Abstract
Several mutations in the surface (S), basal core promoter (BCP), and precore (PC) genes of the hepatitis B virus have been linked to inaccurate diagnosis and the development of immune escape mutants (IEMs) of the infection, which can lead to chronic infection. Understanding the prevalence and spread of these mutations is critical in the global effort to eliminate HBV. Blood samples were collected from 410 people in Osun and Ekiti states, southwest Nigeria, between 2019 and 2021. Participants were drawn from a group of asymptomatic people who were either blood donors, outpatients, or antenatal patients with no record of HBV infection at the medical outpatients’ unit of the hospital. DNA was extracted from plasma using a Qiagen DNEasy kit, followed by nested PCR targeting HBV S and BCP/PC genes. The Sanger sequencing method was used to sequence the positive PCR amplicons, which were further analyzed for IEMs, BCP, and PC mutations. HBV-DNA was detected in 12.4% (51/410) of individuals. After DNA amplification and purification, 47.1% (24) of the S gene and 76.5% (39) of the BCP/PC gene amplicons were successfully sequenced. Phylogenetic analysis showed that all the HBV sequences obtained in this study were classified as HBV genotype E. Mutational analysis of the major hydrophilic region (MHR) and a-determinant domain of S gene sequences revealed the presence of three immune escape mutations: two samples harbored a T116N substitution, six samples had heterogenous D144A/N/S/H substitution, and one sample had a G145E substitution, respectively. The BCP/PC region analysis revealed a preponderance of major BCP mutants, with the prevalence of BCP double substitutions ranging from 38.5% (A1762T) to 43.6% (G1764A). Previously reported classical PC mutant variants were observed in high proportion, including G1896A (33.3%) and G1899A (12.8%) mutations. This study confirms the strong presence of HBV genotype E in Nigeria, the ongoing circulation of HBV IEMs, and a high prevalence of BCP/PC mutants in the cohorts. This has implications for diagnosis and vaccine efficacy for efficient management and control of HBV in the country.
1. Introduction
The hepatitis B virus infection accounts for one-third of all liver cancer deaths globally, with hepatocellular carcinoma (HCC) being the leading cause of death due to cancer [1,2]. In low- and middle-income countries, hepatitis B-related HCC accounts for two-thirds of all liver cancer cases [3]. According to the World Health Organization (2019) estimate, more than 820,000 HBV-related deaths occur yearly due to liver cirrhosis and HCC, while 296 million individuals worldwide suffer from chronic hepatitis B. About 20 million Nigerians are estimated to be infected with HBV [4,5].
HBV is a partly double-stranded virus that has four overlapping open reading frames (ORFs) that code for the surface proteins (PreS1/PreS2/S): the polymerase (Pol), the capsid proteins (PreC/C), and the X protein [6]. The HBV S gene encoding the small HBsAg contains clusters of B-cell epitopes known as the major hydrophilic region (MHR), spanning amino acid position 99–169 [6]. Unlike other DNA viruses, HBV has a variable genome with ten genotypes and sub-genotypes based on 4–8% intra-genotypic divergence [7]. The HBV genotype has a particular geographic distribution worldwide, with genotypes A, D, and E being the most prevalent in Africa. The most prevalent HBV genotype in West Africa is E [8]. HBV has a higher mutation rate than other DNA viruses because it replicates via an intermediate RNA and a reverse transcriptase incapable of proofreading [9].
Despite chronically increasing HBV DNA levels and active liver illness, HBV strains from various nations have been shown to fail to produce HBeAg. This failure has been linked to mutations in the basal core promoter (BCP) and precore (PC) regions [10,11,12]. Specifically, it has been noted that G1896A and G1899A result in a translational stop codon that is predicted to affect HBeAg expression, ultimately leading to HBeAg loss and sometimes HBeAg-negative chronic HBV (CHB) infection [13,14,15]. According to reports, 1762T/1764A double BCP mutations have been suggested to be one of the risk factors for HCC [16,17,18,19]. Furthermore, these mutations may lead to long-term HBV therapy, making patients susceptible to nucleos(t)ide analog drug resistance gene evolution [20,21,22]. With an estimated incidence ranging between 2.5% and 40% over the entire country, Nigeria has long been one of the most HBV-endemic nations in sub-Saharan Africa [23,24]. Despite this, there is a severe shortage of reliable epidemiological data, little public awareness, and a critical need for accurate diagnosis and clinical care of HBV infection, most importantly among asymptomatic and apparently healthy individuals in Nigeria, particularly regarding IEMs and DRMs.
The majority of asymptomatic people are unaware carriers; this tends to increase the continuous spread and transmission within the community. Possible modes of transmission include but are not limited to blood transfusions, vertical transmission from mother to child during childbirth, injecting drug use, re-use of contaminated needles and syringes, and sexual contact [25,26]. The risk of asymptomatic infection with HBV is enormous and has long-term implications for disease elimination and eradication. Most importantly, infection with HBV undetected by the regular and most common testing assay in blood donors may be transmitted by blood transfusion. Therefore, active genomic surveillance of HBV infection is important to effectively control the disease.
This study provides insight into the possible emergence and spread of IEMs and BCP/PC mutants in asymptomatic HBV genotype E carriers in southwestern Nigeria.
2. Materials and Methods
2.1. Ethical Consideration and Clinical Sample Collection
This research was approved by the State Specialist Hospital Health Research and Ethics Committee, Osun State (HREC/27/04/2019/SSHO/502), Ekiti State University Teaching Hospital Ethical Research Committee (EKSUTH/A67/2019/08/003), and the Ladoke Akintola University of Technology (LAUTECH) Hospital Research Ethics Committee (LTH/2019/12/438). Before sample collection, all enrolled participants provided their written informed consent. Blood samples were obtained from 410 individuals in the southwestern Nigerian states of Osun and Ekiti between 2019 and 2021. At the time of blood collection, all the study participants were from a cohort of either blood donors, outpatients, or antenatal patients at the hospital’s medical outpatients’ units of the selected hospitals in Osun and Ekiti states and apparently asymptomatic for liver diseases. The serological profile is based on ELISA testing of this study’s cohorts’ HBsAg elsewhere [27]. All samples were tested for six HBV serological markers, including hepatitis B virus surface antigen (HBsAg), hepatitis B virus surface antibody (HBsAb), hepatitis B virus e antigen (HBeAg), antibody to hepatitis B virus E antigen (anti-HBe), hepatitis B core antigen to immunoglobulin M (HBcAb-IgM), and hepatitis B core total (antiHBc) using commercially available enzyme-linked immunosorbent assay (ELISA) kits (Melsin Medical Co., Changchun, China) following the manufacturer’s instruction.
2.2. HBV Genomic DNA Extraction
Total HBV-DNA were extracted from 200ul of serum from the four hundred and ten (410) patients’ sera using a Qiagen DNEasy kit (Lot number 163049045; Qiagen, Germany), according to the manufacturer’s instructions. DNA was eluted in 50 µL and stored at −80 °C until further use.
2.3. PCR Amplification of the HBV S-Gene
The nested PCR technique was used for the amplification of the fragments of the HBV S gene according to the method of Forbi and others (2010) and Umego and others (2022). The 400 bp fragment of the partial “S” gene region was amplified using the PuReTaqTM Ready-To-Go PCR Beads in strip tubes (Sigma-Aldrich® Darmstadt, Germany Lot # 17262518). The position of the codons in the S gene covered by the PCR ranges from 22 to 170. The PCR primers used were HBV_S1F (5”-CTAGGACCCCTGCTCGTGTT-3’) and HBV_S1R (5′-CGAACCACTGAACAAATGGCACT-3’) for the first round, and the second-round primers were HBV_SNF (5′-GTTGACAAGAATCCTCACAATACC-3’) and HBV_SNR (5′-GAGGCCCACTCCCATA-3). First- and second-round PCR reaction conditions were similar, except that extracted DNA was used as the template for the first-round PCR, while the first-round PCR products were used as a template for the second-round PCR amplification. The PCR amplification was carried out in a 20 µL reaction by adding 13 µL RNase-free water, 1µL of each of the primers (made in 20 µM concentrations), and 5 µL of a DNA template into the strip tube containing the puReTaq beads. A PCR procedure was carried out using an Eppendorf Thermal cycler (Eppendorf, Stevenage, UK) as follows: 94 °C for 3 min, followed by 45 cycles of denaturation, 94 °C for 30 s, annealing, 55 °C for 60 s and elongation, and 70 °C for 40 s with a ramp of 40% from 55 °C to 70 °C. The reaction was further elongated at 72 °C for 7 min and held at 4 °C until the reaction was terminated. PCR products were resolved on 2% agarose gel stained with ethidium bromide and viewed using a UV transilluminator.
2.4. PCR Amplification BCP/PC Genome Region
The BCP/PC region was amplified based on a previously reported nested PCR protocol targeting a 360bp fragment of the BCP/PC region, as described by [28]. Briefly, DNA amplification was performed using puReTaqTM Ready-To-Go PCR Beads in strips (Sigma-Aldrich® Lot # 17262518). The first round of PCR amplification was carried out using a 20 µL reaction, adding 16µL of RNase-free water, 1 µL of each of the primers (made in 20 µM concentrations), and 2 µL of a DNA template into the strip tube containing the puReTaq bead. The first round of PCR primers used are BCP_PC F1 (5-GCATGGAGACCACCGTGAAC-3) and BCP_PC R1 (5GGAAAGAAGTCCGAGGGCAA-3). The thermal cycling conditions were an initial denaturation of 30 s at 94 °C, followed by 55 °C at 60 s, 35 cycles of 60 s at 72 °C denaturation, 2 min at 72 °C annealing, 30 s at 68 °C extension, and a final extension for 10 min at 68 °C. The second round of PCR amplification was carried out using a 20 µL reaction, adding 16 µL of RNase-free water, 1 µL of each of the primers (made in 20 µM concentrations), and 2µL of DNA template from the first round of PCR. The second round of PCR primers used are BCP_PC F2 (5- CATAAGAGGACTCTTGGACT-3 and BCP_PC R2 (5- GGCAAAAAACAGAGTAACTC-3). The cycling conditions were an initial denaturing of 94 °C for 3 min, followed by 30 cycles (denaturation, 94 °C for 30 s, annealing, 55 °C for 30 s and elongation, 72 °C for 60 s with a ramp of 40% from 55 °C to 70 °C). The reaction was further elongated at 72 °C for 2 min and held at 4 °C until the reaction was terminated. The PCR procedure used an Eppendorf Thermal cycler (Eppendorf, UK). The PCR products were resolved in 2% agarose gel stained with ethidium bromide and viewed using a UV transilluminator.
2.5. Sequencing of the S and BCP/PC Genes Using the Sanger Sequencing Method
Five microliters of the secondary amplicons from the HBS gene and BCP/PC gene with the expected band size (400 bp and 360 bp, respectively, codon positions 22–170 and 54–271, respectively) were purified with 2 uL of ExoSAP-IT (Affymetrix, Santa Clara, CA, USA) at 37 °C for 15 min and inactivated at 80 °C for 15 min. For sequencing prep, 4 uL of big dye terminator 3.1 ready reaction mix (Life Technologies, Carlsbad, CA, USA), 7 uL of nuclease-free water, 2 uL each of secondary PCR (HBV_SNF and HBV_SNR), and (BCP_PCF1 and BCP_PCF2) primers were added to the purified secondary amplicons. PCR amplification was performed on a thermocycler (Eppendorf Vapo. Protect Mastercycler pro, Germany) in a final volume of 20 μL at cycling conditions of 96 °C for 60 s, 30 cycles 96 °C for 10 s, 50 °C for 5 s, and 70 °C for 4 min. The sequencing products were further purified using the bigdye Xterminator kit (Life Technologies, Carlsbad, CA, USA) (90 uL of SAM solution and bigdye Xterminator bead solution per sample) vortexed for 20 min at room temperature. Sanger sequencing was performed on the Applied Biosystems 3500 XL series Genetic Analyzer at the African Centre of Excellence for Genomics of Infectious Diseases, Redeemer’s University, Nigeria.
2.6. Phylogenetic and Gene Variability Analysis of HBV Partial S Gene and BCP/PC Gene Sequences
The forward and reverse sequences per sample were stitched into contigs for the HBV S and BCP/PC genes. Each contig was subjected to a Blastn search on the National Centre for Biotechnology Information (NCBI) page. Aligning nucleotide sequences and deduced amino acids with those of reference HBV genotypes A-H from the NCBI virus was performed using the MAFFT online service [29]. Phylogenetic trees were constructed using IQ-TREE version 1.6.12 [30] with ModelFinder [31] and ultrafast bootstrap (1000 replicates) [32]. The tree was visualized using Interactive Tree of Life (iTOL) v5 [33].
For the S gene ORF, we examined both the major hydrophilic region (MHR) (amino acid position 99-169) and the a-determinant domain (found within the MHR; amino acid position 124-147) using BioEdit and geno2pheno software (https://hbv.geno2pheno.org/ accessed on 25 March 2023). All our sequences were aligned with the HBV genotype E reference sequence (accession number LC513651) from the NCBI.
During BCP/PC sequences analysis, we examined nucleotide changes at the TA-rich genome regions, including nt 1750-1755, 1758-1762, 1771-1775, and 1778-1795. Kozak sequence mutants (translational genes-nt1809-1812), PC initiation (1814-1816), the post-translational mutant gene (G1862T), and the translational stop codon (G1896A with C1858T) were used [28,34]. The partial basal core promoter (BCP) and precore (PC) region gene sequences obtained from this study were aligned with the previously reported wild type (X75657.1) and mutant (AF28996.1).
2.7. Statistical Analysis
A chi-squared test was performed on categorical data to test for significant correlations between the different factors using SPSS version 21 (IBM Corp. Released 2012. IBM SPSS Statistics for Windows, Version 21.0. Armonk, NY, USA). A less than 0.05 p-value was used to determine statistical significance.
3. Results
3.1. Socio-Demographic Characteristics of the Respondents
In this study, 150 samples were from Ekiti and 260 were from Osun. The mean age of the 410 participants included in the study was 36.77 ± 11.26, and 36.8% of the respondents were between 25 and 34 years of age. A total of 283 (69%) respondents were female, while 127 (21%) were male. A total of 308 (75.1%) of the 410 participants were married. A total of 100 (24.4%) are single, and 2 (0.5%) are divorced. Those who had completed graduate studies were 184 (44.9%), while 221 (53.9%) of the respondents were self-employed (Table 1).

Table 1.
Socio-demographic characteristics of the total respondents.
3.2. HB S and BCP/PC Gene-Specific PCR Amplification Results
The 410 samples were initially screened by ELISA, of which 51 (12.4%) were positive to HBsAg and 12 (6.6%) were positive to HBeAg. The results of the serological profile are published elsewhere [27]. HBV (HB gene- and/or BCP/CP gene-positive PCR) was detected in 12.4% (n = 51/410) of individuals who were also HbsAg-positive by ELISA. The mean age of the 51 samples is 36.08 ±8.03. Of the 51 HBV-PCR-positive samples, 60.8% (n = 31) were from Osun, while 39.2% (n = 20) were from Ekiti. Of the 51 HBV DNA-positive samples, 32 (62.7%) and 19 (37.3%) were detected in females and males, respectively. According to the categories of individuals, 43.1% (n = 22) were from blood donors, 41.2% (n = 21) were from outpatients, and 15.7% (n = 8) were from pregnant women. Seven (13.7%) were positive to HbeAg, while forty-four (86.3%) were negative to HBeAg, respectively (Table 2, Supplementary Table S2).
Table 2.
Demographic profile of participants with HBV-positive PCR outcomes analyzed in this study.
3.3. Phylogenetic Analysis of the HBV S and BCP/PC Genes
PCR amplification of the S gene was successful in 24 (47%) of the 51 isolates that tested positive for HBsAg. Results from both geno2pheno and phylogenetic analysis using IQTREE showed that all the 24 HBV S gene sequences obtained in this study were classified as HBV genotype E and clustered around samples from Nigeria, Guinea, and the Central African Republic (Figure 1).
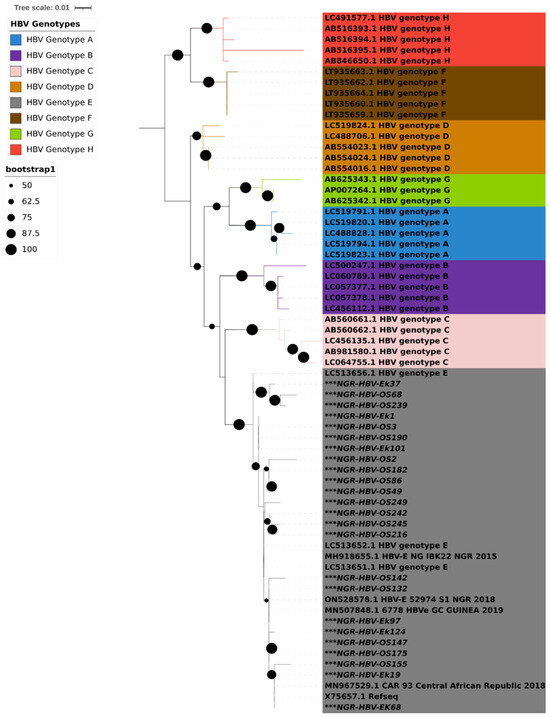
Figure 1.
Maximum likelihood tree with ModelFinder based on partial S gene sequences with 1000 bootstrap replications. All HBV genotypes are color coded, as shown in the legend. HBV sequences reported in this study are asterisked (***) in black. The Interactive Tree of Life (iTOL) v5 with midpoint rooting was used to visualize the tree.
3.4. Analysis of Immune Escape Mutations Present in the HBV Samples
Of the 410 samples screened by PCR, 51 (12.4%) were positive for HB gene amplification and were sequenced. Only 47.1% (n = 24) of the S gene amplicon sequences were of good quality for downstream analysis. Mutational analysis of the MHR and a-determinant domain of S gene sequences in this study revealed the presence of three immune escape mutations in 37.5% (9/24) of the samples: two samples (8.3%) (NGR-HBV-OS239 and NGR-HBV-OS68) had a T116N substitution, six samples (25%) (NGR-HBV-OS239, NGR-HBV-OS68, NGR-HBV-OS155, NGR-HBV-OS242, NGR-HBV-OS216, and NGR-HBV-OS245) had heterogenous D144A/N/S/H substitution, and one sample (4.2%) (NGR-HBV-OS155) had a G145E substitution (Figure 2).
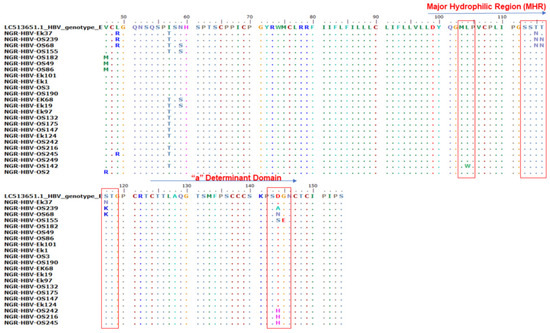
Figure 2.
Alignment of the partial S gene sequences with the HBV genotype E reference sequence (accession number LC513651). The various mutations at the major hydrophilic region (amino acid position 99–169) and the a-determinant domain found within the MHR (amino acid position 124–147).
3.5. Analysis of the Basal Core Protein/Precore (BCP/PC) Genome Variability in the HBV Samples
Of the 51 BCP/PC PCR-positive samples sequenced, 39 (76.5%) sequences were of good quality for mutational analysis. The mutational analysis of the BCP/PC region revealed various substitutions in the TA-rich genome regions. Specifically, we observed a T1753G/C in two samples (NGR-HBV-OS245 and NGR-HBV-EK37), while samples NGR-HBV-OS2 and NGR-HBV-EK73 both had T1754A and A1755G substitutions, respectively (Figure 3A). We equally observed a preponderance of major BCP mutants with the prevalence of BCP double substitutions ranging from 38.5% (A1762T) to 43.6% (G1764A). Interestingly, the BCP double mutants were highest among the outpatient group, as well as among individuals with HBeAg-negative status (33.3% and 38.5%) (Table 3). Analysis of the presence of Kozak sequence mutants (nt 1809-1812) showed that two samples (NGR-HBV-EK105 and NGR-HBV-OS80) had a G1809C/A substitution, while one sample (NGR-HBV-OS49) had an A1811C substitution (Figure 3B). Analysis of the PC initiation region (nt1814-1816) revealed one sample (NGR-HBV-EK105) having an A1814C substitution. No post-translational mutant gene (G1862T) was detected in our study (Figure 4). Previously reported classical PC mutant variants were observed in high proportion, including G1896A (33.3%) and G1899A (12.8%), and were equally higher among individuals with HbeAg-negative status (20.5% and 7.7%) (Figure 4).
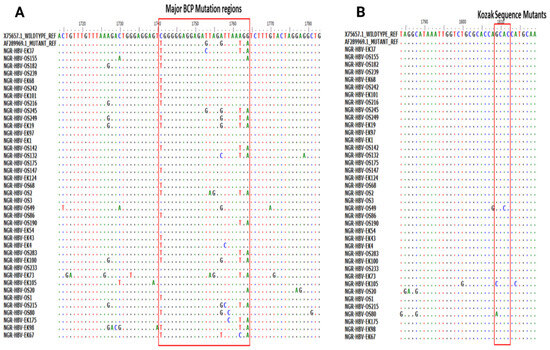
Figure 3.
(A) Partial Basal core promoter (BCP) region gene sequences obtained from this study and aligned with previously reported wildtype (X75657.1) and mutant (AF28996.1) Alignment showed the various mutations at the BCP region with empahsis toon the G1896/G1899A vaiant and (B) various mutations at the Kozak sequence region (nt 1809-1812).
Table 3.
Frequency distribution of BCP and PC mutation among HBeAg status and study population.
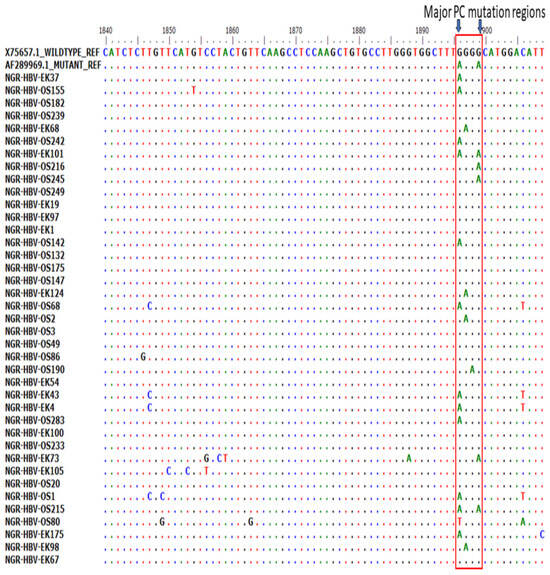
Figure 4.
Precore (PC) region gene sequences obtained from this study aligned with previously reported wild type (X75657.1) and mutant (AF28996.1). Results showed the presence of various mutations at the PC region with emphasis on G1896/G1899A variants.
The HBV sequences generated in this study were deposited into the NCBI GenBank under accession numbers OR001858, OR001861, OR001862, OR001859, OR001869, OR001876, OR001865, OR001877, OR001878, OR001879, OR001870, OR001871, OR001866, OR001880, OR001872, OR001873, OR001874, OR001875, OR001860, OR001881, OR001867, OR001863, OR001864, and OR001868.
4. Discussion
The prevalence of HBV infection in Nigeria is between 10% and 15%. Thus, it is a major public health concern, despite the availability of the HBV vaccine for adults and children [35]. Vaccine- and diagnostic-escape phenomena have reportedly been caused by immune escape mutations in general [36]. This study documents the prevalence of immune escape mutants (IEMs) and BCP/PC HBV mutants circulating among southwestern Nigeria’s HBV genotype E-infected cohort without symptoms. All the HBV isolates analyzed in this study were all genotype E of the virus. This is not surprising, as this is the most common virus genotype reported in the country [37]. Other genotypes reported in Nigeria are A, B, C, and D [36,38,39]. Specifically, we identified important substitutions in the “a”-determinant domain and clinically relevant mutations in the BCP and PC HBV genomic regions in asymptomatic hospital attendees. The results of this study showed the presence of three IEMs: T116N, D144A/N/S/H, and G145E, which have been linked to decreased antibody binding to wild type S protein by virions and subviral particles, resulting in breakthrough infections and diagnostic failure [40,41,42,43].
The prevalence of HBV IEMs in the restricted, immunodominant “a”-determinant region (ADR) between amino acid positions 124–147 was 37.5% (n = 9/24), where nine connotes the number of samples with IEM mutations occurring in the antigenic determinant region. The prevalence of HBV IEMs in this study was slightly higher than what was earlier reported by Osasana in Nigeria and Lazarevec in Brazil, where a prevalence of 29% and 10.7% were reported, respectively [39,44]. This high prevalence may be due to the fact that the study participants were asymptomatic and, therefore, have yet to undergo HBV infection treatment. This study further corroborates the circulation of IEMs in Nigeria. However, the pattern of IEMs documented in this study differs from previously documented dominant Q129H [36] and G145K [23,45] IEMs in Nigeria. Other notable non-immune escape mutations detected in the MHR domain were L104W (1), T115N (3), and S117N/K (3).
Interestingly, all the IEMs were detected in HBV sequences from outpatients and apparently healthy prospective blood donors in Osun state. Previously detected IEMs in Nigeria were documented in different populations, including apparently healthy community dwellers, pregnant women, and prospective blood donors in Ekiti, Ondo, and Oyo [36,44]. The spread of these IEMs in Nigeria may jeopardize HBV vaccine efficacy and have public health implications in managing and controlling the infection if not addressed. The varying patterns of IEMs imply the continuous evolution of IEMs in the region; thus, there is a need for more robust HBV surveillance in different parts of the country to ascertain their true prevalence and identify factors that may enhance their transmission.
Mutations within and outside the “a”-determinant domain have been linked to Occult HBV Infection (OBI) [46,47,48]. Substitutions that result in hydrophobicity, the presence of a phenyl group, and charges (previously absent) in the side chain of amino acid residues located in the MHR may enhance immune escape strain adaptation and circulation via decreased HBsAg secretion and impaired reactivity with anti-HBs antibodies among the asymptomatic study cohort [47].
A high proportion of classical BCP/PC mutant variants identified in the current study had been previously associated with HBeAg-negative CHB infection and the down-regulation of PC mRNA transcription [49]. The classical BCP double mutations A1762T/G1764A (38.5%/43.6%) were the most common and were equally highest among the outpatient group (23.1%/28.2%) and HBeAg-negative participants (33.3%/38.5%), respectively (Table 3). The classical BCP double mutations A1762T/G1764A were the most common (Table 3). Additionally, the G to A point mutation at nucleotide 1896 of the precore region results in the conversion of tryptophan (TGG) to a stop codon (TAG), which is known to abolish HBeAg synthesis [13,50,51], was found to be 33.3% in the HBV genotype E-infected cohort in this study. The frequency of BCP/PC mutations found in this study is similar to the previously reported prevalence found in the ART-naïve HIV/HBV co-infected genotype E-infected cohort in Nigerians and calls for robust HBV surveillance in the country [52]. Regardless of the study group, the majority of classical BCP/PC mutants, such as BCP double mutant sequences, PC initiation, and G1896A with G1899A mutations, were significantly more common in HbeAg-negative subjects. The limitation of this study is that we could not explain the observed high proportion and implications of classical BCP/PC mutant variants identified across the study subjects because participants’ liver disease status and HBV viral load were not assessed due to financial constraints.
However, the continuous spread of the IEMs and BCP/PC mutations among asymptomatic patients may contribute to vaccine and diagnostic failure [53,54] and the evolution of new strains. Molecular surveillance of circulating HBV must be conducted within the country to guide the implementation of effective management and control methods.
Some implications of these mutations observed among asymptomatic patients are the ability to easily transmit the virus within the community, as most patients are unaware carriers of HBsAg, and the mutations. These findings from our study also emphasize the risk of hepatitis B virus transmission through blood donation or transfusion. These findings also been implicated in cryptogenic liver diseases, progressive acute and chronic HBV infection, and hepatocellular carcinoma development. It is, therefore, imperative to conduct more in-depth epidemiological studies on the success or failure of HBV vaccination programs in Nigeria and Africa and ensure all vaccinated individuals complete the expected dose of vaccine.
In conclusion, a high frequency of IEM, BCP, and PC HBV mutants were found in asymptomatic outpatients from southwestern Nigeria’s HBV genotype E-infected cohort. This suggests the increased likelihood of potential HBeAg-negative chronic HBV infection and the emergence of HBV strains, which may result in breakthrough infections and diagnostic failure in this setting. The IEM pattern observed in this study differed from previously documented dominant IEMs in Nigeria, implying the continuous evolution of IEM strains in the country. More research is needed to determine their prevalence and identify factors contributing to their spread. There is a need for active serological and molecular surveillance of the circulating HBV and mutations on its transmission, spread, and effect on asymptomatic carriers, as well as its interactions with the existing vaccine.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/v15112188/s1, Table S1: Prevalence of HBV biomarkers by socio-demographic characteristics of participants; Table S2: Demographic data and molecular results of HBsAg-positive cohorts analyzed in this study.
Author Contributions
Conceptualization, O.A.S., O.F. and I.K.; data curation, O.A.S.; formal analysis, O.A.S.; investigation, O.A.S., J.U.O., B.A. and P.E.; methodology, O.A.S., J.U.O., B.A. and P.E.; resources: C.H., O.F. and I.K.; supervision, C.H., O.F. and I.K.; validation, O.A.S. and J.U.O.; writing—original draft, O.A.S.; writing—review and editing, J.U.O., P.E., O.F. and I.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from NIH-H3Africa (https://h3africa.org), accessed on 1 June 2021 (U01HG007480 and U54HG007480); the World Bank Grant (worldbank.org) accessed on 1 June 2021 (ACE IMPACT-ACE 019), and the sentinel/Audacious Project.
Institutional Review Board Statement
This research was approved by the State Specialist Hospital Health Research and Ethics Committee, Osun State (HREC/27/04/2019/SSHO/502), Ekiti State University Teaching Hospital Ethical Research Committee (EKSUTH/A67/2019/08/003), and the Ladoke Akintola University of Technology (LAUTECH) Hospital Research Ethics Committee (LTH/2019/12/438).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Genome sequences of HBV reported in this study have been deposited in GenBank under accession numbers OR001858–OR001881.
Acknowledgments
We would like to thank the medical staff of the Medical Outpatient Clinic at the LAUTECH Teaching Hospital, Osogbo, Osun State, and Ekiti State University Teaching Hospital, Ekiti State, during the collection of the samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Global Burden of Disease Liver Cancer Collaboration; Akinyemiju, T.; Abera, S.; Ahmed, M.; Alam, N.; Alemayohu, M.A.; Allen, C.; Al-Raddadi, R.; Alvis-Guzman, N.; Amoako, Y.; et al. The Burden of Primary Liver Cancer and Underlying Etiologies from 1990 to 2015 at the Global, Regional, and National Level: Results from the Global Burden of Disease Study 2015. JAMA Oncol. 2017, 3, 1683–1691. [Google Scholar] [CrossRef]
- MacLachlan, J.H.; Cowie, B.C. Liver Cancer Is the Fastest Increasing Cause of Cancer Death in Australians. Med. J. Aust. 2012, 197, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-C.; Huang, D.Q.; Nguyen, M.H. Global Burden of Hepatitis B Virus: Current Status, Missed Opportunities and a Call for Action. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 524–537. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Consolidated Guidelines on HIV Prevention, Testing, Treatment, Service Delivery and Monitoring: Recommendations for a Public Health Approach; World Health Organization: Geneva, Switzerland, 2021; ISBN 9789240031593. [Google Scholar]
- Ajuwon, B.I.; Yujuico, I.; Roper, K.; Richardson, A.; Sheel, M.; Lidbury, B.A. Hepatitis B Virus Infection in Nigeria: A Systematic Review and Meta-Analysis of Data Published between 2010 and 2019. BMC Infect. Dis. 2021, 21, 1120. [Google Scholar] [CrossRef] [PubMed]
- Caligiuri, P.; Cerruti, R.; Icardi, G.; Bruzzone, B. Overview of Hepatitis B Virus Mutations and Their Implications in the Management of Infection. World J. Gastroenterol. 2016, 22, 145–154. [Google Scholar] [CrossRef]
- Kurbanov, F.; Tanaka, Y.; Mizokami, M. Geographical and Genetic Diversity of the Human Hepatitis B Virus. Hepatol. Res. 2010, 40, 14–30. [Google Scholar] [CrossRef]
- Kafeero, H.M.; Ndagire, D.; Ocama, P.; Kato, C.D.; Wampande, E.; Walusansa, A.; Kajumbula, H.; Kateete, D.; Ssenku, J.E.; Sendagire, H. Mapping Hepatitis B Virus Genotypes on the African Continent from 1997 to 2021: A Systematic Review with Meta-Analysis. Sci. Rep. 2023, 13, 5723. [Google Scholar] [CrossRef]
- Harrison, A.; Lemey, P.; Hurles, M.; Moyes, C.; Horn, S.; Pryor, J.; Malani, J.; Supuri, M.; Masta, A.; Teriboriki, B.; et al. Genomic Analysis of Hepatitis B Virus Reveals Antigen State and Genotype as Sources of Evolutionary Rate Variation. Viruses 2011, 3, 83–101. [Google Scholar] [CrossRef]
- Chalid, M.T.; Turyadi; Ie, S.I.; Sjahril, R.; Wahyuni, R.; Nasrum Massi, M.; Muljono, D.H. A Cautionary Note to Hepatitis B E Antigen (HBeAg)-Negative Test Results in Pregnant Women in an Area Prevalent of HBeAg-Negative Chronic Hepatitis B. J. Med. Virol. 2023, 95, e28125. [Google Scholar] [CrossRef]
- Rodriguez-Frias, F.; Jardi, R.; Buti, M.; Schaper, M.; Hermosilla, E.; Valdes, A.; Allende, H.; Martell, M.; Esteban, R.; Guardia, J. Hepatitis B Virus Genotypes and G1896A Precore Mutation in 486 Spanish Patients with Acute and Chronic HBV Infection. J. Viral Hepat. 2006, 13, 343–350. [Google Scholar] [CrossRef]
- Papatheodoridis, G.V.; Hadziyannis, S.J. Diagnosis and Management of Pre-Core Mutant Chronic Hepatitis B. J. Viral Hepat. 2001, 8, 311–321. [Google Scholar] [CrossRef]
- Laras, A.; Koskinas, J.; Avgidis, K.; Hadziyannis, S.J. Incidence and Clinical Significance of Hepatitis B Virus Precore Gene Translation Initiation Mutations in E Antigen-Negative Patients. J. Viral Hepat. 1998, 5, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulou, A.; Karayiannis, P. HBeAg Negative Variants and Their Role in the Natural History of Chronic Hepatitis B Virus Infection. World J. Gastroenterol. 2014, 20, 7644–7652. [Google Scholar] [CrossRef]
- Chen, C.-H.; Lee, C.-M.; Hung, C.-H.; Hu, T.-H.; Wang, J.-H.; Wang, J.-C.; Lu, S.-N.; Changchien, C.-S. Clinical Significance and Evolution of Core Promoter and Precore Mutations in HBeAg-Positive Patients with HBV Genotype B and C: A Longitudinal Study. Liver Int. 2007, 27, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.-H.; Chen, P.-J.; Lai, M.-Y.; Chen, D.-S. Basal Core Promoter Mutations of Hepatitis B Virus Increase the Risk of Hepatocellular Carcinoma in Hepatitis B Carriers. Gastroenterology 2003, 124, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, S.-A.; Do, S.Y.; Kim, B.-J. Precore/core Region Mutations of Hepatitis B Virus Related to Clinical Severity. World J. Gastroenterol. 2016, 22, 4287–4296. [Google Scholar] [CrossRef] [PubMed]
- Baptista, M.; Kramvis, A.; Kew, M.C. High Prevalence of 1762(T) 1764(A) Mutations in the Basic Core Promoter of Hepatitis B Virus Isolated from Black Africans with Hepatocellular Carcinoma Compared with Asymptomatic Carriers. Hepatology 1999, 29, 946–953. [Google Scholar] [CrossRef]
- Yang, Z.; Zhuang, L.; Lu, Y.; Xu, Q.; Tang, B.; Chen, X. Naturally Occurring Basal Core Promoter A1762T/G1764A Dual Mutations Increase the Risk of HBV-Related Hepatocellular Carcinoma: A Meta-Analysis. Oncotarget 2016, 7, 12525–12536. [Google Scholar] [CrossRef]
- Chen, C.-H.; Lee, C.-M.; Lu, S.-N.; Changchien, C.-S.; Wang, J.-C.; Wang, J.-H.; Hung, C.-H.; Hu, T.-H. Comparison of Sequence Changes of Precore and Core Promoter Regions in HBeAg-Positive Chronic Hepatitis B Patients with and without HBeAg Clearance in Lamivudine Therapy. J. Hepatol. 2006, 44, 76–82. [Google Scholar] [CrossRef]
- Shin, J.W.; Chung, Y.-H.; Choi, M.H.; Kim, J.A.; Ryu, S.H.; Jang, M.K.; Kim, I.S.; Park, N.H.; Lee, H.C.; Lee, Y.S.; et al. Precore Stop Codon Mutation of Hepatitis B Virus Is Associated with Low Breakthrough Rate Following Long-Term Lamivudine Therapy. J. Gastroenterol. Hepatol. 2005, 20, 844–849. [Google Scholar] [CrossRef]
- Kuo, L.-F.; Lee, C.-M.; Hung, C.-H.; Wang, J.-H.; Hu, T.-H.; Lu, S.-N.; Changchien, C.-S.; Chen, C.-H. High Risk of Hepatitis B Virus Reactivation in Nucleos(t)ide Analogue-Induced Hepatitis B E Antigen Seroconverters Older than 40 Years. Dig. Dis. Sci. 2014, 59, 2580–2587. [Google Scholar] [CrossRef]
- Faleye, T.O.C.; Adewumi, O.M.; Ifeorah, I.M.; Akere, A.; Bakarey, A.S.; Omoruyi, E.C.; Oketunde, K.; Awonusi, O.B.; Ajayi, M.R.; Adeniji, J.A. Detection and Circulation of Hepatitis B Virus Immune Escape Mutants among Asymptomatic Community Dwellers in Ibadan, Southwestern Nigeria. Int. J. Infect. Dis. 2015, 39, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Olayinka, A.T.; Oyemakinde, A.; Balogun, M.S.; Ajudua, A.; Nguku, P.; Aderinola, M.; Egwuenu-Oladejo, A.; Ajisegiri, S.W.; Sha’aibu, S.; Musa, B.O.P.; et al. Seroprevalence of Hepatitis B Infection in Nigeria: A National Survey. Am. J. Trop. Med. Hyg. 2016, 95, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Sabeena, S.; Ravishankar, N. Horizontal Modes of Transmission of Hepatitis B Virus (HBV): A Systematic Review and Meta-Analysis. Iran. J. Public Health 2022, 51, 2181–2193. [Google Scholar] [CrossRef]
- Screening and Testing Recommendations for Chronic Hepatitis B Virus Infection (HBV). Available online: https://www.cdc.gov/hepatitis/hbv/testingchronic.htm (accessed on 25 September 2023).
- Sobajo, O.A.; George, U.E.; Osasona, O.G.; Eromon, P.; Aborisade, O.Y.; Ajayi, O.D.; Folarin, O.A.; Komolafe, I.O.O. Seroprevalence, Co-Infection and Risk of Transmission of Hepatitis B and D Virus among Hospital Attendees in Two South-Western States in Nigeria. J. Immunoass. Immunochem. 2023, 44, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Belyhun, Y.; Liebert, U.G.; Maier, M. Analysis of HBV Basal Core Promoter/precore Gene Variability in Patients with HBV Drug Resistance and HIV Co-Infection in Northwest Ethiopia. PLoS ONE 2018, 13, e0191970. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An Online Tool for Phylogenetic Tree Display and Annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, R.; Kazim, S.N.; Bhattacharjee, J.; Sakhuja, P.; Sarin, S.K. Basal Core Promoter, Precore Region Mutations of HBV and Their Association with E Antigen, Genotype, and Severity of Liver Disease in Patients with Chronic Hepatitis B in India. J. Med. Virol. 2006, 78, 1047–1054. [Google Scholar] [CrossRef] [PubMed]
- Ogoina, D.; Pondei, K.; Adetunji, B.; Chima, G.; Isichei, C.; Gidado, S. Prevalence of Hepatitis B Vaccination among Health Care Workers in Nigeria in 2011-12. Int. J. Occup. Environ. Med. 2014, 5, 51–56. [Google Scholar]
- Adesina, O.A.; Akanbi, O.A.; Opaleye, O.O.; Japhet, M.O.; Wang, B.; Oluyege, A.O.; Klink, P.; Bock, C.-T. Detection of Q129H Immune Escape Mutation in Apparently Healthy Hepatitis B Virus Carriers in Southwestern Nigeria. Viruses 2021, 13, 1273. [Google Scholar] [CrossRef]
- Oladeinde, B.H.; Ekejindu, I.M.; Omoregie, R.; Odia, I.; Aguh, O.D.; Okwu, U.M. New Strains of Hepatitis B Virus Genotype E Circulating in Nigeria. Int. J. Health Sci. 2018, 12, 25–29. [Google Scholar]
- Ahmad, A.E.; Bakari, A.G.; Musa, B.O.P.; Mustapha, S.K.; Jamoh, B.Y.; Abdullahi, I.N.; Tahir, M.I.; Olatunji, A.O.; Maishanu, S.H.; Suleiman, A.B.; et al. Pattern of Prevalent Hepatitis B Virus Genotypes in Zaria, Nigeria. Niger. Postgrad. Med. J. 2019, 26, 80–86. [Google Scholar] [CrossRef]
- Osasona, O.G.; Oguntoye, O.O.; Arowosaye, A.O.; Abdulkareem, L.O.; Adewumi, M.O.; Happi, C.; Folarin, O. Patterns of hepatitis b virus immune escape and pol/rt mutations across clinical cohorts of patients with genotypes a, e and occult hepatitis b infection in Nigeria: A multi-centre study. Virulence 2023, 14, 2218076. [Google Scholar] [CrossRef]
- Cooreman, M.P.; Leroux-Roels, G.; Paulij, W.P. Vaccine- and Hepatitis B Immune Globulin-Induced Escape Mutations of Hepatitis B Virus Surface Antigen. J. Biomed. Sci. 2001, 8, 237–247. [Google Scholar] [CrossRef]
- Komatsu, H.; Inui, A.; Umetsu, S.; Tsunoda, T.; Sogo, T.; Konishi, Y.; Fujisawa, T. Evaluation of the G145R Mutant of the Hepatitis B Virus as a Minor Strain in Mother-to-Child Transmission. PLoS ONE 2016, 11, e0165674. [Google Scholar] [CrossRef] [PubMed]
- Zanetti, A.R.; Tanzi, E.; Manzillo, G.; Maio, G.; Sbreglia, C.; Caporaso, N.; Thomas, H.; Zuckerman, A.J. Hepatitis B Variant in Europe. Lancet 1988, 2, 1132–1133. [Google Scholar] [CrossRef]
- Lazarevic, I. Clinical Implications of Hepatitis B Virus Mutations: Recent Advances. World J. Gastroenterol. 2014, 20, 7653–7664. [Google Scholar] [CrossRef]
- Araujo, N.M.; Teles, S.A.; Spitz, N. Comprehensive Analysis of Clinically Significant Hepatitis B Virus Mutations in Relation to Genotype, Subgenotype and Geographic Region. Front. Microbiol. 2020, 11, 616023. [Google Scholar] [CrossRef]
- Faleye, T.O.C.; Adewumi, M.O.; Ifeorah, I.M.; Omoruyi, E.C.; Bakarey, S.A.; Akere, A.; Awokunle, F.; Ajibola, H.O.; Makanjuola, D.O.; Adeniji, J.A. Detection of Hepatitis B Virus Isolates with Mutations Associated with Immune Escape Mutants among Pregnant Women in Ibadan, Southwestern Nigeria. Springerplus 2015, 4, 43. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Svicher, V.; Cento, V.; Bernassola, M.; Neumann-Fraune, M.; Van Hemert, F.; Chen, M.; Salpini, R.; Liu, C.; Longo, R.; Visca, M.; et al. Novel HBsAg Markers Tightly Correlate with Occult HBV Infection and Strongly Affect HBsAg Detection. Antivir. Res. 2012, 93, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-H.; Yuan, Q.; Chen, P.-J.; Zhang, Y.-L.; Chen, C.-R.; Zheng, Q.-B.; Yeh, S.-H.; Yu, H.; Xue, Y.; Chen, Y.-X.; et al. Influence of Mutations in Hepatitis B Virus Surface Protein on Viral Antigenicity and Phenotype in Occult HBV Strains from Blood Donors. J. Hepatol. 2012, 57, 720–729. [Google Scholar] [CrossRef]
- Elkady, A.; Iijima, S.; Aboulfotuh, S.; Mostafa Ali, E.; Sayed, D.; Abdel-Aziz, N.M.; Ali, A.M.; Murakami, S.; Isogawa, M.; Tanaka, Y. Characteristics of Escape Mutations from Occult Hepatitis B Virus Infected Patients with Hematological Malignancies in South Egypt. World J. Hepatol. 2017, 9, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Buckwold, V.E.; Hon, M.W.; Ou, J.H. Mechanism of Suppression of Hepatitis B Virus Precore RNA Transcription by a Frequent Double Mutation. J. Virol. 1999, 73, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Jammeh, S.; Tavner, F.; Watson, R.; Thomas, H.C.; Karayiannis, P. Effect of Basal Core Promoter and Pre-Core Mutations on Hepatitis B Virus Replication. J. Gen. Virol. 2008, 89, 901–909. [Google Scholar] [CrossRef]
- Carman, W.F.; Jacyna, M.R.; Hadziyannis, S.; Karayiannis, P.; McGarvey, M.J.; Makris, A.; Thomas, H.C. Mutation Preventing Formation of Hepatitis B E Antigen in Patients with Chronic Hepatitis B Infection. Lancet 1989, 2, 588–591. [Google Scholar] [CrossRef]
- Grant, J.; Agbaji, O.; Kramvis, A.; Yousif, M.; Auwal, M.A.; Penugonda, S.; Ugoagwu, P.; Murphy, R.; Hawkins, C. Hepatitis B Virus Sequencing and Liver Fibrosis Evaluation in HIV/HBV Co-Infected Nigerians. Trop. Med. Int. Health 2017, 22, 744–754. [Google Scholar] [CrossRef]
- Khodadad, N.; Seyedian, S.S.; Moattari, A.; Biparva Haghighi, S.; Pirmoradi, R.; Abbasi, S.; Makvandi, M. In silico functional and structural characterization of hepatitis B virus PreS/S-gene in Iranian patients infected with chronic hepatitis B virus genotype D. Heliyon 2020, 6, e04332. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, M.; Faleo, G.; Leitner, T.; Zheng, W.; Zhang, Y.; Hassan, A.; Alwazzeh, M.J.; Fiore, J.R.; Ismail, M.; Santantonio, T.A. Molecular and Genetic Characterization of Hepatitis B Virus (HBV) among Saudi Chronically HBV-Infected Individuals. Viruses 2023, 15, 458. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).