
The Circulating miRNA Profile of Chronic Hepatitis D and B Patients Is Comparable but Differs from That of Individuals with HBeAg-Negative HBV Infection

 , , , ,
, , , ,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Assays
2.3. Isolation of Circulating HBsAg Particles
2.4. miRNA NGS Reads Bioinformatical Analysis
2.5. Read Mapping and Quantification of Gene Expression
2.6. MiR-B-Index
3. Results
3.1. NGS and Quality Controls
3.2. miRNA Profiling of HBsAg Particles
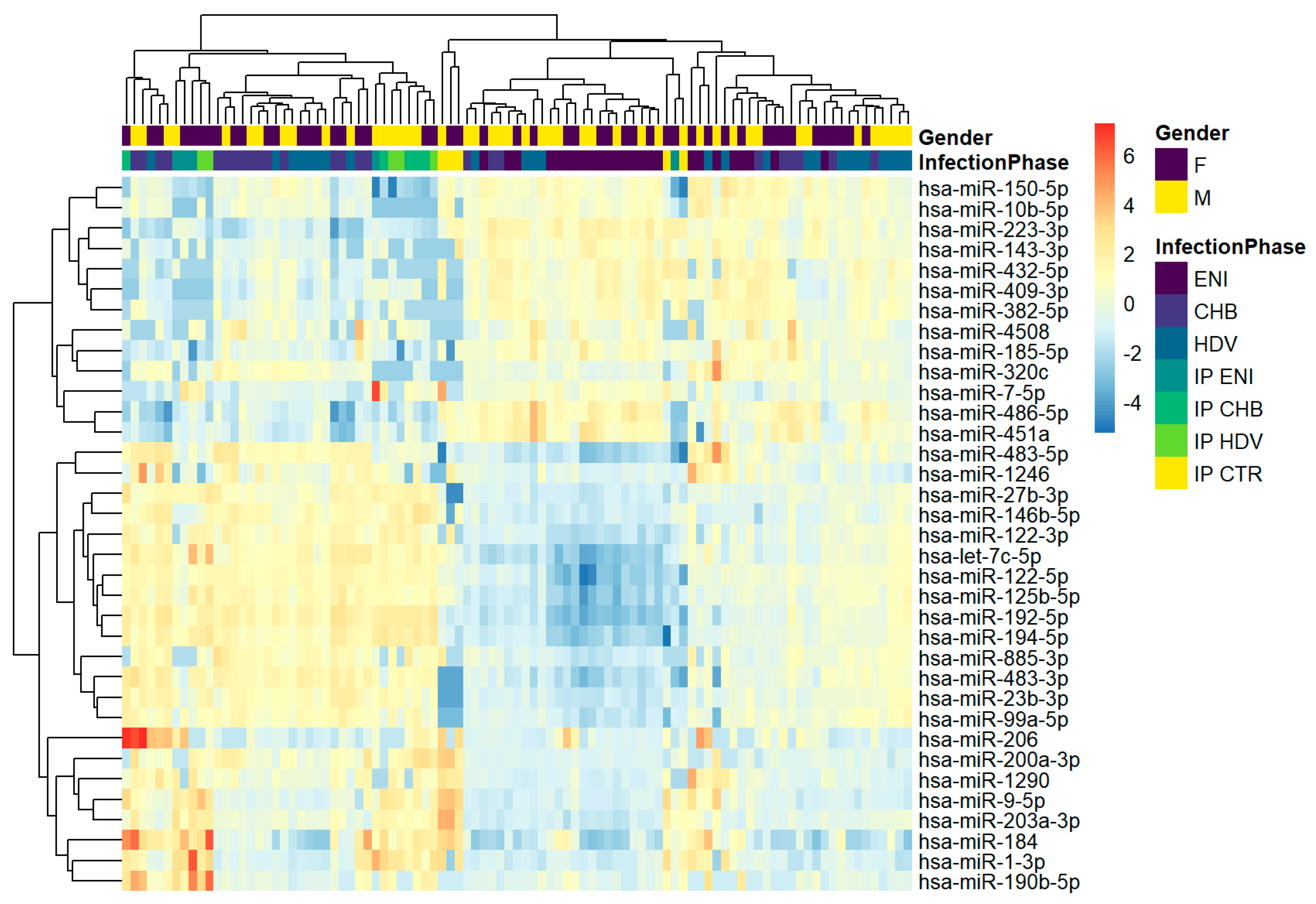
3.3. miRNA Profiling of Whole Serum
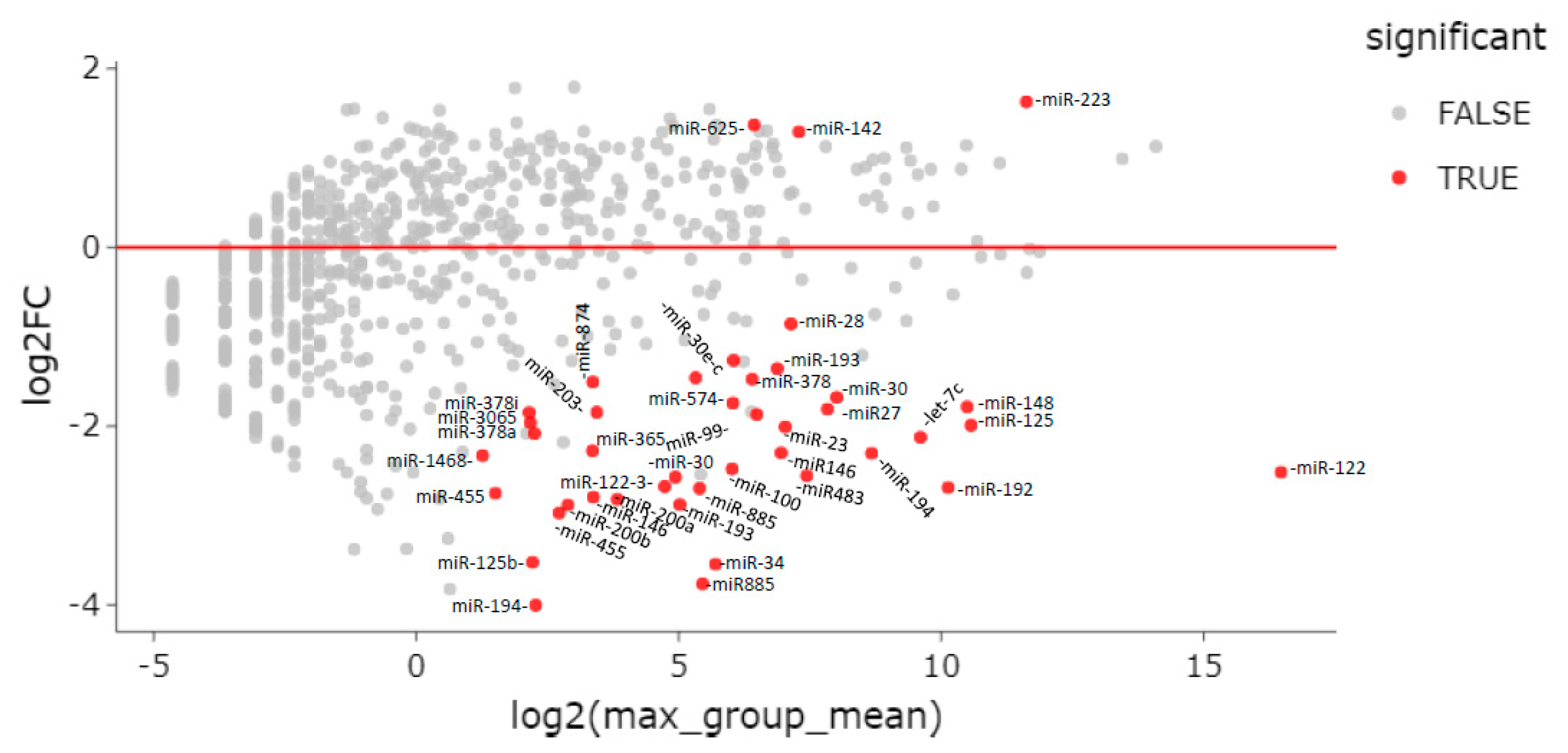
3.4. Differential miRNA Expression among Chronic Hepatitis D and B Patients and Individuals with HBeAg Negative Infection
3.5. MiR-B-Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonino, F.; Heermann, K.H.; Rizzetto, M.; Gerlich, W.H. Hepatitis delta virus: Protein composition of delta antigen and its hepatitis B virus-derived envelope. J. Virol. 1986, 58, 945–950. [Google Scholar] [CrossRef]
- Caviglia, G.P.; Ciancio, A.; Rizzetto, M. A Review of HDV Infection. Viruses 2022, 14, 1749. [Google Scholar] [CrossRef]
- Netter, H.J.; Barrios, M.H.; Littlejohn, M.; Yuen, L.K.W. Hepatitis Delta Virus (HDV) and Delta-Like Agents: Insights Into Their Origin. Front. Microbiol. 2021, 12, 652962. [Google Scholar] [CrossRef]
- Hepojoki, J.; Hetzel, U.; Paraskevopoulou, S.; Drosten, C.; Harrach, B.; Murilo, F.; Zerbini, M.; Koonin, E.V.; Krupovic, M.; Dolja, V.V.; et al. Create One New Realm (Ribozyviria) Including One New Family (Kolmioviridae) Including Genus Deltavirus and Seven New Genera for a Total of 15 Species; Report Number: ICTV [International Committee for Taxonomy of Viruses] Proposal (Taxoprop) No. 2020.012D; Research Gate: Berlin, Germany, 2021. [Google Scholar]
- Taylor, J.M. Infection by Hepatitis Delta Virus. Viruses 2020, 12, 648. [Google Scholar] [CrossRef]
- Chang, W.S.; Pettersson, J.H.; Le Lay, C.; Shi, M.; Lo, N.; Wille, M.; Eden, J.S.; Holmes, E.C. Novel hepatitis D-like agents in vertebrates and invertebrates. Virus Evol. 2019, 5, 2–10. [Google Scholar] [CrossRef]
- Pérez-Vargas, J.; Pereira de Oliveira, R.; Jacquet, S.; Pontier, D.; Cosset, F.; Freitas, N. HDV-Like Viruses. Viruses 2021, 13, 1207. [Google Scholar] [CrossRef]
- Kuo, M.Y.; Sharmeen, L.; Dinter-Gottlieb, G.; Taylor, J. Characterization of self-cleaving RNA sequence on the genome and antigenome of human hepatitis delta virus. J. Virol. 1988, 62, 4439–4444. [Google Scholar] [CrossRef]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Been, M.D. HDV ribozymes. Curr. Top. Microbiol. Immunol. 2006, 307, 47–65. [Google Scholar]
- Casey, J.L. RNA editing in hepatitis delta virus. Curr. Top. Microbiol. Immunol. 2006, 307, 67–89. [Google Scholar]
- Chang, J.; Nie, X.; Chang, H.E.; Han, Z.; Taylor, J. Transcription of hepatitis delta virus RNA by RNA polymerase II. J. Virol. 2008, 82, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Le Gal, F.; Brichler, S.; Drugan, T.; Alloui, C.; Roulot, D.; Pawlotsky, J.M.; Dény, P.; Gordien, E. Genetic diversity and worldwide distribution of the deltavirus genus: A study of 2152 clinical strains. Hepatology 2017, 66, 1826–1841. [Google Scholar] [CrossRef] [PubMed]
- Rizzetto, M.; Verme, G.; Recchia, S.; Bonino, F.; Farci, P.; Aricò, S.; Calzia, R.; Picciotto, A.; Colombo, M.; Popper, H. Chronic hepatitis in carriers of hepatitis B surface antigen, with intrahepatic expression of the delta antigen. An active and progressive disease unresponsive to immunosuppressive treatment. Ann. Intern. Med. 1983, 98, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Bonino, F.; Smedile, A. Delta Agent (Type D) hepatitis. Sem. Liver Dis. 1986, 6, 28–33. [Google Scholar] [CrossRef]
- Smedile, A.; Rosina, F.; Saracco, G.; Chiaberge, E.; Lattore, V.; Fabiano, A.; Brunetto, M.R.; Verme, G.; Rizzetto, M.; Bonino, F. Hepatitis B virus replication modulates pathogenesis of hepatitis D virus in chronic hepatitis D. Hepatology 1991, 13, 413–416. [Google Scholar] [CrossRef] [PubMed]
- Saracco, G.; Rosina, F.; Brunetto, M.R.; Amoroso, P.; Caredda, F.; Farci, P.; Piantino, P.; Bonino, F.; Rizzetto, M. Rapidly progressive HBsAg-positive hepatitis in Italy. The role of hepatitis delta virus infection. J. Hepatol. 1987, 5, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Ricco, G.; Popa, D.C.; Cavallone, D.; Iacob, S.; Salvati, A.; Tabacelia, D.; Oliveri, F.; Mascolo, G.; Bonino, F.; Yuan, Q.; et al. Quantification of serum markers of hepatitis B (HBV) and Delta virus (HDV) infections in patients with chronic HDV infection. J. Viral Hepat. 2018, 25, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Raimondo, G.; Brunetto, M.R.; Pontisso, P.; Smedile, A.; Maina, A.M.; Saitta, C.; Squadrito, G.; Tono, N. Longitudinal evaluation reveals a complex spectrum of virological profiles in hepatitis B virus/hepatitis C virus-coinfected patients. Hepatology 2006, 43, 100–107. [Google Scholar] [CrossRef]
- Schaper, M.; Rodriguez-Frias, F.; Jardi, R.; Tabernero, D.; Homs, M.; Riuz, G.; Quer, J.; Esteban, R.; Buti, M. Quantitative longitudinal evaluations of hepatitis delta virus RNA and hepatitis B virus DNA shows a dynamic, complex replicative profile in chronic hepatitis B and D. J. Hepatol. 2010, 52, 658–664. [Google Scholar]
- Novellino, L.; Rossi, R.L.; Bonino, F.; Cavallone, D.; Abrignani, S.; Pagani, M.; Brunetto, M.R. Circulating hepatitis B surface antigen particles carry hepatocellular microRNAs. PLoS ONE 2012, 7, e31952. [Google Scholar] [CrossRef]
- Blondal, T.; Brunetto, M.R.; Cavallone, D.; Mikkelsen, M.; Thorsen, M.; Mang, Y.; Pinheiro, H.; Bonino, F.; Mouritzen, P. Genome-Wide Comparison of Next-Generation Sequencing and qPCR Platforms for microRNA Profiling in Serum. Methods Mol. Biol. 2017, 1580, 21–44. [Google Scholar]
- Brunetto, M.R.; Cavallone, D.; Oliveri, F.; Moriconi, F.; Colombatto, P.; Coco, B.; Ciccorossi, P.; Rastelli, C.; Romagnoli, V.; Cherubini, B.; et al. A serum MicroRNA signature is associated with the immune control of chronic hepatitis B virus infection. PLoS ONE 2014, 9, e110782. [Google Scholar] [CrossRef] [PubMed]
- Lampertico, P.; Agarwal, K.; Berg, T.; Buti, M.; Janssen, H.L.; Papatheodoridis, G.; Zoulim, F.; Tacke, F. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J. Hepatol. 2017, 67, 370–398. [Google Scholar] [CrossRef] [PubMed]
- Brunetto, M.R.; Oliveri, F.; Colombatto, P.; Moriconi, F.; Ciccorossi, P.; Coco, B.; Romagnoli, V.; Cherubini, B.; Moscato, G.; Maina, A.M.; et al. Hepatitis B surface antigen serum levels help to distinguish active from inactive hepatitis B virus genotype D carriers. Gastroenterology 2010, 139, 483–490. [Google Scholar] [CrossRef]
- Oliveri, F.; Surace, L.; Cavallone, D.; Ricco, G.; Salvati, N.; Coco, B.; Romagnoli, V.; Gattai, R.; Salvati, A.; Moriconi, F.; et al. Long-term outcome of inactive and active, low viremic HB eAg-negative hepatitis B virus infection: Benign course towards HBsAg clearance. Liver Int. 2017, 37, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; Smyth, G.K. Small-sample estimation of negative binomial dispersion, with applications to SAGE data. Biostatistics 2008, 9, 321–332. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Chen, Y.; Zhou, X. The roles of microRNAs in epigenetic regulation. Curr. Opin. Chem. Biol. 2019, 51, 11–17. [Google Scholar] [CrossRef]
- Wang, X.; He, Y.; Mackowiak, B.; Gao, B. MicroRNAs as regulators, biomarkers and therapeutic targets in liver diseases. Gut 2021, 70, 784–795. [Google Scholar] [CrossRef]
- Ottobrelli, A.; Marzano, A.; Smedile, A.; Recchia, S.; Salizzoni, M.; Cornu, C.; Lamy, M.E.; Otte, J.B.; De Hemptinne, B.; Geubel, A.; et al. Patterns of hepatitis delta virus reinfection and disease in liver transplantation. Gastroenterology 1991, 101, 1649–1655. [Google Scholar] [CrossRef]
- Samuel, D.; Zignego, A.L.; Reynes, M.; Feray, C.; Arulnaden, J.L.; David, M.F.; Gigou, M.; Bismuth, A.; Mathieu, D.; Gentilini, P.; et al. Long-term clinical and virological outcome after liver transplantation for cirrhosis caused by chronic delta hepatitis. Hepatology 1995, 21, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Mederacke, I.; Filmann, N.; Yurdaydin, C.; Bremer, B.; Puls, F.; Zacher, B.J.; Heidrich, B.; Tillmann, H.L.; Rosenau, J.; Bock, C.T.; et al. Rapid early HDV RNA decline in the peripheral blood, but prolonged intrahepatic hepatitis delta antigen persistence after liver transplantation. J. Hepatol. 2012, 56, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Freitas, N.; Cunha, C.; Menne, S.; Gudima, S.O. Envelope proteins derived from naturally integrated hepatitis B virus DNA support assembly and release of infectious hepatitis delta virus particles. J. Virol. 2014, 88, 5742–5754. [Google Scholar] [CrossRef] [PubMed]
- Giersch, K.; Bhadra, O.D.; Volz, T.; Allweiss, L.; Riecken, K.; Fehse, B.; Lohse, A.W.; Petersen, J.; Sureau, C.; Urban, S.; et al. Hepatitis delta virus persists during liver regeneration and is amplified through cell division both in vitro and in vivo. Gut 2019, 68, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Netter, H.J.; Gerin, J.L.; Tennant, B.C.; Taylor, J.M. Apparent helper-independent infection of woodchucks by hepatitis delta virus and subsequent rescue with woodchuck hepatitis virus. J. Virol. 1994, 68, 5344–5350. [Google Scholar] [CrossRef]
- Perez-Vargas, J.; Amirache, F.; Boson, B.; Mialon, C.; Freitas, N.; Sureau, C.; Fusil, F.; Cosset, F.L. Enveloped viruses distinct from HBV induce dissemination of hepatitis D virus in vivo. Nat. Comm. 2019, 10, 2098. [Google Scholar] [CrossRef]
- Pflüger, L.S.; Schulze, Z.W.J.; Polywka, S.; Lütgehetmann, M. Hepatitis delta virus propagation enabled by hepatitis C virus—Scientifically intriguing, but is it relevant to clinical practice? J. Viral Hepat. 2020, 28, 213–216. [Google Scholar] [CrossRef]
- Verma, K.; Jyotsana, N.; Buenting, I.; Luther, S.; Pfanne, A.; Thum, T.; Ganser, A.; Heuser, M.; Weissinger, E.M.; Hambach, L. MiR-625-3p is upregulated in CD8+ T cells during early immune reconstitution after allogeneic stem cell transplantation. PLoS ONE 2017, 12, e0183828. [Google Scholar] [CrossRef]
- Bayomy, N.R.; Abo Alfottoh, W.M.; Ali Eldeep, S.A.; Ibrahim Mabrouk Mersal, A.M.S.; Abd El-Bary, H.M.A.; Abd El Gayed, E.M. Mir-142-5p as an indicator of autoimmune processes in childhood idiopathic nephrotic syndrome and as a part of MicroRNAs expression panels for its diagnosis and prediction of response to steroid treatment. Mol. Immunol. 2022, 141, 21–32. [Google Scholar] [CrossRef]
- Yuan, X.; Berg, N.; Lee, J.W.; Le, T.T.; Neudecker, V.; Jing, N.; Eltzschig, H. MicroRNA miR-223 as regulator of innate immunity. J. Leukoc. Biol. 2018, 104, 515–524. [Google Scholar] [CrossRef]
- Bonino, F.; Colombatto, P.; Brunetto, M.R. HBeAg-Negative/Anti-HBe-Positive Chronic Hepatitis B: A 40-Year-Old History. Viruses 2022, 14, 1691. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Hamada-Tsutsumi, S.; Yamamoto, Y.; Kogure, A.; Yoshioka, Y.; Watashi, K.; Ochiya, T.; Tanaka, Y. Screening of microRNAs for a repressor of hepatitis B virus replication. Oncotarget 2018, 9, 29857–29868. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.Y.; Chen, H.L.; Shih, C. MicroRNA miR-204 and miR-1236 inhibit hepatitis B virus replication via two different mechanisms. Sci. Rep. 2016, 6, 34740–34750. [Google Scholar] [CrossRef]
- Shi, T.; Hua, Q.; Ma, Z.; Lv, Q. Downregulation of miR-200a-3p induced by hepatitis B Virus X (HBx) Protein promotes cell proliferation and invasion in HBV-infection-associated hepatocarcinoma. Pathol. Res. Pract. 2017, 213, 1464–1469. [Google Scholar] [CrossRef]
- Li, W.; Li, Y.; Li, P.; Ma, F.; Liu, M.; Kong, S.; Xue, H. miR-200a-3p and miR-181-5p Mediated HOXB5 Upregulation Promotes HCC Progression by Transcriptional Activation of EGFR. Front. Oncol. 2022, 29, 822760. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ge, G.; Pan, T.; Wen, D.; Chen, L.; Yu, X.; Zhou, X.; Gan, J. A serum microRNA panel as potential biomarkers for hepatocellular carcinoma related with hepatitis B virus. PLoS ONE 2014, 9, e107986. [Google Scholar] [CrossRef]
- Pascut, D.; Cavalletto, L.; Pratama, M.Y.; Bresolin, S.; Trentin, L.; Basso, G.; Bedogni, G.; Tiribelli, C.; Chemello, L. Serum miRNA Are Promising Biomarkers for the Detection of Early Hepatocellular Carcinoma after Treatment with Direct-Acting Antivirals. Cancers 2019, 11, 1773. [Google Scholar] [CrossRef]
- Papatheodoridis, G.; Dalekos, G.; Sypsa, V.; Yurdaydin, C.; Buti, M.; Goulis, J.; Calleja, J.L.; Chi, H.; Manolakopoulos, S.; Mangia, G.; et al. PAGE-B predicts the risk of developing hepatocellular carcinoma in Caucasians with chronic hepatitis B on 5-year antiviral therapy. J. Hepatol. 2016, 64, 800–806. [Google Scholar] [CrossRef]
- Papatheodoridis, G.V.; Chan, H.L.; Hansen, B.E.; Janssen, H.L.; Lampertico, P. Risk of hepatocellular carcinoma in chronic hepatitis B: Assessment and modification with current antiviral therapy. J. Hepatol. 2015, 62, 956–967. [Google Scholar] [CrossRef]
- Liang, G.; Wu, Y.; Guan, Y.; Dong, Y.; Jiang, L.; Mao, G.; Wu, R.; Huang, Z.; Jiang, H.; Qi, L.; et al. The correlations between the serum expression of miR-206 and the severity and prognosis of sepsis. Ann. Palliat. Med. 2020, 9, 3222–3234. [Google Scholar] [CrossRef]
- Bandiera, S.; Pfeffer, S.; Baumert, T.F.; Zeisel, B.M. miR-122—A key factor and therapeutic target n liver disease. J. Hepatol. 2015, 62, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Cheung, O.; Puri, P.; Eicken, C.; Contos, M.J.; Mirshahi, F.; Maher, W.J.; Kellum, J.M.; Min, H.; Luketic, A.V.; Sanyal, A.J. Nonalcoholic steatohepatitis is associated with altered hepatic microRNA expression. Hepatology 2008, 48, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Qiu, L.; Yan, X.; Jin, W.; Wang, Y.; Chen, L.; Wu, E.; Ye, X.; Gao, F.G.; Wang, F.; et al. Loss of microRNA 122 expression in patients with hepatitis B enhances hepatitis B virus replication through cyclin G(1)—Modulated P53 activity. Hepatology 2012, 55, 730–741. [Google Scholar] [CrossRef]
- Fan, C.G.; Wang, C.M.; Tian, C.; Wang, Y.; Li, L.; Sun, W.S.; Li, R.F.; Liu, Y.G. miR-122 inhibits viral replication and cell proliferation in hepatitis B virus-related hepatocellular carcinoma and targets NDRG3. Oncol. Rep. 2011, 26, 1281–1286. [Google Scholar] [PubMed]
- Chen, Y.; Shen, A.; Rider, P.J.; Yu, Y.; Wu, K.; Mu, Y.; Hao, Q.; Liu, Y.; Gong, H.; Zhu, Y.; et al. A liver-specific microRNA binds to a highly conserved RNA sequence of hepatitis B virus and negatively regulates viral gene expression and replication. FASEB J. 2011, 25, 4511–4521. [Google Scholar] [CrossRef] [PubMed]
- Ji, F.; Yang, B.; Peng, X.; Ding, H.; You, H.; Tien, P. Circulating microRNAs in hepatitis B virus-infected patients. J. Viral Hepat. 2011, 18, e242–e251. [Google Scholar] [CrossRef] [PubMed]
- Hayes, C.N.; Akamatsu, S.; Tsuge, M.; Miki, D.; Akiyama, R.; Abe, H.; Ochi, H.; Hiraga, N.; Imamura, M.; Takahashi, S.; et al. Virus-Specific miRNAs and Argonaute2 Play a Role in the Viral Life Cycle. PLoS ONE 2012, 10, e47490. [Google Scholar] [CrossRef]
- Akuta, N.; Suzuki, F.; Kobayashi, M.; Hosaka, T.; Fujiyama, S.; Kawamura, Y.; Sezaki, H.; Kobayashi, M.; Saitoh, S.; Suzuki, Y.; et al. Circulating microRNA-122 levels are important predictor of hepatitis B virus surface antigen seroclearance. J. Med. Vir. 2018, 90, 1586–1592. [Google Scholar] [CrossRef]
- Lin, Y.; Deng, W.; Pang, J.; Kemper, T.; Hu, J.; Yin, J.; Zhang, J.; Lu, M. The microRNA-99 family modulates hepatitis B virus replication by promoting IGF-1R/PI3K/Akt/mTOR/ULK1 signaling induced autophagy. Cell. Microbiol. 2017, 19, e12709. [Google Scholar] [CrossRef]
- Zhou, J.; Yu, L.; Gao, X.; Hu, J.; Wang, J.; Dai, Z.; Wang, J.F.; Zhang, Z.; Lu, S.; Huang, X.; et al. Plasma microRNA panel to diagnose hepatitis B virus-related hepatocellular carcinoma. J. Clin. Oncol. 2011, 29, 4781–4788. [Google Scholar] [CrossRef]
- Li, L.M.; Hu, Z.B.; Zhou, Z.X.; Chen, X.; Liu, F.Y.; Zhang, J.F.; Shen, H.B.; Zhang, C.Y.; Zen, K. Serum microRNA profiles serve as novel biomarkers for HBV infection and diagnosis of HBV-positive hepatocarcinoma. Cancer Res. 2010, 70, 9798–9807. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENI (25) | CHB (25) | CHD (25) | p | ||
|---|---|---|---|---|---|
| Age | years | 49.4 (28.5/70.1) | 44.8 (31.8/70.6) | 38.0 (17.9/60.4) | ENI vs. CHB: p = 0.184 ENI vs. CHD: p < 0.003 CHB vs. CHD: p = 0.017 Overall: p = 0.004 |
| Gender | F M | 13 (52.0) 12 (48.0) | 12 (48.0) 13 (52.0) | 13 (52.0) 12 (48.0) | ENI vs. CHB: p = 1.000 ENI vs. CHD: p = 1.000 CHB vs. CHD: p = 1.000 Overall: p = 0.948 |
| Nationality | Italian Non-Italian | 20 (80) 5 (20) | 21 (84) 4 (16) | 5 (20) 20 (80) | ENI vs. CHB: p = 1.000 ENI vs. CHD: p < 0.001 CHB vs. CHD: p < 0.001 Overall: p < 0.001 |
| ALT (U/L) | Median (range) | 21 (9/33) | 110 (40/695) | 65 (19/176) | ENI vs. CHB: p < 0.001 ENI vs. CHD: p < 0.001 CHB vs. CHD: p = 0.001 Overall: p < 0.001 |
| HBV Genotype | D Non-D unknown | 25 (100) / / | 25 (100) / / | 10 (83.3) 2 (16.7) 13 | ENI vs. CHB: / ENI vs. CHD: p = 0.035 CHB vs. CHD: p = 0.035 Overall: p = 0.012 |
| HBsAg | Log IU/mL | 2.71 (1.75/3.10) | 3.75 (3.05/4.22) | 3.89 (3.09/4.59) | ENI vs. CHB: p < 0.001 ENI vs. CHD: p < 0.001 CHB vs. CHD: p = 0.273 Overall: p < 0.001 |
| HBV DNA | Log IU/mL | 2.27 (0.70/3.26) | 6.23 (4.61/8.04) | 1.0 (0.70/7.61) | ENI vs. CHB: p < 0.001 ENI vs. CHD: p < 0.001 CHB vs. CHD: p < 0.001 Overall: p < 0.001 |
| Assay | Max Groupmean | Log2 Foldchange | Foldchange | p-Value | FDR p-Value | Bonferroni |
|---|---|---|---|---|---|---|
| hsa-miR-34a-5p | 52.00 | −3.54 | −11.66 | 1.6 × 10−30 | 8.5 × 10−28 | 2.0 × 10−27 |
| hsa-miR-885-3p | 43.84 | −3.76 | −13.59 | 2.9 × 10−30 | 8.5 × 10−28 | 3.7 × 10−27 |
| hsa-miR-192-5p | 1121.40 | −2.69 | −6.44 | 8.2 × 10−30 | 1.6 × 10−20 | 1.0 × 10−19 |
| hsa-miR-148-3p | 1441.72 | −1.78 | −3.44 | 3.0 × 10−21 | 4.1 × 10−19 | 3.7 × 10−18 |
| hsa-miR-27b-3p | 227.80 | −1.81 | −3.51 | 3.5 × 10−21 | 4.1 × 10−19 | 4.5 × 10−18 |
| hsa-miR-193b-5p | 32.44 | −2.88 | −7.35 | 1.5 × 10−20 | 1.3 × 10−18 | 1.9 × 10−17 |
| hsa-miR-194-5p | 408.28 | −2.30 | −4.94 | 1.6 × 10−20 | 1.3 × 10−18 | 2.0 × 10−17 |
| hsa-miR-885-5p | 42.12 | −2.70 | −6.48 | 8.3 × 10−19 | 5.4 × 10−17 | 1.0 × 10−15 |
| hsa-miR-100-5p | 64.68 | −2.48 | −5.57 | 8.4 × 10−19 | 5.4 × 10−17 | 1.1 × 10−15 |
| hsa-miR-30a-3p | 30.60 | −2.57 | −5.94 | 2.3 × 10−18 | 1.3 × 10−16 | 2.9 × 10−15 |
| hsa-miR-146b-5p | 123.68 | −2.30 | −4.92 | 3.3 × 10−18 | 1.8 × 10−16 | 4.2 × 10−15 |
| hsa-miR-122-5p | 90,951.56 | −2.51 | −5.71 | 3.7 × 10−18 | 1.8 × 10−16 | 4.6 × 10−15 |
| hsa-miR-30a-5p | 257.48 | −1.68 | −3.20 | 7.8 × 10−18 | 3.5 × 10−16 | 9.8 × 10−15 |
| hsa-miR-483-3p | 173.36 | −2.55 | −5.87 | 4.8 × 10−17 | 2.0 × 10−15 | 6.0 × 10−14 |
| hsa-miR-125b-5p | 1516.76 | −1.99 | −3.97 | 4.6 × 10−16 | 1.8 × 10−14 | 5.8 × 10−13 |
| hsa-miR-23b-3p | 130.60 | −2.01 | −4.02 | 1.3 × 10−14 | 4.8 × 10−13 | 1.7 × 10−11 |
| hsa-miR-146b-3p | 10.36 | −2.79 | −6.93 | 3.0 × 10−14 | 1.0 × 10−12 | 3.7 × 10−11 |
| hsa-miR-378a-3p | 84.44 | −1.48 | −2.78 | 1.0 × 10−13 | 3.4 × 10−12 | 1.3 × 10−10 |
| hsa-let-7c-5p | 778.48 | −2.12 | −4.36 | 2.4 × 10−13 | 7.2 × 10−12 | 3.0 × 10−10 |
| hsa-miR-193a-5p | 117.48 | −1.36 | −2.56 | 2.6 × 10−13 | 7.4 × 10−12 | 3.2 × 10−10 |
| hsa-miR-194-3p | 4.84 | −4.00 | −16.02 | 5.7 × 10−13 | 1.6 × 10−11 | 7.2 × 10−10 |
| hsa-miR-125b-2-3p | 4.64 | −3.52 | −11.49 | 6.0 × 10−13 | 1.6 × 10−11 | 7.6 × 10−10 |
| hsa-miR-200a-3p | 14.20 | −2.82 | −7.05 | 1.2 × 10−12 | 3.1 × 10−11 | 1.6 × 10−9 |
| hsa-miR-99a-5p | 89.64 | −1.87 | −3.65 | 1.4 × 10−12 | 3.3 × 10−11 | 1.7 × 10−9 |
| hsa-miR-122-3p | 26.60 | −2.67 | −6.39 | 1.7 × 10−12 | 3.9 × 10−11 | 2.1 × 10−9 |
| hsa-miR-30c-5p | 65.96 | −1.26 | −2.40 | 5.6 × 10−12 | 1.3 × 10−10 | 7.1 × 10−9 |
| hsa-miR-200b-3p | 7.40 | −2.88 | −7.37 | 1.3 × 10−11 | 2.8 × 10−10 | 1.6 × 10−8 |
| hsa-miR-365a-3p | 10.24 | −2.27 | −4.84 | 1.6 × 10−11 | 3.4 × 10−10 | 2.1 × 10−8 |
| hsa-miR-455-3p | 6.60 | −2.97 | −7.84 | 2.7 × 10−11 | 5.4 × 10−10 | 3.4 × 10−8 |
| hsa-miR-574-3p | 65.44 | −1.74 | −3.35 | 4.0 × 10−11 | 7.7 × 10−10 | 5.0 × 10−8 |
| hsa-miR-378a-5p | 4.76 | −2.08 | −4.23 | 1.2 × 10−8 | 2.2 × 10−7 | 1.5 × 10−6 |
| hsa-miR-223-3p | 3152.68 | 1.63 | 3.09 | 1.3 × 10−8 | 2.3 × 10−7 | 1.6 × 10−5 |
| hsa-miR-455-3p | 2.84 | −2.75 | −6.73 | 2.4 × 10−8 | 4.2 × 10−7 | 3.0 × 10−5 |
| hsa-miR-203a-3p | 10.84 | −1.85 | −3.59 | 3.4 × 10−8 | 5.8 × 10−7 | 4.3 × 10−5 |
| hsa-miR-142-5p | 156.32 | 1.29 | 2.45 | 4.8 × 10−8 | 8.0 × 10−7 | 6.1 × 10−5 |
| hsa-miR-30e-3p | 39.96 | −1.46 | −2.75 | 8.1 × 10−8 | 1.3 × 10−7 | 1.0 × 10−4 |
| hsa-miR-28-3p | 140.84 | −0.86 | −1.81 | 8.6 × 10−8 | 1.3 × 10−7 | 1.1 × 10−4 |
| hsa-miR-1468-5p | 2.40 | −2.33 | −5.02 | 1.7 × 10−7 | 2.6 × 10−7 | 2.2 × 10−4 |
| hsa-miR-378i | 4.44 | −1.85 | −3.60 | 1.9 × 10−7 | 2.8 × 10−7 | 2.4 × 10−4 |
| hsa-miR-625-3p | 86.60 | 1.37 | 2.58 | 3.7 × 10−7 | 5.3 × 10−7 | 4.7 × 10−4 |
| hsa-miR-3065-5p | 4.52 | −1.96 | −3.90 | 3.8 × 10−7 | 5.4 × 10−7 | 4.8 × 10−4 |
| hsa-miR-874-3p | 10.28 | −1.51 | −2.84 | 5.6 × 10−7 | 7.7 × 10−7 | 7.0 × 10−4 |
| Assay | Max Groupmean | log2 Foldchange | Foldchange | p-Value | FDR p-Value | Bonferroni |
|---|---|---|---|---|---|---|
| hsa-miR-885-3p | 48.16 | −3.37 | −10.32 | 6.3 × 10−25 | 3.9 × 10−22 | 8.0 × 10−22 |
| hsa-miR-885-5p | 70.76 | −2.71 | −6.53 | 2.2 × 10−19 | 6.5 × 10−17 | 2.8 × 10−16 |
| hsa-miR-193b-5p | 46.92 | −2.74 | −6.68 | 3.2 × 10−19 | 6.5 × 10−17 | 4.0 × 10−16 |
| hsa-miR-100-5p | 116.84 | −2.40 | −5.28 | 5.2 × 10−18 | 7.6 × 10−16 | 6.5 × 10−15 |
| hsa-miR-34a-5p | 41.88 | −2.65 | −6.29 | 6.2 × 10−18 | 7.6 × 10−16 | 7.8 × 10−15 |
| hsa-miR-192-5p | 1675.12 | −2.32 | −5.00 | 1.8 × 10−17 | 1.9 × 10−15 | 2.3 × 10−14 |
| hsa-miR-148a-3p | 2186.08 | −1.60 | −3.02 | 2.3 × 10−17 | 2.0 × 10−15 | 2.8 × 10−14 |
| hsa-miR-122-5p | 140,835.00 | −2.31 | −4.96 | 1.4 × 10−15 | 1.0 × 10−13 | 1.7 × 10−12 |
| hsa-miR-194-5p | 600.92 | −1.97 | −3.93 | 1.6 × 10−15 | 1.1 × 10−13 | 2.0 × 10−12 |
| hsa-miR-27b-3p | 295.20 | −1.47 | −2.77 | 1.1 × 10−14 | 6.6 × 10−13 | 1.4 × 10−11 |
| hsa-miR-365a-3p | 21.28 | −2.51 | −5.70 | 1.7 × 10−14 | 9.4 × 10−13 | 2.1 × 10−11 |
| hsa-miR-483-3p | 253.28 | −2.30 | −4.93 | 3.1 × 10−14 | 1.6 × 10−12 | 3.9 × 10−11 |
| hsa-miR-23b-3p | 255.72 | −1.96 | −3.90 | 3.9 × 10−14 | 1.8 × 10−12 | 4.9 × 10−11 |
| hsa-miR-125b-5p | 2728.28 | −1.84 | −3.57 | 6.0 × 10−14 | 2.6 × 10−12 | 7.5 × 10−11 |
| hsa-miR-30a-3p | 45.40 | −2.18 | −4.54 | 6.3 × 10−14 | 2.6 × 10−12 | 8.0 × 10−11 |
| hsa-miR-99a-5p | 179.40 | −1.93 | −3.80 | 1.7 × 10−13 | 6.6 × 10−12 | 2.1 × 10−10 |
| hsa-miR-146b-5p | 175.20 | −1.91 | −3.75 | 4.3 × 10−13 | 1.6 × 10−11 | 5.5 × 10−10 |
| hsa-miR-122-3p | 54.72 | −2.66 | −6.33 | 1.3 × 10−12 | 4.4 × 10−11 | 1.6 × 10−9 |
| hsa-miR-146b-3p | 16.12 | −2.54 | −5.83 | 1.9 × 10−12 | 6.1 × 10−11 | 2.3 × 10−9 |
| hsa-let-7c-5p | 1429.64 | −2.03 | −4.09 | 2.3 × 10−12 | 7.1 × 10−11 | 2.9 × 10−9 |
| hsa-miR-194-3p | 8.08 | −3.78 | −13.76 | 4.8 × 10−12 | 1.4 × 10−10 | 6.1 × 10−9 |
| hsa-miR-30c-5p | 122.72 | −1.24 | −2.37 | 5.4 × 10−12 | 1.5 × 10−10 | 6.8 × 10−9 |
| hsa-miR-574-3p | 141.24 | −1.80 | −3.47 | 6.1 × 10−12 | 1.6 × 10−10 | 7.7 × 10−9 |
| hsa-miR-125b-2-3p | 6.44 | −3.22 | −9.29 | 2.3 × 10−11 | 5.9 × 10−10 | 2.9 × 10−8 |
| hsa-miR-378a-5p | 10.60 | −2.33 | −5.02 | 2.4 × 10−11 | 5.9 × 10−10 | 3.0 × 10−8 |
| hsa-miR-30a-5p | 335.88 | −1.29 | −2.45 | 2.9 × 10−11 | 7.0 × 10−10 | 3.7 × 10−8 |
| hsa-miR-455-5p | 10.16 | −2.66 | −6.31 | 1.4 × 10−9 | 3.2 × 10−8 | 1.8 × 10−6 |
| hsa-miR-1468-5p | 4.40 | −2.44 | −5.44 | 8.6 × 10−9 | 1.9 × 10−7 | 1.1 × 10−5 |
| hsa-miR-193a-5p | 133.12 | −1.04 | −2.06 | 1.3 × 10−8 | 2.7 × 10−7 | 1.6 × 10−5 |
| hsa-miR-204-5p | 12.04 | −1.68 | −3.20 | 6.8 × 10−8 | 1.4 × 10−6 | 8.6 × 10−5 |
| hsa-miR-345-5p | 25.48 | −1.24 | −2.36 | 8.5 × 10−8 | 1.7 × 10−6 | 1.1 × 10−4 |
| hsa-miR-28-3p | 244.56 | −0.81 | −1.75 | 3.3 × 10−7 | 6.4 × 10−6 | 4.1 × 10−4 |
| hsa-miR-99a-3p | 4.16 | −2.49 | −5.60 | 5.4 × 10−7 | 1.0 × 10−5 | 6.8 × 10−4 |
| hsa-miR-455-3p | 4.08 | −2.40 | −5.28 | 6.3 × 10−7 | 1.2 × 10−5 | 8.0 × 10−4 |
| ENI vs. CHD | ENI vs. CHB | |||
|---|---|---|---|---|
| Assay | Fold Change | Bonferroni | Fold Change | Bonferroni |
| hsa-miR-204-5p | −3.20 | 8.60 × 10−05 | −2.89 | 3.16 × 10−03 |
| hsa-miR-345-5p | −2.36 | 1.08 × 10−04 | −2.21 | 2.84 × 10−03 |
| hsa-miR-99a-3p | −5.60 | 6.75 × 10−04 | −4.89 | 1.37 × 10−02 |
| ENI vs. CHB | ENI vs. CHD | |||
|---|---|---|---|---|
| Assay | Fold Change | Bonferroni | Fold Change | Bonferroni |
| hsa-miR-378a-3p | −2.78 | 1.32 × 10−10 | −1.90 | 3.18 × 10−03 |
| hsa-miR-200a-3p | −7.05 | 1.56 × 10−09 | −2.04 | 1.00 |
| hsa-miR-200b-3p | −7.37 | 1.64 × 10−08 | −2.90 | 4.34 × 10−01 |
| hsa-miR-203a-3p | −3.59 | 4.31 × 10−05 | −3.02 | 1.44 × 10−03 |
| hsa-miR-30e-3p | −2.75 | 1.03 × 10−04 | −2.46 | 1.67 × 10−03 |
| hsa-miR-378i | −3.60 | 2.37 × 10−04 | −3.00 | 4.52 × 10−03 |
| hsa-miR-3065-5p | −3.90 | 4.82 × 10−04 | −2.40 | 1.00 |
| hsa-miR-874-3p | −2.84 | 7.03 × 10−04 | −2.19 | 1.39 × 10−01 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavallone, D.; Ornos, E.D.B.; Ricco, G.; Oliveri, F.; Coco, B.; Colombatto, P.; De Rosa, L.; Dalmacio, L.M.M.; Bonino, F.; Brunetto, M.R. The Circulating miRNA Profile of Chronic Hepatitis D and B Patients Is Comparable but Differs from That of Individuals with HBeAg-Negative HBV Infection. Viruses 2023, 15, 2257. https://doi.org/10.3390/v15112257
Cavallone D, Ornos EDB, Ricco G, Oliveri F, Coco B, Colombatto P, De Rosa L, Dalmacio LMM, Bonino F, Brunetto MR. The Circulating miRNA Profile of Chronic Hepatitis D and B Patients Is Comparable but Differs from That of Individuals with HBeAg-Negative HBV Infection. Viruses. 2023; 15(11):2257. https://doi.org/10.3390/v15112257
Chicago/Turabian StyleCavallone, Daniela, Eric David B. Ornos, Gabriele Ricco, Filippo Oliveri, Barbara Coco, Piero Colombatto, Laura De Rosa, Leslie Michelle M. Dalmacio, Ferruccio Bonino, and Maurizia Rossana Brunetto. 2023. "The Circulating miRNA Profile of Chronic Hepatitis D and B Patients Is Comparable but Differs from That of Individuals with HBeAg-Negative HBV Infection" Viruses 15, no. 11: 2257. https://doi.org/10.3390/v15112257
APA StyleCavallone, D., Ornos, E. D. B., Ricco, G., Oliveri, F., Coco, B., Colombatto, P., De Rosa, L., Dalmacio, L. M. M., Bonino, F., & Brunetto, M. R. (2023). The Circulating miRNA Profile of Chronic Hepatitis D and B Patients Is Comparable but Differs from That of Individuals with HBeAg-Negative HBV Infection. Viruses, 15(11), 2257. https://doi.org/10.3390/v15112257