
ATG8f Interacts with Chilli Veinal Mottle Virus 6K2 Protein to Limit Virus Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Virus Inoculation
2.2. Vector Construction and Agrobacterium Infiltration
2.3. Transmission Electron Microscopy (TEM)
2.4. RNA Extraction and Quantitative RT-PCR (qRT-PCR)
2.5. Protein Extraction and Western Blot Analysis
2.6. Chemical Treatments
2.7. Yeast Two-Hybrid (Y2H) Assay
2.8. Bimolecular Fluorescence Complementation (BiFC) Assay
2.9. Co-Immunoprecipitation (Co-IP) Assay
2.10. Statistical Analysis
3. Results
3.1. ChiVMV Infection Activates Autophagy in N. benthamiana
3.2. Autophagy Plays a Positive Role in the Antiviral Process of N. benthamiana
3.3. ATG8f Interacts with ChiVMV-6K2 In Vitro and In Vivo
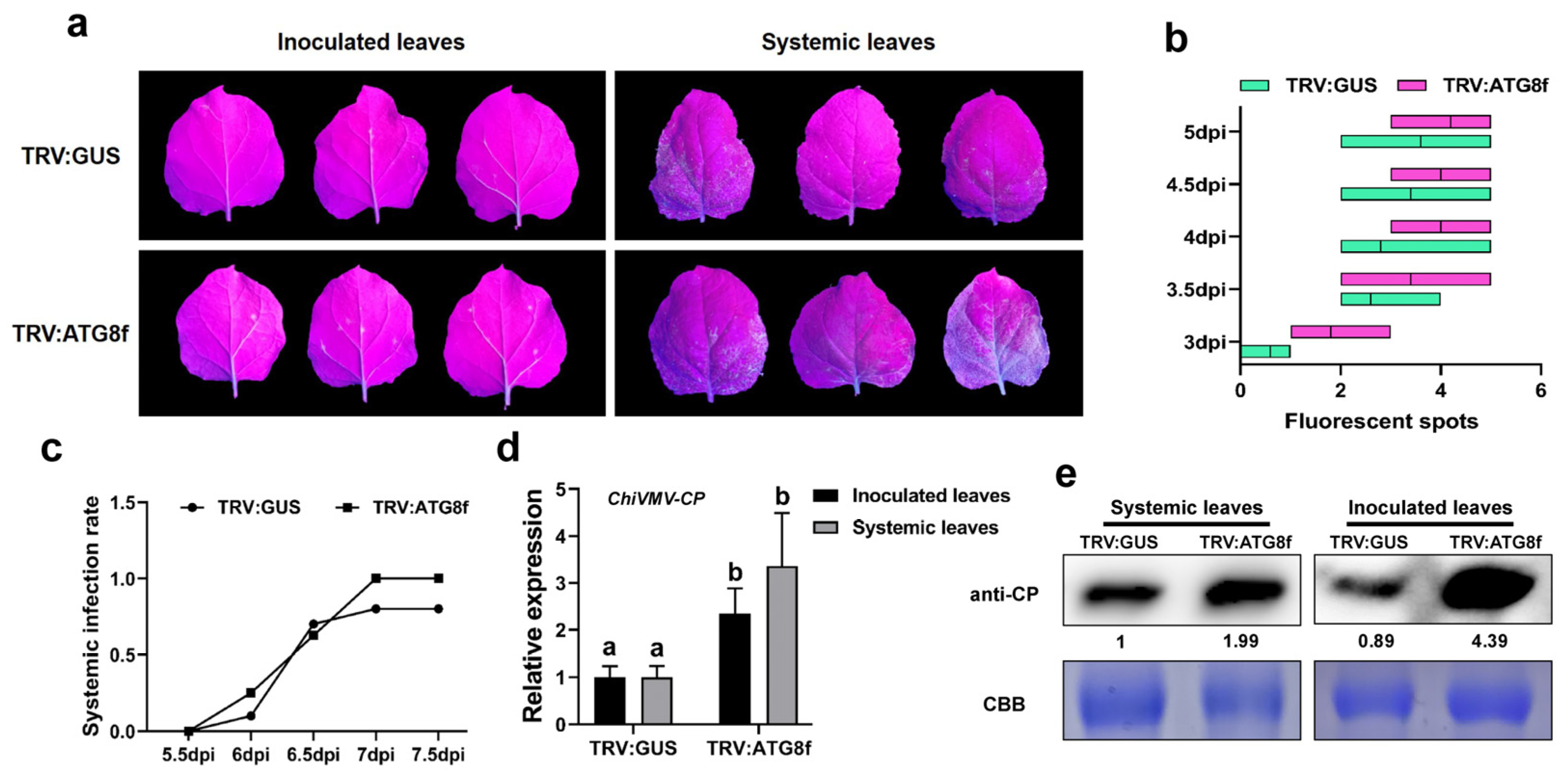
3.4. Silencing of ATG8f Promotes the Accumulation of ChiVMV-GFP
3.5. Overexpression of ATG8f Inhibits the Infection of ChiVMV-GFP
3.6. 6K2 Is Degraded by Autophagy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, L.; Chen, Y.; Tooze, S.A. Autophagy pathway: Cellular and molecular mechanisms. Autophagy 2018, 14, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhang, X.; Otegui, M.S. Plant autophagy: New flavors on the menu. Curr. Opin. Plant Biol. 2018, 46, 113–121. [Google Scholar] [CrossRef]
- Tasset, I.; Cuervo, A.M. Role of chaperone-mediated autophagy in metabolism. FEBS J. 2016, 283, 2403–2413. [Google Scholar] [CrossRef] [PubMed]
- Mijaljica, D.; Prescott, M.; Devenish, R.J. Microautophagy in mammalian cells: Revisiting a 40-year-old conundrum. Autophagy 2011, 7, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. An Overview of the Molecular Mechanism of Autophagy. In Autophagy in Infection and Immunity; Levine, B., Yoshimori, T., Deretic, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–32. [Google Scholar] [CrossRef]
- Mizushima, N.; Yoshimori, T.; Ohsumi, Y. The role of Atg proteins in autophagosome formation. Annu. Rev. Cell Dev. Biol. 2011, 27, 107–132. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 2010, 22, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Popelka, H.; Klionsky, D.J. Multiple structural rearrangements mediated by high-plasticity regions in Atg3 are key for efficient conjugation of Atg8 to PE during autophagy. Autophagy 2021, 17, 1805–1808. [Google Scholar] [CrossRef]
- Noda, N.N.; Fujioka, Y.; Hanada, T.; Ohsumi, Y.; Inagaki, F. Structure of the Atg12-Atg5 conjugate reveals a platform for stimulating Atg8-PE conjugation. EMBO Rep. 2013, 14, 206–211. [Google Scholar] [CrossRef]
- Davis, S.; Wang, J.; Ferro-Novick, S. Crosstalk between the Secretory and Autophagy Pathways Regulates Autophagosome Formation. Dev. Cell 2017, 41, 23–32. [Google Scholar] [CrossRef]
- Obara, K.; Sekito, T.; Niimi, K.; Ohsumi, Y. The Atg18-Atg2 complex is recruited to autophagic membranes via phosphatidylinositol 3-phosphate and exerts an essential function. J. Biol. Chem. 2008, 283, 23972–23980. [Google Scholar] [CrossRef] [PubMed]
- Bassham, D.C. Plant autophagy—More than a starvation response. Curr. Opin. Plant Biol. 2007, 10, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Bozhkov, P.V. Plant autophagy: Mechanisms and functions. J. Exp. Bot. 2018, 69, 1281–1285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhou, T.; Zhang, Y.; Chen, J.; Song, H.; Wu, P.; Yue, C.; Huang, J.; Zhang, Z.; Hua, Y. Genome-Wide Identification and Functional Characterization Reveals the Pivotal Roles of BnaA8.ATG8F in Salt Stress Tolerance and Nitrogen Limitation Adaptation in Allotetraploid Rapeseed. Int. J. Mol. Sci. 2022, 23, 1318. [Google Scholar] [CrossRef] [PubMed]
- Ueda, M.; Tsutsumi, N.; Fujimoto, M. Salt stress induces internalization of plasma membrane aquaporin into the vacuole in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2016, 474, 742–746. [Google Scholar] [CrossRef]
- Chen, L.; Su, Z.Z.; Huang, L.; Xia, F.N.; Qi, H.; Xie, L.J.; Xiao, S.; Chen, Q.F. The AMP-Activated Protein Kinase KIN10 Is Involved in the Regulation of Autophagy in Arabidopsis. Front. Plant Sci. 2017, 8, 1201. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, M.; Wang, E.; Hu, L.; Hawkesford, M.J.; Zhong, L.; Chen, Z.; Xu, Z.; Li, L.; Zhou, Y.; et al. Genome-wide analysis of autophagy-associated genes in foxtail millet (Setaria italica L.) and characterization of the function of SiATG8a in conferring tolerance to nitrogen starvation in rice. BMC Genom. 2016, 17, 797. [Google Scholar] [CrossRef]
- Lai, Z.; Wang, F.; Zheng, Z.; Fan, B.; Chen, Z. A critical role of autophagy in plant resistance to necrotrophic fungal pathogens. Plant J. 2011, 66, 953–968. [Google Scholar] [CrossRef]
- Maqbool, A.; Hughes, R.K.; Dagdas, Y.F.; Tregidgo, N.; Zess, E.; Belhaj, K.; Round, A.; Bozkurt, T.O.; Kamoun, S.; Banfield, M.J. Structural Basis of Host Autophagy-related Protein 8 (ATG8) Binding by the Irish Potato Famine Pathogen Effector Protein PexRD54. J. Biol. Chem. 2016, 291, 20270–20282. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef]
- Li, F.; Chung, T.; Pennington, J.G.; Federico, M.L.; Kaeppler, H.F.; Kaeppler, S.M.; Otegui, M.S.; Vierstra, R.D. Autophagic recycling plays a central role in maize nitrogen remobilization. Plant Cell 2015, 27, 1389–1408. [Google Scholar] [CrossRef]
- Buerte, B.; Zeng, Z.; Zhou, C.; Lian, G.; Guo, F.; Wang, J.; Han, N.; Zhu, M.; Bian, H. Identification of New ATG8s-Binding Proteins with Canonical LC3-Interacting Region in Autophagosomes of Barley Callus. Plant Cell Physiol. 2022, 63, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Haxim, Y.; Ismayil, A.; Jia, Q.; Wang, Y.; Zheng, X.; Chen, T.; Qian, L.; Liu, N.; Wang, Y.; Han, S.; et al. Autophagy functions as an antiviral mechanism against geminiviruses in plants. Elife 2017, 6, e23897. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, C.; Li, Y.; Wu, G.; Hou, X.; Zhou, X.; Wang, A. Beclin1 restricts RNA virus infection in plants through suppression and degradation of the viral polymerase. Nat. Commun. 2018, 9, 1268. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, M.; Zhang, C.; Zhou, X. Nuclear autophagy degrades a geminivirus nuclear protein to restrict viral infection in solanaceous plants. New Phytol. 2020, 225, 1746–1761. [Google Scholar] [CrossRef]
- Jiang, L.; Lu, Y.; Zheng, X.; Yang, X.; Chen, Y.; Zhang, T.; Zhao, X.; Wang, S.; Zhao, X.; Song, X.; et al. The plant protein NbP3IP directs degradation of Rice stripe virus p3 silencing suppressor protein to limit virus infection through interaction with the autophagy-related protein NbATG8. New Phytol. 2021, 229, 1036–1051. [Google Scholar] [CrossRef]
- Li, F.; Zhang, C.; Tang, Z.; Zhang, L.; Dai, Z.; Lyu, S.; Li, Y.; Hou, X.; Bernards, M.; Wang, A. A plant RNA virus activates selective autophagy in a UPR-dependent manner to promote virus infection. New Phytol. 2020, 228, 622–639. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Molecular biology of potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [CrossRef]
- Jiang, J.; Patarroyo, C.; Cabanillas, D.G.; Zheng, H.; Laliberté, J.F. The vesicle-forming 6K2 protein of turnip mosaic virus interacts with the COPII coatomer Sec24a for viral systemic infection. J. Virol. 2015, 89, 6695–6710. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Shen, L.; Qin, Y.; Liu, X.; Hao, K.; Li, Y.; Wang, J.; Yang, J.; Wang, F. CLC-Nt1 affects Potato Virus Y infection via regulation of endoplasmic reticulum luminal Ph. New Phytol. 2018, 220, 539–552. [Google Scholar] [CrossRef]
- Peng, Q.; Yang, D.; Yang, T.; Cheng, Y.; Yang, Y.; Xi, D. Construction of full-length cDNA infectious clones of Chilli veinal mottle virus. Virus Res. 2022, 322, 198948. [Google Scholar] [CrossRef] [PubMed]
- Quenouille, J.; Vassilakos, N.; Moury, B. Potato virus Y: A major crop pathogen that has provided major insights into the evolution of viral pathogenicity. Mol. Plant Pathol. 2013, 14, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Hafrén, A.; Üstün, S.; Hochmuth, A.; Svenning, S.; Johansen, T.; Hofius, D. Turnip Mosaic Virus Counteracts Selective Autophagy of the Viral Silencing Suppressor HCpro. Plant Physiol. 2018, 176, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, A. The Potyvirus Silencing Suppressor Protein VPg Mediates Degradation of SGS3 via Ubiquitination and Autophagy Pathways. J. Virol. 2017, 91, e01478-16. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, T.T.; Zhong, J.; Zhao, L.L.; Guo, Y.L.; Lin, Y.C.; Gao, G.; Cao, Y.; Ding, M. First report of chilli veinal mottle virus infecting tobacco in Guizhou, China. J. Plant Pathol. 2022, 104, 1159–1160. [Google Scholar] [CrossRef]
- Yang, T.; Qiu, L.; Huang, W.; Xu, Q.; Zou, J.; Peng, Q.; Lin, H.; Xi, D. Chilli veinal mottle virus HCPro interacts with catalase to facilitate virus infection in Nicotiana tabacum. J. Exp. Bot. 2020, 71, 5656–5668. [Google Scholar] [CrossRef]
- Laprom, A.; Nilthong, S.; Chukeatirote, E. Incidence of viruses infecting pepper in Thailand. Biomol. Concepts 2019, 10, 184–193. [Google Scholar] [CrossRef]
- Zhu, F.; Xi, D.H.; Yuan, S.; Xu, F.; Zhang, D.W.; Lin, H.H. Salicylic acid and jasmonic acid are essential for systemic resistance against tobacco mosaic virus in Nicotiana benthamiana. Mol. Plant Microbe Interact. 2014, 27, 567–577. [Google Scholar] [CrossRef]
- Zhou, X.; Zhu, T.; Zhu, L.-S.; Luo, S.-S.; Deng, X.-G.; Lin, H.-H.; Xi, D.-H. The Role of Photoreceptors in Response to Cucumber Mosaic Virus in Arabidopsis thaliana. J. Plant Growth Regul. 2017, 36, 257–270. [Google Scholar] [CrossRef]
- Yang, D.; Peng, Q.; Cheng, Y.; Xi, D. Glucose-6-phosphate dehydrogenase promotes the infection of Chilli veinal mottle virus through affecting ROS signaling in Nicotiana benthamiana. Planta 2022, 256, 96. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, J.; Zhao, F.; Ren, H.; Zhu, L.; Xi, D.; Lin, H. The interaction between Turnip crinkle virus p38 and Cucumber mosaic virus 2b and its critical domains. Virus Res. 2016, 222, 94–105. [Google Scholar] [CrossRef]
- Huang, Y.P.; Huang, Y.W.; Hsiao, Y.J.; Li, S.C.; Hsu, Y.H.; Tsai, C.H. Autophagy is involved in assisting the replication of Bamboo mosaic virus in Nicotiana benthamiana. J. Exp. Bot. 2019, 70, 4657–4670. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The Master of Bulk and Selective Recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, Y.; Xie, X.; Yue, N.; Li, J.; Wang, X.B.; Han, C.; Yu, J.; Liu, Y.; Li, D. Barley stripe mosaic virus γb Protein Subverts Autophagy to Promote Viral Infection by Disrupting the ATG7–ATG8 Interaction. Plant Cell 2018, 30, 1582–1595. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Chen, Q.; Havé, M. Regulation of nutrient recycling via autophagy. Curr. Opin. Plant Biol. 2017, 39, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Hafrén, A.; Macia, J.L.; Love, A.J.; Milner, J.J.; Drucker, M.; Hofius, D. Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles. Proc. Natl. Acad. Sci. USA 2017, 114, E2026–E2035. [Google Scholar] [CrossRef]
- Cui, H.G.; Wang, A.M. Plum pox virus 6K1 protein Is required for viral replication and targets the viral replication complex at the early stage of infection. J. Virol. 2016, 90, 5119–5131. [Google Scholar] [CrossRef]
- Wu, G.W.; Jia, Z.X.; Ding, K.D.; Zheng, H.Y.; Lu, Y.W.; Lin, L.; Peng, J.J.; Rao, S.F.; Wang, A.M.; Chen, J.P.; et al. Turnip mosaic virus co-opts the vacuolar sorting receptor VSR4 to promote viral genome replication in plants by targeting viral replication vesicles to the endosome. PLoS Pathog. 2022, 18, e1010257. [Google Scholar] [CrossRef] [PubMed]
- Grangeon, R.; Jiang, J.; Wan, J.; Agbeci, M.; Zheng, H.; Laliberté, J.F. 6K2-induced vesicles can move cell to cell during turnip mosaic virus infection. Front. Microbiol. 2013, 4, 351. [Google Scholar] [CrossRef]
- Jiang, T.; Du, K.; Wang, P.; Wang, X.; Zang, L.; Peng, D.; Chen, X.; Sun, G.; Zhang, H.; Fan, Z.; et al. Sugarcane mosaic virus orchestrates the lactate fermentation pathway to support its successful infection. Front. Plant Sci. 2022, 13, 1099362. [Google Scholar] [CrossRef] [PubMed]
- Niu, E.; Liu, H.; Zhou, H.; Luo, L.; Wu, Y.; Andika, I.B.; Sun, L. Autophagy Inhibits Intercellular Transport of Citrus Leaf Blotch Virus by Targeting Viral Movement Protein. Viruses 2021, 13, 2189. [Google Scholar] [CrossRef]
- Cheng, X.; Xiong, R.; Li, Y.; Li, F.; Zhou, X.; Wang, A. Sumoylation of Turnip mosaic virus RNA Polymerase Promotes Viral Infection by Counteracting the Host NPR1-Mediated Immune Response. Plant Cell 2017, 29, 508–525. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Yang, Z.; Zhang, H.; Zhang, J.; Xu, J. Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol. 2020, 225, 2122–2139. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, C.; Zhou, J.; Yang, D.; Yuan, B.; Tang, R.; Liu, Y.; Xi, D. ATG8f Interacts with Chilli Veinal Mottle Virus 6K2 Protein to Limit Virus Infection. Viruses 2023, 15, 2324. https://doi.org/10.3390/v15122324
Ji C, Zhou J, Yang D, Yuan B, Tang R, Liu Y, Xi D. ATG8f Interacts with Chilli Veinal Mottle Virus 6K2 Protein to Limit Virus Infection. Viruses. 2023; 15(12):2324. https://doi.org/10.3390/v15122324
Chicago/Turabian StyleJi, Chenglong, Jingya Zhou, Daoyong Yang, Bowen Yuan, Rongxia Tang, Yong Liu, and Dehui Xi. 2023. "ATG8f Interacts with Chilli Veinal Mottle Virus 6K2 Protein to Limit Virus Infection" Viruses 15, no. 12: 2324. https://doi.org/10.3390/v15122324