damage and improves mitochondrial functionality in human dermal fibroblasts exposed to an oxidizing agent. Food Funct. 2014, 5, 1939–1948. [Google Scholar] [CrossRef]
- Hu, Z.; Yuan, Y.; Zhang, X.; Lu, Y.; Dong, N.; Jiang, X.; Xu, J.; Zheng, D. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Attenuate Oxygen-Glucose Deprivation/Reperfusion-Induced Microglial Pyroptosis by Promoting FOXO3a-Dependent Mitophagy. Oxid. Med. Cell. Longev. 2021, 2021, 6219715. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhong, Z.; Zhang, Y.; Tan, H.; Deng, B.; Meng, G. Human umbilical cord mesenchymal stem cell-derived exosomal miR-335-5p attenuates the inflammation and tubular epithelial-myofibroblast transdifferentiation of renal tubular epithelial cells by reducing ADAM19 protein levels. Stem Cell Res. Ther. 2022, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Lee, E.N.; Park, J.H.; Lee, J.K.; Cho, G.J.; Park, I.H.; Shin, O.S. Anti-Viral Activities of Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Against Human Respiratory Viruses. Front. Cell. Infect. Microbiol. 2022, 12, 850744. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Wang, H.; Bian, L.; Huang, J.; Wu, D.; Zhang, R.; Fei, F.; Chen, Y.; Xia, J. Nebulization Therapy with Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes for COVID-19 Pneumonia. Stem Cell Rev. Rep. 2022, 18, 2152–2163. [Google Scholar] [CrossRef]
- Zhang, K.; Yu, L.; Li, F.R.; Li, X.; Wang, Z.; Zou, X.; Zhang, C.; Lv, K.; Zhou, B.; Mitragotri, S.; et al. Topical Application of Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells in Combination with Sponge Spicules for Treatment of Photoaging. Int. J. Nanomed. 2020, 15, 2859–2872. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xiao, Q.; Tang, J.; Xiong, L.; Li, L. Extracellular Vesicles: Emerging Therapeutics in Cutaneous Lesions. Int. J. Nanomed. 2021, 16, 6183–6202. [Google Scholar] [CrossRef]
- Han, H.S.; Shin, J.S.; Myung, D.B.; Ahn, H.S.; Lee, S.H.; Kim, H.J.; Lee, K.T. Hydrangea serrata (Thunb.) Ser. Extract Attenuate UVB-Induced Photoaging through MAPK/AP-1 Inactivation in Human Skin Fibroblasts and Hairless Mice. Nutrients 2019, 11, 533. [Google Scholar] [CrossRef]
- Duan, X.; Wu, T.; Liu, T.; Yang, H.; Ding, X.; Chen, Y.; Mu, Y. Vicenin-2 ameliorates oxidative damage and photoaging via modulation of MAPKs and MMPs signaling in UVB radiation exposed human skin cells. J. Photochem. Photobiol. B 2019, 190, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Zhan, J.Y.; Wang, X.F.; Liu, Y.H.; Zhang, Z.B.; Wang, L.; Chen, J.N.; Huang, S.; Zeng, H.F.; Lai, X.P. Andrographolide Sodium Bisulfate Prevents UV-Induced Skin Photoaging through Inhibiting Oxidative Stress and Inflammation. Mediat. Inflamm. 2016, 2016, 3271451. [Google Scholar] [CrossRef] [PubMed]
- Kwak, C.S.; Yang, J.; Shin, C.Y.; Chung, J.H. Topical or oral treatment of peach flower extract attenuates UV-induced epidermal thickening, matrix metalloproteinase-13 expression and pro-inflammatory cytokine production in hairless mice skin. Nutr. Res. Pract. 2018, 12, 29–40. [Google Scholar] [CrossRef]
- Abbaszadeh, H.; Ghorbani, F.; Derakhshani, M.; Movassaghpour, A.; Yousefi, M. Human umbilical cord mesenchymal stem cell-derived extracellular vesicles: A novel therapeutic paradigm. J. Cell. Physiol. 2020, 235, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Qiu, W.; Xu, X.; Kang, J.; Wang, J.; Wen, Y.; Tang, X.; Yan, Y.; Qian, H.; Zhang, X.; et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate inflammatory bowel disease in mice through ubiquitination. Am. J. Transl. Res. 2018, 10, 2026–2036. [Google Scholar]
- Zhang, Q.; Bastard, P.; Effort, C.H.G.; Cobat, A.; Casanova, J.L. Human genetic and immunological determinants of critical COVID-19 pneumonia. Nature 2022, 603, 587–598. [Google Scholar] [CrossRef]
- Abdelgawad, M.; Bakry, N.S.; Farghali, A.A.; Abdel-Latif, A.; Lotfy, A. Mesenchymal stem cell-based therapy and exosomes in COVID-19: Current trends and prospects. Stem Cell Res. Ther. 2021, 12, 469. [Google Scholar] [CrossRef]
- Pestka, S. The interferons: 50 years after their discovery, there is much more to learn. J. Biol. Chem. 2007, 282, 20047–20051. [Google Scholar] [CrossRef]
- Okumura, F.; Zou, W.; Zhang, D.E. ISG15 modification of the eIF4E cognate 4EHP enhances cap structure-binding activity of 4EHP. Genes Dev. 2007, 21, 255–260. [Google Scholar] [CrossRef]
- Abbas, Y.M.; Pichlmair, A.; Gorna, M.W.; Superti-Furga, G.; Nagar, B. Structural basis for viral 5’-PPP-RNA recognition by human IFIT proteins. Nature 2013, 494, 60–64. [Google Scholar] [CrossRef]
- Kim, Y.; Park, N.; Rim, Y.A.; Nam, Y.; Jung, H.; Lee, K.; Ju, J.H. Establishment of a complex skin structure via layered co-culture of keratinocytes and fibroblasts derived from induced pluripotent stem cells. Stem Cell Res. Ther. 2018, 9, 217. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primers |
|---|---|
| Tnf-α | F: 5′-CTGCACTTTGGAGTGATCGG |
| R: 5′-AGGGTTTGCTACAACATGGG | |
| Cxcl-12 | F: 5′-TCTTCGAAAGCCATGTTGCC |
| R: 5′-CTTCGGGTCAATGCACACTT | |
| Il-6 | F: 5′-AATGAGGAGACTTGCCTGGT |
| R: 5′-GCAGGAACTGGATCAGGACT | |
| EV71 | F: 5′-CTGTGCGAATTAAGGACAG |
| R: 5′-GAGTTCCATAGGTGACAGC | |
| CVB3 | F: 5′-CGGTACCTTTGTGCGCCTGTT |
| R: 5′-GCGGTGCTCATCGACCTGA | |
| Gapdh | F: 5′-ATGTTTGTGATGGGTGTGAA |
| R: 5′-ATGCCAAAGTTGTCATGGAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, Y.; Li, C.; Liang, Y.; Jiang, Y.; Zhang, H.; Ouyang, J.; Zhang, W.; Deng, R.; Tan, Q.; Yu, X.; et al. Umbilical Cord Mesenchymal-Stem-Cell-Derived Exosomes Exhibit Anti-Oxidant and Antiviral Effects as Cell-Free Therapies. Viruses 2023, 15, 2094. https://doi.org/10.3390/v15102094
Meng Y, Li C, Liang Y, Jiang Y, Zhang H, Ouyang J, Zhang W, Deng R, Tan Q, Yu X, et al. Umbilical Cord Mesenchymal-Stem-Cell-Derived Exosomes Exhibit Anti-Oxidant and Antiviral Effects as Cell-Free Therapies. Viruses. 2023; 15(10):2094. https://doi.org/10.3390/v15102094
Chicago/Turabian StyleMeng, Yi, Chengcheng Li, Yicong Liang, Yu Jiang, Haonan Zhang, Jianhua Ouyang, Wen Zhang, Rumei Deng, Qiuping Tan, Xiaolan Yu, and et al. 2023. "Umbilical Cord Mesenchymal-Stem-Cell-Derived Exosomes Exhibit Anti-Oxidant and Antiviral Effects as Cell-Free Therapies" Viruses 15, no. 10: 2094. https://doi.org/10.3390/v15102094