
A Renaissance for Oncolytic Adenoviruses?


1
Goodman Cancer Institute, McGill University, Montréal, QC H3A 1A3, Canada
2
Department of Biochemistry, McGill University, Montréal, QC H3G 1Y6, Canada
3
Department of Microbiology and Immunology, McGill University, Montréal, QC H3G 1Y6, Canada
*
Author to whom correspondence should be addressed.
Viruses 2023, 15(2), 358; https://doi.org/10.3390/v15020358
Submission received: 21 November 2022
/
Revised: 18 January 2023
/
Accepted: 22 January 2023
/
Published: 26 January 2023
(This article belongs to the Special Issue New Aspects of Adenoviral Vaccine Vectors and Adenoviral Gene Therapy)
Abstract
:In the 1990s, adenovirus became one of the first virus types to be genetically engineered to selectively destroy cancer cells. In the intervening years, the field of “oncolytic viruses” has slowly progressed and culminated in 2015 with the FDA approval of Talimogene laherparepvec, a genetically engineered herpesvirus, for the treatment of metastatic melanoma. Despite the slower progress in translating oncolytic adenovirus to the clinic, interest in the virus remains strong. Among all the clinical trials currently using viral oncolytic agents, the largest proportion of these are using recombinant adenovirus. Many trials are currently underway to use oncolytic virus in combination with immune checkpoint inhibitors (ICIs), and early results using oncolytic adenovirus in this manner are starting to show promise. Many of the existing strategies to engineer adenoviruses were designed to enhance selective tumor cell replication without much regard to interactions with the immune system. Adenovirus possesses a wide range of viral factors to attenuate both innate anti-viral pathways and immune cell killing. In this review, we summarize the strategies of oncolytic adenoviruses currently in clinical trials, and speculate how the mutational backgrounds of these viruses may impact upon the efficacy of these agents in oncolytic and immunotherapy. Despite decades of research on human adenoviruses, the interactions that these viruses have with the immune system remains one of the most understudied aspects of the virus and needs to be improved to rationally design the next generation of engineered viruses.
The first example of a virus that was genetically modified to selectively replicate in cancer cells came from a 1991 study of Herpes Simplex Virus 1(HSV-1) [1]. In the case of HSV-1 the genetic modification of virus was a deletion of the thymidine kinase gene, which made the virus dependent upon infecting dividing cells that maintain higher nucleotide pools. The development of HSV-1 as an oncolytic moved through several iterations, culminating in the development and FDA approval of Talimogene laherparepvec (TVEC) in 2015 for the treatment of melanoma. In addition to deleting viral genes involved in virus pathology and immune suppression, TVEC also included the strategy of arming the virus with a cellular gene, GM-CSF, to stimulate immunity and mount an immune response against the tumor. Whereas early research to optimize oncolytic viruses focussed on enhancing differential expression between normal and tumor cells, the current emphasis has shifted towards maximizing immune responses against tumor cells infected with the oncolytic virus. The clinical success of immune checkpoint inhibitors (ICIs) has diverted some attention away from the use of viral agents as monotherapy agents; however, the limitations of ICIs have also raised opportunities for combination therapy with oncolytic viruses. Cancer therapies with ICIs typically have a major portion of patients that fail to respond because the tumors do not have sufficient immune infiltration and are immunologically “cold”. The use of viral agents to turn immunologically “cold” tumors “hot” and improve ICI therapeutic response has raised interest in the use of oncolytic viruses [2]. The ability of recombinant viruses to be armed with transgenes expressing pro-inflammatory proteins makes them ideal tools to couple with ICI therapy. The clincicaltrials.gov website currently shows that there are 130 ongoing clinical trials using oncolytic viruses, and of these 33 are testing combinations with a viral oncolytic and ICIs.
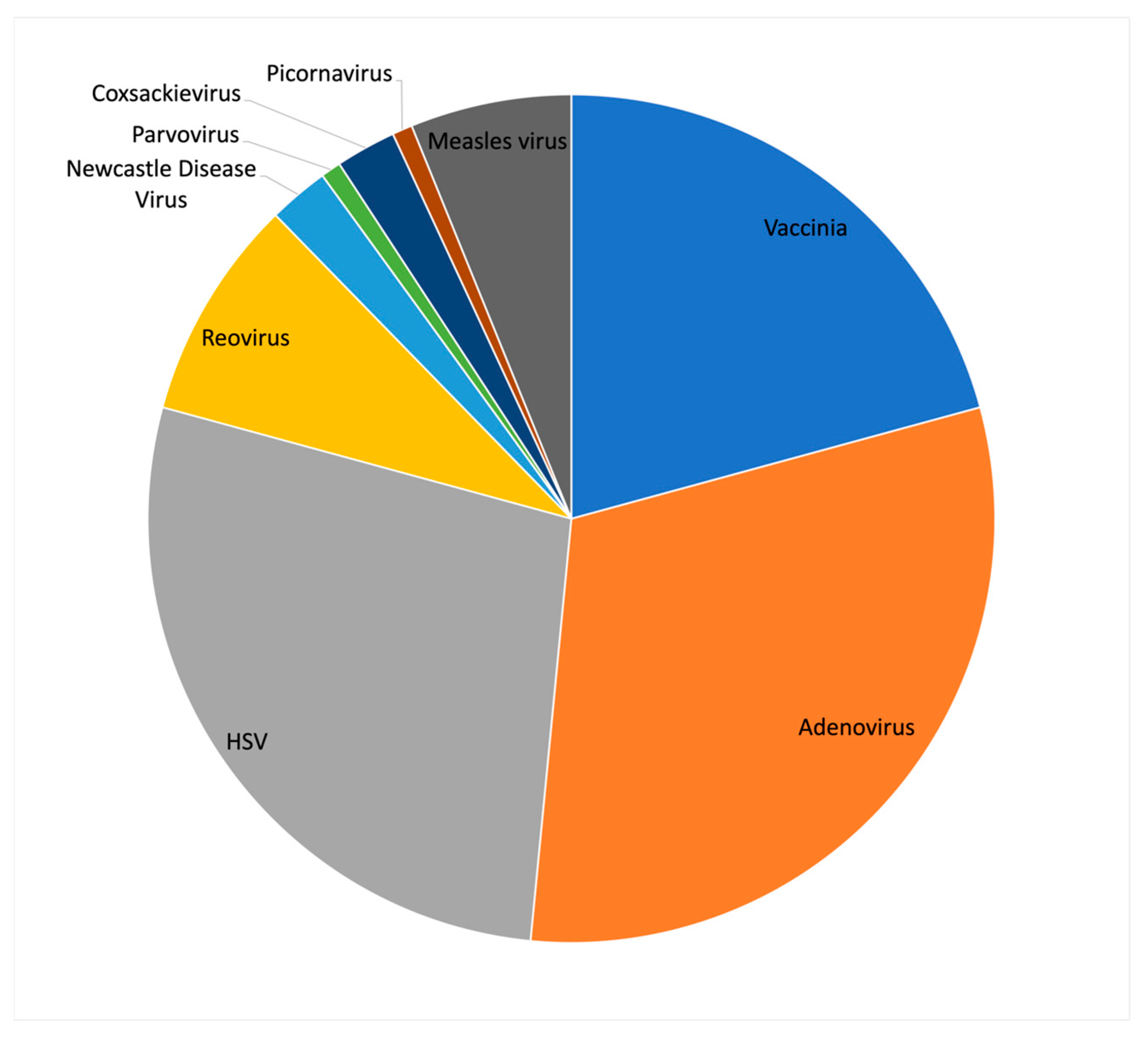
Although TVEC was the first, and currently the only, US FDA approved oncolytic virus, it was not the first to be approved in the world. The first oncolytic virus to be approved was H101 (oncorin), an adenoviral oncolytic, which was approved in 2005 by the Chinese State Food and Drug Administration for the treatment of head and neck cancers [3]. The design strategy for H101 came from previous studies from the lab of Frank McCormick, which showed that a mutant virus carrying a deletion of the E1B-55K gene in Adenovirus type 5 (Ad5), later called Onyx-015, resulted in selective viral replication in cancer cells with p53 mutations [4]. The E1B-55K gene encodes a viral protein that binds and inhibits the p53 protein to prevent virus induced apoptosis and cell cycle arrest. Since the publication of this study, the mechanism of action of the Onyx-015 virus has been disputed, and alternative mechanisms such as differences in mRNA transport have been proposed [5]. Although the clinical application of Onyx-015 was not pursued further, H101 has multiple clinical trials underway targeting a variety of cancer types. Despite the challenges posed by using adenovirus as an oncolytic agent, interest in the virus for clinical applications remains strong. Examining the 130 oncolytic virus clinical trials listed on the clincicaltrials.gov website shows that the largest fraction of these (31%) utilize Adenovirus, followed by HSV (28%) and Vaccinia (21%) (Figure 1).
There are several properties of human adenoviruses that make them ideally suited as next generation oncolytic agents used in combination with ICIs. First, the molecular virology of adenovirus has been studied for several decades, and it is one of the most well characterized viral families. Second, deriving recombinant adenovirus has become relatively straightforward. Importantly, most individuals are seropositive for Adenovirus type 5 (Ad5), the most commonly used serotype as an oncolytic agent [6]. The use of Ad5 based vectors for vaccine delivery or as gene vectors is complicated by the high seropositivity in the population; however, this is likely a beneficial property when considering Adenovirus in combination with ICIs. Since most cancer patients will be seropositive for Ad5, they will be able to rapidly mount a response against infected tumor cells. Early data using oncolytic Ad5 in combination with ICIs are beginning to show promise. At the recent American Association for Cancer Research (AACR) and ASCO 2022 meetings, the biotech company GC Oncology presented striking clinical phase I/II data using an oncolytic Ad5 (CG0070) in combination with the ICI, Keytruda (Pembrolizumab). Although the cohort consisted of only 16, patients, 14 of these achieved a complete response after only three months of treatment [7]. The development of adenovirus-based vectors as oncolytic agents has progressed through several generations and 20 different genetically engineered version of vectors have been recently used in clinical trials (Table 1). These recombinant adenoviruses are in use in 42 clinical trials (Table 2). Several modifications have been made to the viral genome in efforts to make the replication of the virus more selective to cancer cells. In some cases, oncolytic adenovirus (OAd) vectors have been armed with cellular genes to enhance immune responses. Two OAds include the cDNA encoding Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF), a similar strategy utilized in the development of TVEC. Interestingly, CG0070, which has shown promising results in the clinic, also includes a CM-CSF transgene. Other immune stimulating transgenes used include CD40 ligand, TNFα, IFNβ, OXO40 Ligand, IL-2, CXCL9, and CXCL10 (Table 1).
1. Combining Oncolytic Adenovirus with Immune Checkpoint Inhibitors
The rationale for the clinical use of OAd in combination with ICIs is supported by several preclinical studies demonstrating enhanced therapeutic effects. Using an immunohumanized glioblastoma model, an Ad vector encoding a PD-L1 antibody (XVir-N-31) was shown to induce strong immunogenic anti-tumor effects that exceed those with viral treatment alone [31]. Another study using an E1A delta24 OAd expressing a modified version of PD-1 (Ad-CAB) demonstrated excellent immune cell killing of tumor cell both with small cell lung carcinoma [32]. Combination of H101 with anti-PD-1 therapy was shown to enhance anti-tumor response in murine colorectal cancer [33].
Interestingly, even OAd that have already been armed with immune stimulating cytokines can be enhanced even further in combination with ICIs. TILT-123, for example, is armed with TNFa and IL-2 (Table 1) and has been used in combination with PD-1 inhibitors to treat ovarian and head and neck cancers in mouse [34,35]. This same combination was also shown to effectively prevent growth and metastasis of murine melanoma and colorectal cancer [36]. Impressively, metastasis was also inhibited, and mice were rendered resistant to subsequent challenge with injected tumor cells. An OAd armed with GM-CSF was also shown to be greatly enhanced in combination with anti-cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) or anti-PD1 antibodies for the treatment of triple negative breast cancers [37]. Similarly, an OAd expressing GM-CSF, IL12, and relaxin was enhanced by combination with a PD-1 inhibitor in a hamster pancreatic cancer model [38]. The molecular virology of Adenovirus has been studied for 50 years and is well understood; however, the question of which viral genes should be mutated to enhance the oncolytic properties is still an open question. Like many viruses, Adenoviruses also possess an array of viral factors that target immune responses to infected cells and the innate cellular anti-viral pathways such as interferon (IFN) signaling. If Ad-based oncolytic viruses are to be used in combination with ICIs, the mutations introduced in the viral genome need to be re-evaluated for this purpose. Existing laboratory strains of Ad5 commonly contain mutations in the E3 transcription unit of the virus. E3 mutations were often used in the early days of adenovirus research to facilitate molecular cloning of the genome, and since this transcription unit appeared to be dispensable for efficient viral replication in vitro. Subsequent research has shown that the genes encoded by E3 are a multi-functional immune inhibitory cassette that is essential for viral evasion of immune surveillance. To enhance immune stimulating effects of adenovirus, mutating some the E3 functions would certainly be beneficial. In addition to E3, the E1A protein is also known to mediate suppressive effects on IFN-dependent gene expression [39,40,41]. Similarly, adenovirus expresses non-coding RNAs (VA-RNAs), that also attenuates innate anti-viral responses [42,43,44]. Future iterations of OAd should be optimised for selective replication in cancer cells while enhancing pro-inflammatory effects around the tumor. In the sections below we summarize the genetic modifications that are included in OAd vectors currently in clinical trials and provide an assessment of how these mutations can potentially affect the properties of the virus as a mono therapy or in combination with ICIs. Beyond existing OAd vectors, we speculate on other modifications that may enhance tumor selective replication and/or increase anti-tumor immunity.
2. Modifications in the E1A Gene
The E1A transcription unit is the first to be expressed during adenovirus infection. The protein encoded on E1A is a critical transcription factor required to drive early viral gene expression and is absolutely required for viral replication. E1A is also oncogenic, inducing cell cycle progression and expression of S-phase genes in preparation for viral DNA replication. Different strategies have been implemented to render replication of the OAds more selective to tumor cells rather than normal cells. One of these was to make E1A ineffective in non-dividing cells. E1A contains two conserved regions (CR1 and CR2) that are required to bind to the tumor suppressor protein Rb to release its interaction from the cellular transcription factors E2Fs. The E2Fs are required for activation of the adenovirus E2 region coding for the viral replication machinery and to enhance expression of cellular S-phase genes. Fueyo et al. [45] deleted the CR2 region (this mutant is now called the delta 24 mutation (deleting 8 aa)), reasoning that a virus with this mutation would kill target tumor cells while spreading the infection within the tumor but not the non-dividing cells surrounding the tumor. This mutation has been frequently adopted in combination with other alterations. Of the 17 different oncolytic viruses described in this study that were not deleted for the entire E1 region, 9 contain this deletion.
Another strategy is to modify the promoter of the E1A gene to make it selectively active in cancer cells. One such strategy was to include E2F regulatory elements in the E1A promoter, which makes the E1A promoter more selective to cells that are already dividing [46]. Another was to make a deletion of 50 bp in the E1A promoter, which resulted in a marked reduction in the expression of both E1A and E1B in growth arrested cells but not in tumor cells [15]. In other cases, the promoter was replaced with that of the telomerase gene [17] or the survivin gene [29].
As mentioned above, E1A protein is also known to mediate suppressive effects on IFN-dependent gene expression [39,40,41]. Although none of the virus mutants in the clinical trials have use the strategy to mutate E1A in a way for it not to interfere with IFN gene expression, it is nonetheless a strategy that could be taken in future studies. The major mechanism for this effect appears to be that E1A represses the transcription of interferon-stimulated genes (ISG) by decreasing the levels of ISRE- binding factors, an effect dependent on the CR1 region [39,40,41]. However, since this region is critical for activating E2F, such mutations may also come with a cost of attenuated viral replication. There are, however, other mechanisms used by E1A. The N-terminal region of E1A (before the CR1 motif) is involved in the binding to hBre1 complex to block IFN-induced H2B mono-ubiquitination that is required for ISG expression [47,48]. The N-terminal region (as well as the CR3) is also involved in binding a component of the immunoproteasome, MECL1 [49]. It is thought that this interaction could result in reduced antigen presentation by the MHCI system [49]. The C-terminal of E1A has been known for some time to bind three proteins, FOXK, DCAF7 and CtBP. More recently, Zemke and Berk have shown that these interactions with E1A result in the downregulation of a subset of ISGs [50]. It may therefore be of interest to determine if mutations in these regions of E1A improve the immunogenicity of OAd vectors.
3. Modification of the E1B Gene
The E1B genes have also been modified in some OAds. This transcription unit encodes two different proteins, E1B-55K and E1B-19K, on overlapping reading frames. The E1B-55K was the gene mutated in the first OAd Onyx-015 (see above) as it is required for the inactivation of the p53 tumor suppressor. Two of the viruses that are undergoing clinical trials, Oncorin and MEM-288, have mutations that eliminate expression of E1B-55K. In addition to inactivation of p53, E1B-55K has other viral functions including viral mRNA transport and shut-off of host cell mRNA translation [51]. Viral mutants of E1B-55K do appear to be selectively oncolytic, although the mechanism of this effect is likely more complex than originally hypothesized [52,53].
The E1B-19K gene encodes a functional homolog of the cellular Bcl-2 protein and functions as a suppressor of apoptosis during viral replication [54]. Viral mutants that do not express E1B-19K are more lytic and have much larger plaque sizes when grown in vitro [55]. The E1B-19K gene has been deleted in one vector currently in clinical trials (AdAPT-001). It appears that in this case the E1B-19K gene was removed to insert the transgene in its place, and it is unclear if it was intended to also improve the oncolytic functions of the vector [16]. The impact of E1B-19K on the efficacy of adenovirus as an oncolytic has never been directly addressed in the literature but should be considered. One study has shown that E1B-19K represses pro-inflammatory signaling during viral replication [56]. The corpses of cells killed by virus lacking expression of E1B-19K elicited stronger pro-inflammatory cytokine production when presented to macrophages. This observation suggests that deletion of E1B-19K would be beneficial in the design of an OAd to enhance immune infiltration of tumors and may also enhance effects in co-operation with ICIs.
4. Modification of the Fibre Protein
Although most modifications to the adenovirus genome for oncolytic purposes have been within non-structural proteins, there have also been some efforts to alter the tropism of the virus by modifying capsid proteins and making infection more selective to tumor cells. The fiber protein has a critical role for the entry of adenoviruses in cells. With Ad5-based viruses the knob domain of the fiber protein first establishes a high affinity interaction with the cellular Coxsackie Adenovirus Receptor (CAR) [57], followed by an internalization induced via the binding of the RGD motif in the penton base to the alphaV integrins [58]. While the CAR is widely expressed in most cell types, it is poorly expressed in cells of hematopoietic origins [59] and is frequently lost in tumors [60,61,62,63,64,65]. The group B Adenoviruses, of which Ad3, 11 and 35 are members, utilize the CD46 and DSG-2 as receptors [66,67,68,69]. Both CD46 and DSG-2 [70] have been shown to be overexpressed in several tumors [71,72,73,74,75,76,77,78,79]. Thus, a strategy often used to ensure efficient infection of the oncolytic viruses in the tumor cells has been to modify the fiber gene. One approach has been to use a group B virus as backbone to generate the oncolytic viruses. Three viruses currently in clinical trials are based on a chimera of Ad11 and Ad3 with the fiber gene contributed by the Ad11 virus (see Table 1). Another approach has been to modify the fiber gene of the Ad5 serotype to contain the fiber shaft and knob domain of Ad35 [80], or the knob domain of Ad3 [22]. Another frequently used approach is to improve the entry of Ad5-based oncolytic viruses by cloning the RGD motif normally present in the Penton protein to the HI-loop of the fiber protein [81,82]. This modification reduces dependence on the CAR receptor by allowing the virus to utilize the RGD-integrin as an alternative pathway. Adding a polylysine (pK7) peptide to the C terminus of the RGD-fiber has been shown to further improve CAR-independent viral entry through the ability of the pK7 to bind polyanion motifs such as heparan sulfate, which are ubiquitously expressed on the surface of most cell types [83].
5. Modifications of the E3 Region
The E3 region of adenovirus is the least studied region of the virus, mostly because it was quickly found to be dispensable for virus replication in cultured cells [84]. Indeed, many early studies of adenoviruses were performed using the dl309 as a wild-type virus in which a major part of the E3 region was deleted as it was easier to make viral mutations in this background [85]. Most of the oncolytic viruses currently in clinical trials are of the Ad5 background, and the viruses of the Ad3/11 hybrid background has a deletion removing almost the entirety of the E3 region. As such, this section will focus on the E3 region of the Ad5 (group C) serotype. The E3 region of Ad5 expresses seven proteins that are thought to be immunomodulatory in function (reviewed in [86]). These properties make the E3 region of particular interest in the study of oncolytic viruses, considering that involvement of an immune response is beneficial for the success of oncolytic therapy. However, most of the oncolytic viruses currently in use have complete or partial deletions of the E3 region to facilitate the inclusion of transgenes. It remains to be determined if these E3 mutations are beneficial or detrimental to oncolytic or immunotherapy.
The proteins encoded on the E3 region have a wide range of effects on immune functions that can potentially enhance or inhibit oncoloytic activity (for an in-depth review, see [86]). The proteins encoded on E3 include: 12.5K, RIDa, RIDb, 6.7K, 14.7K, GP19K, and ADP. Remarkably, despite decades of research on Ad5, the functions of the E3-12.5K remain completely unknown and therefore its impact on oncolytic therapy cannot even be speculated without further investigation. The Receptor Internalization and Degradation proteins (RIDα and RIDβ) function as a heterodimer to block apoptosis induced by a variety of death ligands [87,88,89,90,91,92]. The RID complex also blocks TNF-mediated death [93,94] and may block EGFR-mediated inflammatory responses [95]. RID has also been shown to inhibit the interleukin 1- and TNF-induced NF-kB activation. The E3-6.7K is known as a general inhibitor of apoptosis, blocking both intrinsic and extrinsic pathways [96]. The E3-14.7K can protect cells from extrinsic apoptosis induced by some cytokines [86] and is a general inhibitor of TNF-mediated apoptosis [97,98]. E3-14.7K may also affect TNF-mediated inflammation as it inhibits TNF-induced activation of cPLA2 and thus the release of pro-inflammatory molecules [94,99]. The GP19K protein was shown to reduce the CTL-mediated killing of infected cells by blocking the transport of MHC class I molecules to the cell surface [100,101,102,103,104,105]. It does this, in part, by binding to and sequestering MHC class 1 molecules to the ER [101,102,104,106]. The final product of this region, the adenovirus death protein (ADP), is regulated by both the E3 promoter and the major late promoter and therefore reaches maximal expression in the late phase of infection. ADP kills infected cells by an unknown mechanism involving membrane degradation that is not apoptosis [107]. It is thought that this function of ADP helps with cell lysis and viral spread at the end of the infection cycle.
From the known functions of these E3 proteins it is possible to speculate which would be beneficial to leave in the oncolytic viruses and which could be deleted to make space for transgenes. To enhance viral spread and killing activity it would seem that expression of the ADP through the major late promoter should be maintained in oncolytic Ad vectors. Mutants of Ad that lack expression of ADP do not easily exit the cell, and methods such as freeze/thaw cycles need to be used to promote cell lysis and viral egress [84]. However, a recent study has shown that in some specific cases it may be better to remove ADP to achieve the best expression of transgene [108]. In this study the authors use viral vectors in which all E3 genes are removed except ADP. This results in increase expression of ADP and increased lysis of infected cells [109]. In addition, as their transgene (sodium iodide symporter, NIS) is a transmembrane protein they found that removal of the ADP gene resulted in better expression of NIS, and thus better imaging of infected tumor cells [108].
Conversely, since the RID complex and E3-14-7K appears to block inflammation it is likely that deleting these genes would be beneficial to increase immune killing of tumor cells. Similarly, since GP19K downregulates MHC complexes, deleting this gene would be essential to maintain immune cell killing of infected cancer cells. It is unfortunate that E3 remains one of the less studied transcription units of Ad since it will likely play a critical role in viral and immune system-mediated killing by oncolytic adenoviral vectors. Understanding the function of the E3-12.5K and its impact on tumor cell killing should also be prioritized since virtually nothing is currently known about this protein.
6. VA-RNAs
Adenoviruses produce two small, non-coding RNAs transcribed by RNA polymerase III, called VA-RNA I and VA-RNA II, that are approximately 160 nucleotide long [110]. They form a complex RNA structure that is stable and conserved, even if the actual sequence is not [111,112]. The structure is critical for binding and inhibiting an antiviral response protein, PKR [42,43,44]. These RNAs have been shown to be required for efficient viral replication [44,113,114,115] and the mechanism is at least in part due to its inhibition of PKR [43]. More recently, VA-RNAs were shown to activate the promoter of type 1 IFNs [116,117,118] sufficiently to induce expression of some IFN-induced genes; however, this does not happen by the classical pathway involving IRF-3 [117]. Although none of the currently used OAd vectors were designed with the status of VA-RNAs in mind, it may be a strategy to consider in the future. Whether to retain or remove the VA-RNAs is complicated by the still controversial role of PRK in cancer. Traditionally PRK has been considered a tumor suppressor since the mRNA is often downregulated in several cancer types (reviewed in [119]). In these tumors, deletion of the VA-RNAs may act as another factor to increase tumor selectivity. However, other studies have suggested an opposite effect of PRK and it has been shown to be overexpressed in some tumors [120,121,122,123]. The case for deletion of VA-RNAs in oncolytic Ad vectors may therefore depend upon the cancer type being targeted and the expression of PKR in these cell types.
7. Conclusions
The application of adenovirus as an oncolytic viral therapy is entering an exciting phase. The combination of these vectors with ICIs is generating provocative results in the clinic and may provide a solution to improve response rates to immunotherapy. However, most of the existing Ad vectors have not been optimized to remove viral genes that attenuate immune and innate anti-viral responses. With the wide range of ongoing clinical trials using Ad vectors, a clearer view may begin to emerge regarding which alterations of viral genes are best to enhance oncolytic properties of the virus and to enhance anti-tumor immune responses. In particular, further research into the E3 transcription unit and its role in suppressing immune cell killing may provide important insights in understanding the interactions of Ad-infected cells with the immune system, and how these effects influence oncolytic virotherapy and immunotherapy.
Author Contributions
Conceptualization and writing contributed equally by P.B. and J.G.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funding from the Canadian Institutes of Health Research (CIHR), Natural Sciences and Engineering Research Council (NSERC) and the Fonds de Recherche du Québec Nature et Technologies (FRQNT) to J.G.T.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Martuza, R.L.; Malick, A.; Markert, J.M.; Ruffner, K.L.; Coen, D.M. Experimental therapy of human glioma by means of a genetically engineered virus mutant. Science 1991, 252, 854–856. [Google Scholar] [CrossRef]
- Twumasi-Boateng, K.; Pettigrew, J.L.; Kwok, Y.Y.E.; Bell, J.C.; Nelson, B.H. Oncolytic viruses as engineering platforms for combination immunotherapy. Nat. Rev. Cancer 2018, 18, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. China Approves World’s First Oncolytic Virus Therapy For Cancer Treatment. J. Natl. Cancer Inst. 2006, 98, 298–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, J.R.; Kirn, D.H.; Williams, A.; Heise, C.; Horn, S.; Muna, M.; Ng, L.; Nye, J.A.; Sampson-Johannes, A.; Fattaey, A.; et al. An Adenovirus Mutant That Replicates Selectively in p53- Deficient Human Tumor Cells. Science 1996, 274, 373–376. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, C.C.; Johnson, L.; Bagus, B.; Choi, S.; Nicholas, C.; Shen, A.; Boyle, L.; Pandey, K.; Soria, C.; Kunich, J.; et al. Late viral RNA export, rather than p53 inactivation, determines ONYX-015 tumor selectivity. Cancer Cell 2004, 6, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Mast, T.C.; Kierstead, L.; Gupta, S.B.; Nikas, A.A.; Kallas, E.G.; Novitsky, V.; Mbewe, B.; Pitisuttithum, P.; Schechter, M.; Vardas, E.; et al. International epidemiology of human pre-existing adenovirus (Ad) type-5, type-6, type-26 and type-36 neutralizing antibodies: Correlates of high Ad5 titers and implications for potential HIV vaccine trials. Vaccine 2010, 28, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Steinberg, G.D.; Uchio, E.M.; Lamm, D.L.; Shah, P.; Kamat, A.M.; Bivalacqua, T.; Packiam, V.T.; Chisamore, M.J.; McAdory, J.; et al. CORE1: Phase 2, single-arm study of CG0070 combined with pembrolizumab in patients with nonmuscle-invasive bladder cancer (NMIBC) unresponsive to bacillus Calmette-Guerin (BCG). J. Clin. Oncol. 2022, 40, 4597. [Google Scholar] [CrossRef]
- Lei, J.; Li, Q.H.; Yang, J.L.; Liu, F.; Wang, L.; Xu, W.M.; Zhao, W.X. The antitumor effects of oncolytic adenovirus H101 against lung cancer. Int. J. Oncol. 2015, 47, 555–562. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.H.; Shine, H.D.; Goodman, J.C.; Grossman, R.G.; Woo, S.L. Gene therapy for brain tumors: Regression of experimental gliomas by adenovirus-mediated gene transfer in vivo. Proc. Natl. Acad. Sci. USA 1994, 91, 3054–3057. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Dai, W.; Antonia, S.; Cantwell, M.J.; Beg, A.A. Abstract 4578: Development of MEM-288, a Dual-Transgene Armed and Conditionally Replication-Enhanced Oncolytic Adenovirus with Potent Systemic Antitumor Immunity. 2020. Available online: https://aacrjournals.org/cancerres/article/80/16_Supplement/4578/643321/Abstract-4578-Development-of-MEM-288-a-dual (accessed on 20 November 2022).
- Eriksson, E.; Milenova, I.; Wenthe, J.; Stahle, M.; Leja-Jarblad, J.; Ullenhag, G.; Dimberg, A.; Moreno, R.; Alemany, R.; Loskog, A. Shaping the Tumor Stroma and Sparking Immune Activation by CD40 and 4-1BB Signaling Induced by an Armed Oncolytic Virus. Clin. Cancer Res. 2017, 23, 5846–5857. [Google Scholar] [CrossRef]
- Eriksson, E.; Moreno, R.; Milenova, I.; Liljenfeldt, L.; Dieterich, L.C.; Christiansson, L.; Karlsson, H.; Ullenhag, G.; Mangsbo, S.M.; Dimberg, A.; et al. Activation of myeloid and endothelial cells by CD40L gene therapy supports T-cell expansion and migration into the tumor microenvironment. Gene Ther 2017, 24, 92–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenthe, J.; Eriksson, E.; Milenova, I.; Moreno, R.; Alemany, R.; Loskog, A. 516. A Novel Oncolytic Adenovirus Expressing Tumor Microenvironment Stimulators to Evoke and Facilitate Anti-Tumor Immune Responses. Cancer-Oncolytic DNA Viruses 2016, 24 (Suppl. 1), S206. [Google Scholar] [CrossRef]
- Burke, J.M.; Lamm, D.L.; Meng, M.V.; Nemunaitis, J.J.; Stephenson, J.J.; Arseneau, J.C.; Aimi, J.; Lerner, S.; Yeung, A.W.; Kazarian, T.; et al. A first in human phase 1 study of CG0070, a GM-CSF expressing oncolytic adenovirus, for the treatment of nonmuscle invasive bladder cancer. J. Urol. 2012, 188, 2391–2397. [Google Scholar] [CrossRef] [PubMed]
- Hedjran, F.; Shantanu, K.; Tony, R. Deletion analysis of Ad5 E1a transcriptional control region: Impact on tumor-selective expression of E1a and E1b. Cancer Gene Ther. 2011, 18, 717–723. [Google Scholar] [CrossRef] [Green Version]
- Larson, C.; Oronsky, B.; Abrouk, N.E.; Oronsky, A.; Reid, T.R. Toxicology and biodistribution of AdAPT-001, a replication-competent type 5 adenovirus with a trap for the immunosuppressive cytokine, TGF-beta. Am. J. Cancer Res. 2021, 11, 5184–5189. [Google Scholar]
- Kawashima, T.; Kagawa, S.; Kobayashi, N.; Shirakiya, Y.; Umeoka, T.; Teraishi, F.; Taki, M.; Kyo, S.; Tanaka, N.; Fujiwara, T. Telomerase-specific replication-selective virotherapy for human cancer. Clin. Cancer Res. 2004, 10, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamasaki, Y.; Tazawa, H.; Hashimoto, Y.; Kojima, T.; Kuroda, S.; Yano, S.; Yoshida, R.; Uno, F.; Mizuguchi, H.; Ohtsuru, A.; et al. A novel apoptotic mechanism of genetically engineered adenovirus-mediated tumour-specific p53 overexpression through E1A-dependent p21 and MDM2 suppression. Eur. J. Cancer 2012, 48, 2282–2291. [Google Scholar] [CrossRef] [Green Version]
- Kuhn, I.; Harden, P.; Bauzon, M.; Chartier, C.; Nye, J.; Thorne, S.; Reid, T.; Ni, S.; Lieber, A.; Fisher, K.; et al. Directed evolution generates a novel oncolytic virus for the treatment of colon cancer. PLoS ONE 2008, 3, e2409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Aiguirre, I.; Cassady, K.A. Chapter Twelve - Oncolytic viruses in immunotherapy. In Cancer Immunology and Immunotherapy; Amiji, M.M., Scheherazade Milane, L., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 375–437. [Google Scholar]
- Lang, F.F.; Conrad, C.; Gomez-Manzano, C.; Yung, W.K.A.; Sawaya, R.; Weinberg, J.S.; Prabhu, S.S.; Rao, G.; Fuller, G.N.; Aldape, K.D.; et al. Phase I Study of DNX-2401 (Delta-24-RGD) Oncolytic Adenovirus: Replication and Immunotherapeutic Effects in Recurrent Malignant Glioma. J. Clin. Oncol. 2018, 36, 1419–1427. [Google Scholar] [CrossRef]
- Kanerva, A.; Zinn, K.R.; Chaudhuri, T.R.; Lam, J.T.; Suzuki, K.; Uil, T.G.; Hakkarainen, T.; Bauerschmitz, G.J.; Wang, M.; Liu, B.; et al. Enhanced therapeutic efficacy for ovarian cancer with a serotype 3 receptor-targeted oncolytic adenovirus. Mol. Ther. 2003, 8, 449–458. [Google Scholar] [CrossRef]
- Koski, A.; Kangasniemi, L.; Escutenaire, S.; Pesonen, S.; Cerullo, V.; Diaconu, I.; Nokisalmi, P.; Raki, M.; Rajecki, M.; Guse, K.; et al. Treatment of cancer patients with a serotype 5/3 chimeric oncolytic adenovirus expressing GMCSF. Mol. Ther. 2010, 18, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Siurala, M.; Bramante, S.; Vassilev, L.; Hirvinen, M.; Parviainen, S.; Tahtinen, S.; Guse, K.; Cerullo, V.; Kanerva, A.; Kipar, A.; et al. Oncolytic adenovirus and doxorubicin-based chemotherapy results in synergistic antitumor activity against soft-tissue sarcoma. Int. J. Cancer 2015, 136, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Havunen, R.; Kalliokoski, R.; Siurala, M.; Sorsa, S.; Santos, J.M.; Cervera-Carrascon, V.; Anttila, M.; Hemminki, A. Cytokine-Coding Oncolytic Adenovirus TILT-123 Is Safe, Selective, and Effective as a Single Agent and in Combination with Immune Checkpoint Inhibitor Anti-PD-1. Cells 2021, 10. [Google Scholar] [CrossRef]
- Atherton, M.J.; Stephenson, K.B.; Nikota, J.K.; Hu, Q.N.; Nguyen, A.; Wan, Y.; Lichty, B.D. Preclinical development of peptide vaccination combined with oncolytic MG1-E6E7 for HPV-associated cancer. Vaccine 2018, 36, 2181–2192. [Google Scholar] [CrossRef]
- Jonker, D.J.; Hotte, S.J.; Razak, A.R.A.; Renouf, D.J.; Lichty, B.; Bell, J.C.; Powers, J.; Breitbach, C.J.; Stojdl, D.F.; Stephenson, K.B.; et al. Phase I study of oncolytic virus (OV) MG1 maraba/MAGE-A3 (MG1MA3), with and without transgenic MAGE-A3 adenovirus vaccine (AdMA3) in incurable advanced/metastatic MAGE-A3-expressing solid tumours: CCTG IND.214. J. Clin. Oncol. 2017, 35, e14637. [Google Scholar] [CrossRef]
- Farrera-Sal, M.; Moreno, R.; Mato-Berciano, A.; Maliandi, M.V.; Bazan-Peregrino, M.; Alemany, R. Hyaluronidase expression within tumors increases virotherapy efficacy and T cell accumulation. Mol. Ther. Oncolytics. 2021, 22, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Ulasov, I.V.; Zhu, Z.B.; Tyler, M.A.; Han, Y.; Rivera, A.A.; Khramtsov, A.; Curiel, D.T.; Lesniak, M.S. Survivin-driven and fiber-modified oncolytic adenovirus exhibits potent antitumor activity in established intracranial glioma. Hum. Gene Ther. 2007, 18, 589–602. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; van Ginkel, J.W.; Au, K.Y.; Alemany, R.; Meulenberg, J.J.; van Beusechem, V.W. ORCA-010, a novel potency-enhanced oncolytic adenovirus, exerts strong antitumor activity in preclinical models. Hum. Gene Ther. 2014, 25, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Klawitter, M.; El-Ayoubi, A.; Buch, J.; Rüttinger, J.; Ehrenfeld, M.; Lichtenegger, E.; Krüger, M.A.; Mantwill, K.; Koll, F.J.; Kowarik, M.C.; et al. The Oncolytic Adenovirus XVir-N-31, in Combination with the Blockade of the PD-1/PD-L1 Axis, Conveys Abscopal Effects in a Humanized Glioblastoma Mouse Model. Int. J. Mol. Sci. 2022, 23, 9965. [Google Scholar] [CrossRef]
- Zhao, Q.; Tong, J.; Liu, X.; Li, S.; Chen, D.; Miao, L. Reversing resistance to immune checkpoint inhibitor by adding recombinant human adenovirus type 5 in a patient with small cell lung cancer with promoted immune infiltration: A case report. J. Cancer Res. Clin. Oncol. 2022, 148, 1269–1273. [Google Scholar] [CrossRef]
- Huang, L.; Zhao, H.; Shan, M.; Chen, H.; Xu, B.; He, Y.; Zhao, Y.; Liu, Z.; Chen, J.; Xu, Q. Oncolytic adenovirus H101 ameliorate the efficacy of anti-PD -1 monotherapy in colorectal cancer. Cancer Med. 2022, 11, 4575–4587. [Google Scholar] [CrossRef] [PubMed]
- Heiniö, C.; Clubb, J.; Kudling, T.; Quixabeira, D.; Cervera-Carrascon, V.; Havunen, R.; Grönberg-Vähä-Koskela, S.; Santos, J.M.; Tapper, J.; Kanerva, A.; et al. Effective Combination Immunotherapy with Oncolytic Adenovirus and Anti-PD-1 for Treatment of Human and Murine Ovarian Cancers. Diseases 2022, 10, 52. [Google Scholar] [CrossRef] [PubMed]
- Clubb, J.H.A.; Kudling, T.V.; Heiniö, C.; Basnet, S.; Pakola, S.; Cervera Carrascón, V.; Santos, J.M.; Quixabeira, D.C.A.; Havunen, R.; Sorsa, S.; et al. Adenovirus Encoding Tumor Necrosis Factor Alpha and Interleukin 2 Induces a Tertiary Lymphoid Structure Signature in Immune Checkpoint Inhibitor Refractory Head and Neck Cancer. Front. Immunol. 2022, 13, 794251. [Google Scholar] [CrossRef]
- Quixabeira, D.C.A.; Cervera-Carrascon, V.; Santos, J.M.; Clubb, J.H.; Kudling, T.V.; Basnet, S.; Heiniö, C.; Grönberg-Vähä-Koskela, S.; Anttila, M.; Havunen, R.; et al. Local therapy with an engineered oncolytic adenovirus enables antitumor response in non-injected melanoma tumors in mice treated with aPD-1. Oncoimmunology 2022, 11, 2028960. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xie, W.; Zhang, Y.; Dong, X.; Liu, C.; Yi, J.; Zhang, S.; Wen, C.; Zheng, L.; Wang, H. Oncolytic adenoviruses synergistically enhance anti-PD-L1 and anti-CTLA-4 immunotherapy by modulating the tumour microenvironment in a 4T1 orthotopic mouse model. Cancer Gene Ther. 2022, 29, 456–465. [Google Scholar] [CrossRef]
- Jung, B.K.; Ko, H.Y.; Kang, H.; Hong, J.; Ahn, H.M.; Na, Y.; Kim, H.; Kim, J.S.; Yun, C.O. Relaxin-expressing oncolytic adenovirus induces remodeling of physical and immunological aspects of cold tumor to potentiate PD-1 blockade. J. Immunother. Cancer 2020, 8, e000763. [Google Scholar] [CrossRef]
- Ackrill, A.M.; Foster, G.R.; Laxton, C.D.; Flavell, D.M.; Stark, G.R.; Kerr, I.M. Inhibition of the cellular response to interferons by products of the adenovirus type 5 E1A oncogene. Nucleic Acids Res. 1991, 19, 4387–4393. [Google Scholar] [CrossRef] [Green Version]
- Gutch, M.J.; Reich, N.C. Repression of the interferon signal transduction pathway by the adenovirus E1A oncogene. Proc. Natl. Acad. Sci. USA 1991, 88, 7913–7917. [Google Scholar] [CrossRef] [Green Version]
- Kalvakolanu, D.V.; Bandyopadhyay, S.K.; Harter, M.L.; Sen, G.C. Inhibition of interferon-inducible gene expression by adenovirus E1A proteins: Block in transcriptional complex formation. Proc. Natl. Acad. Sci. USA 1991, 88, 7459–7463. [Google Scholar] [CrossRef] [Green Version]
- Aparicio, O.; Razquin, N.; Zaratiegui, M.; Narvaiza, I.; Fortes, P. Adenovirus Virus-Associated RNA Is Processed to Functional Interfering RNAs Involved in Virus Production. J. Virol. 2006, 80, 1376–1384. [Google Scholar] [CrossRef] [Green Version]
- Kitajewski, J.; Schneider, R.J.; Safer, B.; Munemitsu, S.M.; Samuel, C.E.; Thimmappaya, B.; Shenk, T. Adenovirus VAI RNA antagonizes the antiviral action of interferon by preventing activation of the interferon-induced eIF-2α kinase. Cell 1986, 45, 195–200. [Google Scholar] [CrossRef]
- O’Malley, R.P.; Mariano, T.M.; Siekierka, J.; Mathews, M.B. A mechanism for the control of protein synthesis by adenovirus VA RNAI. Cell 1986, 44, 391–400. [Google Scholar] [CrossRef]
- Fueyo, J.; Gomez-Manzano, C.; Alemany, R.; Lee, P.S.; McDonnell, T.J.; Mitlianga, P.; Shi, Y.-X.; A Levin, V.; Yung, W.K.A.; Kyritsis, A.P. A mutant oncolytic adenovirus targeting the Rb pathway produces anti-glioma effect in vivo. Oncogene 2000, 19, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Xu, Y.; Zhao, Y.; Li, L.; Sun, P.; Liu, H.; Fan, Q.; Liang, K.; Liang, W.; Sun, H.; et al. Combination of E2F-1 promoter-regulated oncolytic adenovirus and cytokine-induced killer cells enhances the antitumor effects in an orthotopic rectal cancer model. Tumor Biol. 2014, 35, 1113–1122. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, G.J.; Cohen, M.J.; Nichols, A.C.; Barrett, J.W.; Mymryk, J.S. Viral Retasking of hBre1/RNF20 to Recruit hPaf1 for Transcriptional Activation. PLoS Pathog. 2013, 9, e1003411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, G.J.; Thillainadesan, G.; Yousef, A.F.; Ablack, J.N.; Mossman, K.L.; Torchia, J.; Mymryk, J.S. Adenovirus Evasion of Interferon-Mediated Innate Immunity by Direct Antagonism of a Cellular Histone Posttranslational Modification. Cell Host Microbe 2012, 11, 597–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berhane, S.; Aresté, C.; Ablack, J.N.; Ryan, G.B.; Blackbourn, D.J.; Mymryk, J.S.; Turnell, A.S.; Steele, J.C.; Grand, R.J. Adenovirus E1A interacts directly with, and regulates the level of expression of, the immunoproteasome component MECL1. Virology 2011, 421, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Zemke, N.R.; Berk, A.J. The Adenovirus E1A C Terminus Suppresses a Delayed Antiviral Response and Modulates RAS Signaling. Cell Host Microbe 2017, 22, 789–800. [Google Scholar] [CrossRef] [Green Version]
- Hidalgo, P.; Ip, W.H.; Dobner, T.; Gonzalez, R.A. The biology of the adenovirus E1B 55K protein. FEBS Lett. 2019, 593, 3504–3517. [Google Scholar] [CrossRef]
- Dix, B.R.; Edwards, S.J.; Braithwaite, A.W. Does the Antitumor Adenovirus ONYX-015/dl1520 Selectively Target Cells Defective in the p53 Pathway? J. Virol. 2001, 75, 5443–5447. [Google Scholar] [CrossRef] [Green Version]
- McCormick, F. Cancer-specific viruses and the development of ONYX-015. Cancer Biol. Ther. 2003, 2, S157–S160. [Google Scholar] [CrossRef] [PubMed]
- Degenhardt, K.; Perez, D.; White, E. Pathways used by adenovirus E1B 19K to inhibit apoptosis. Symp. Soc. Exp. Biol. 2000, 52, 241–251. [Google Scholar]
- White, E.; Grodzicker, T.; Stillman, B.W. Mutations in the gene encoding the adenovirus early region 1B 19,000-molecular-weight tumor antigen cause the degradation of chromosomal DNA. J. Virol. 1984, 52, 410–419. [Google Scholar] [CrossRef] [Green Version]
- Radke, J.R.; Grigera, F.; Ucker, D.S.; Cook, J.L. Adenovirus E1B 19-Kilodalton Protein Modulates Innate Immunity through Apoptotic Mimicry. J. Virol. 2014, 88, 2658–2669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a Common Receptor for Coxsackie B Viruses and Adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef] [PubMed]
- Wickham, T.J.; Mathias, P.; Cheresh, D.A.; Nemerow, G.R. Integrins αvβ3 and αvβ5 promote adenovirus internalization but not virus attachment. Cell 1993, 73, 309–319. [Google Scholar] [CrossRef]
- Rebel, V.I.; Hartnett, S.; Denham, J.; Chan, M.; Finberg, R.; Sieff, C.A. Maturation and Lineage-Specific Expression of the Coxsackie and Adenovirus Receptor in Hematopoietic Cells. STEM CELLS 2000, 18, 176–182. [Google Scholar] [CrossRef]
- Korn, W.M.; Macal, M.; Christian, C.; Lacher, M.D.; McMillan, A.; Rauen, K.A.; Warren, R.S.; Ferrell, L. Expression of the coxsackievirus- and adenovirus receptor in gastrointestinal cancer correlates with tumor differentiation. Cancer Gene Ther. 2006, 13, 792–797. [Google Scholar] [CrossRef]
- Okegawa, T.; Pong, R.C.; Li, Y.; Bergelson, J.M.; Sagalowsky, A.I.; Hsieh, J.T. The mechanism of the growth-inhibitory effect of coxsackie and adenovirus receptor (CAR) on human bladder cancer: A functional analysis of car protein structure. Cancer Res. 2001, 61, 6592–6600. [Google Scholar]
- A Rauen, K.; Sudilovsky, D.; Le, J.L.; Chew, K.L.; Hann, B.; Weinberg, V.; Schmitt, L.D.; McCormick, F. Expression of the coxsackie adenovirus receptor in normal prostate and in primary and metastatic prostate carcinoma: Potential relevance to gene therapy. Cancer Res. 2002, 62, 3812–3818. [Google Scholar]
- Reeh, M.; Bockhorn, M.; Görgens, D.; Vieth, M.; Hoffmann, T.; Simon, R.; Izbicki, J.R.; Sauter, G.; Schumacher, U.; Anders, M. Presence of the Coxsackievirus and Adenovirus Receptor (CAR) in human neoplasms: A multitumour array analysis. Br. J. Cancer 2013, 109, 1848–1858. [Google Scholar] [CrossRef]
- Stecker, K.; Vieth, M.; Koschel, A.; Wiedenmann, B.; Röcken, C.; Anders, M. Impact of the coxsackievirus and adenovirus receptor on the adenoma–carcinoma sequence of colon cancer. Br. J. Cancer 2011, 104, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.H.; Song, L.B.; Wu, X.J.; Li, R.P.; Zeng, M.S.; Zhu, X.F.; Wan, D.S.; Liu, Q.; Zeng, Y.X.; Zhang, X.S. Proteasome inhibitor MG-132 modifies coxsackie and adenovirus receptor expression in colon cancer cell line lovo. Cell Cycle 2008, 7, 925–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Persson, B.D.; Reiter, D.M.; Marttila, M.; Mei, Y.F.; Casasnovas, J.M.; Arnberg, N.; Stehle, T. Adenovirus type 11 binding alters the conformation of its receptor CD46. Nat. Struct. Mol. Biol. 2007, 14, 164–166. [Google Scholar] [CrossRef]
- Segerman, A.; Atkinson, J.P.; Marttila, M.; Dennerquist, V.; Wadell, G.; Arnberg, N. Adenovirus Type 11 Uses CD46 as a Cellular Receptor. J. Virol. 2003, 77, 9183–9191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinh, H.V.; Lesage, G.; Chennamparampil, V.; Vollenweider, B.; Burckhardt, C.J.; Schauer, S.; Havenga, M.; Greber, U.F.; Hemmi, S. Avidity Binding of Human Adenovirus Serotypes 3 and 7 to the Membrane Cofactor CD46 Triggers Infection. J. Virol. 2012, 86, 1623–1637. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, Z.-Y.; Liu, Y.; Persson, J.; Beyer, I.; Möller, T.; Koyuncu, D.; Drescher, M.R.; Strauss, R.; Zhang, X.-B.; et al. Desmoglein 2 is a receptor for adenovirus serotypes 3, 7, 11 and 14. Nat. Med. 2010, 17, 96–104. [Google Scholar] [CrossRef]
- Elvington, M.; Liszewski, M.K.; Atkinson, J.P. CD46 and Oncologic Interactions: Friendly Fire against Cancer. Antibodies 2021, 9, 59. [Google Scholar] [CrossRef]
- Abbod, M.F.; Hamdy, F.C.; Linkens, D.A.; Catto, J.W. Predictive modeling in cancer: Where systems biology meets the stock market. Expert Rev. Anticancer. Ther. 2009, 9, 867–870. [Google Scholar] [CrossRef] [Green Version]
- Biedermann, K.; Vogelsang, H.; Becker, I.; Plaschke, S.; Siewert, J.R.; Höfler, H.; Keller, G. Desmoglein 2 is expressed abnormally rather than mutated in familial and sporadic gastric cancer. J. Pathol. 2005, 207, 199–206. [Google Scholar] [CrossRef]
- Harada, H.; Iwatsuki, K.; Ohtsuka, M.; Han, G.W.; Kaneko, F. Abnormal desmoglein expression by squamous cell carcinoma cells. Acta Derm. Venereol. 1996, 76, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Kemper, C.; Leung, M.; Stephensen, C.B.; Pinkert, C.A.; Liszewski, M.K.; Cattaneo, R.; Atkinson, J.P. Membrane cofactor protein (MCP; CD46) expression in transgenic mice. Clin. Exp. Immunol. 2001, 124, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, C.J.; Franke, W.W.; Goerdt, S.; Falkowska-Hansen, B.; Rickelt, S.; Peitsch, W.K. Homo- and Heterotypic Cell Contacts in Malignant Melanoma Cells and Desmoglein 2 as a Novel Solitary Surface Glycoprotein. J. Investig. Dermatol. 2007, 127, 2191–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Chai, N.; Gu, Y.; Ding, L.; Yang, Y.; Zhou, J.; Ren, G.; Hao, X.; Fan, D.; Wu, K.; et al. Systematic Immunohistochemical Analysis of the Expression of CD46, CD55, and CD59 in Colon Cancer. Arch. Pathol. Lab. Med. 2014, 138, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Surowiak, P.; Materna, V.; Maciejczyk, A.; Kaplenko, I.; Spaczynski, M.; Dietel, M.; Lage, H.; Zabel, M. CD46 expression is indicative of shorter revival-free survival for ovarian cancer patients. Anticancer. Res. 2007, 26, 4943–4948. [Google Scholar]
- Thorsteinsson, L.; O’Dowd, G.M.; Harrington, P.M.; Johnson, P.M. The complement regulatory proteins CD46 and CD59, but not CD55, are highly expressed by glandular epithelium of human breast and colorectal tumour tissues. Apmis 1998, 106, 869–878. [Google Scholar] [CrossRef]
- Trojan, L.; Schaaf, A.; Steidler, A.; Haak, M.; Thalmann, G.; Knoll, T.; Gretz, N.; Alken, P.; Michel, M.S. Identification of metastasis-associated genes in prostate cancer by genetic profiling of human prostate cancer cell lines. Anticancer. Res. 2005, 25, 183–191. [Google Scholar] [CrossRef]
- Nilsson, M.; Ljungberg, J.; Richter, J.; Kiefer, T.; Magnusson, M.; Lieber, A.; Widegren, B.; Karlsson, S.; Fan, X. Development of an adenoviral vector system with adenovirus serotype 35 tropism; efficient transient gene transfer into primary malignant hematopoietic cells. J. Gene Med. 2004, 6, 631–641. [Google Scholar] [CrossRef]
- Campbell, M.; Qu, S.; Wells, S.; Sugandha, H.; Jensen, R.A. An adenoviral vector containing an arg–gly–asp (RGD) motif in the fiber knob enhances protein product levels from transgenes refractory to expression. Cancer Gene Ther. 2003, 10, 559–570. [Google Scholar] [CrossRef] [Green Version]
- Dmitriev, I.; Krasnykh, V.; Miller, C.R.; Wang, M.; Kashentseva, E.; Mikheeva, G.; Belousova, N.; Curiel, D.T. An Adenovirus Vector with Genetically Modified Fibers Demonstrates Expanded Tropism via Utilization of a Coxsackievirus and Adenovirus Receptor-Independent Cell Entry Mechanism. J. Virol. 1998, 72, 9706–9713. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Seki, T.; Dmitriev, I.; Uil, T.; Kashentseva, E.; Han, T.; Curiel, D.T. Double Modification of Adenovirus Fiber with RGD and Polylysine Motifs Improves Coxsackievirus–Adenovirus Receptor-Independent Gene Transfer Efficiency. Hum. Gene Ther. 2002, 13, 1647–1653. [Google Scholar] [CrossRef] [PubMed]
- Kelly, T.J., Jr.; Lewis, A.M., Jr. Use of Nondefective Adenovirus-Simian Virus 40 Hybrids for Mapping the Simian Virus 40 Genome. J. Virol. 1973, 12, 643–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, N.; Shenk, T. Isolation of deletion and substitution mutants of adenovirus type 5. Cell 1978, 13, 181–188. [Google Scholar] [CrossRef]
- Lichtenstein, D.L.; Toth, K.; Doronin, K.; Tollefson, A.E.; Wold, W.S. Functions and Mechanisms of Action of the Adenovirus E3 Proteins. Int. Rev. Immunol. 2004, 23, 75–111. [Google Scholar] [CrossRef] [PubMed]
- Benedict, C.A.; Norris, P.S.; Prigozy, T.I.; Bodmer, J.L.; Mahr, J.A.; Garnett, C.T.; Martinon, F.; Tschopp, J.; Gooding, L.R.; Ware, C.F. Three Adenovirus E3 Proteins Cooperate to Evade Apoptosis by Tumor Necrosis Factor-related Apoptosis-inducing Ligand Receptor-1 and -2. J. Biol. Chem. 2001, 276, 3270–3278. [Google Scholar] [CrossRef] [PubMed]
- Elsing, A.; Burgert, H.-G. The adenovirus E3/10.4K–14.5K proteins down-modulate the apoptosis receptor Fas/Apo-1 by inducing its internalization. Proc. Natl. Acad. Sci. USA 1998, 95, 10072–10077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNees, A.L.; Garnett, C.T.; Gooding, L.R. The Adenovirus E3 RID Complex Protects Some Cultured Human T and B Lymphocytes from Fas-Induced Apoptosis. J. Virol. 2002, 76, 9716–9723. [Google Scholar] [CrossRef] [Green Version]
- Shisler, J.; Yang, C.; Walter, B.; Ware, C.F.; Gooding, L.R. The adenovirus E3-10.4K/14.5K complex mediates loss of cell surface Fas (CD95) and resistance to Fas-induced apoptosis. J. Virol. 1997, 71, 8299–8306. [Google Scholar] [CrossRef] [Green Version]
- Tollefson, A.E.; Hermiston, T.W.; Lichtenstein, D.L.; Colle, C.F.; Tripp, R.A.; Dimitrov, T.; Toth, K.; Wells, C.E.; Doherty, P.C.; Wold, W.S. Forced degradation of Fas inhibits apoptosis in adenovirus-infected cells. Nature 1998, 392, 726–730. [Google Scholar] [CrossRef]
- Tollefson, A.E.; Toth, K.; Doronin, K.; Kuppuswamy, M.; Doronina, O.A.; Lichtenstein, D.L.; Hermiston, T.W.; Smith, C.A.; Wold, W.S. Inhibition of TRAIL-Induced Apoptosis and Forced Internalization of TRAIL Receptor 1 by Adenovirus Proteins. J. Virol. 2001, 75, 8875–8887. [Google Scholar] [CrossRef] [Green Version]
- Gooding, L.R.; Aquino, L.; Duerksen-Hughes, P.J.; Day, D.; Horton, T.M.; Yei, S.P.; Wold, W.S. The E1B 19,000-molecular-weight protein of group C adenoviruses prevents tumor necrosis factor cytolysis of human cells but not of mouse cells. J. Virol. 1991, 65, 3083–3094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krajcsi, P.; Dimitrov, T.; Hermiston, T.W.; E Tollefson, A.; Ranheim, T.S.; Vande Pol, S.B.; Stephenson, A.H.; Wold, W.S. The adenovirus E3-14.7K protein and the E3-10.4K/14.5K complex of proteins, which independently inhibit tumor necrosis factor (TNF)-induced apoptosis, also independently inhibit TNF-induced release of arachidonic acid. J. Virol. 1996, 70, 4904–4913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wold, W.S.; Doronin, K.; Toth, K.; Kuppuswamy, M.; Lichtenstein, D.L.; Tollefson, A.E. Immune responses to adenoviruses: Viral evasion mechanisms and their implications for the clinic. Curr. Opin. Immunol. 1999, 11, 380–386. [Google Scholar] [CrossRef]
- Moise, A.R.; Grant, J.R.; Vitalis, T.Z.; Jefferies, W.A. Adenovirus E3-6.7K Maintains Calcium Homeostasis and Prevents Apoptosis and Arachidonic Acid Release. J. Virol. 2002, 76, 1578–1587. [Google Scholar] [CrossRef] [Green Version]
- Gooding, L.R.; Ranheim, T.S.; Tollefson, A.E.; Aquino, L.; Duerksen-Hughes, P.; Horton, T.M.; Wold, W.S. The 10,400- and 14,500-dalton proteins encoded by region E3 of adenovirus function together to protect many but not all mouse cell lines against lysis by tumor necrosis factor. J. Virol. 1991, 65, 4114–4123. [Google Scholar] [CrossRef]
- Gooding, L.R.; Sofola, I.O.; Tollefson, A.E.; Duerksen-Hughes, P.; Wold, W.S. The adenovirus E3-14.7K protein is a general inhibitor of tumor necrosis factor-mediated cytolysis. J. Immunol. 1990, 145, 3080–3086. [Google Scholar] [CrossRef]
- Thorne, T.E.; Voelkel-Johnson, C.; Casey, W.M.; Parks, L.W.; Laster, S.M. The activity of cytosolic phospholipase A2 is required for the lysis of adenovirus-infected cells by tumor necrosis factor. J. Virol. 1996, 70, 8502–8507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; McMichael, A.; Peterson, P.A. Reduced allorecognition of adenovirus-2 infected cells. J. Immunol. 1987, 138, 3960–3966. [Google Scholar] [CrossRef]
- Andersson, M.; Paabo, S.; Nilsson, T.; Peterson, P.A. Impaired intracellular transport of class I MHC antigens as a possible means for adenoviruses to evade immune surveillance. Cell 1985, 43, 215–222. [Google Scholar] [CrossRef]
- Burgert, H.G.; Kvist, S. An adenovirus type 2 glycoprotein blocks cell surface expression of human histocompatibility class I antigens. Cell 1985, 41, 987–997. [Google Scholar] [CrossRef]
- Burgert, H.G.; Maryanski, J.L.; Kvist, S. “E3/19K” protein of adenovirus type 2 inhibits lysis of cytolytic T lymphocytes by blocking cell-surface expression of histocompatibility class I antigens. Proc. Natl. Acad. Sci. USA 1987, 84, 1356–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, J.H.; Bennink, J.R.; Yewdell, J.W. Retention of adenovirus E19 glycoprotein in the endoplasmic reticulum is essential to its ability to block antigen presentation. J. Exp. Med. 1991, 174, 1629–1637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawle, F.C.; Tollefson, A.E.; Wold, W.S.; Gooding, L.R. Mouse anti-adenovirus cytotoxic T lymphocytes. Inhibition of lysis by E3 gp19K but not E3 14.7K. J. Immunol. 1989, 143, 2031–2037. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.R.; Nilsson, T.; Peterson, P.A. Identification of a consensus motif for retention of transmembrane proteins in the endoplasmic reticulum. EMBO J. 1990, 9, 3153–3162. [Google Scholar] [CrossRef]
- Tollefson, A.E.; Scaria, A.; Hermiston, T.W.; Ryerse, J.S.; Wold, L.J.; Wold, W.S. The adenovirus death protein (E3-11.6K) is required at very late stages of infection for efficient cell lysis and release of adenovirus from infected cells. J. Virol. 1996, 70, 2296–2306. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.G.; Eidenschink, B.B.; Iguchi, E.; Zakharkin, S.O.; LaRocca, C.J.; Tolosa, E.J.; Truty, M.J.; Jacobsen, K.; Fernandez-Zapico, M.E.; Davydova, J. Cancer imaging and therapy utilizing a novel NIS-expressing adenovirus: The role of adenovirus death protein deletion. Mol. Ther. Oncolytics 2021, 20, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Davydova, J.; Gavrikova, T.; Brown, E.J.; Luo, X.; Curiel, D.T.; Vickers, S.M.; Yamamoto, M. In Vivo bioimaging tracks conditionally replicative adenoviral replication and provides an early indication of viral antitumor efficacy. Cancer Sci. 2010, 101, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Mathews, M.B.; Shenk, T. Adenovirus virus-associated RNA and translation control. J. Virol. 1991, 65, 5657–5662. [Google Scholar] [CrossRef] [Green Version]
- Akusjärvi, G.; Mathews, M.B.; Andersson, P.; Vennström, B.; Pettersson, U. Structure of genes for virus-associated RNAI and RNAII of adenovirus type 2. Proc. Natl. Acad. Sci. USA 1980, 77, 2424–2428. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Mathews, M.B. Structure, function, and evolution of adenovirus-associated RNA: A phylogenetic approach. J. Virol. 1996, 70, 5083–5099. [Google Scholar] [CrossRef] [Green Version]
- Bhat, R.A.; Thimmappaya, B. Adenovirus mutants with DNA sequence perturbations in the intragenic promoter of VAI RNA gene allow the enhanced transcription of VAII RNA gene in HeLa cells. Nucleic Acids Res. 1984, 12, 7377–7388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, S.; Bhat, R.A.; Rundell, M.K.; Thimmappaya, B. Suppression of the translation defect phenotype specific for a virus-associated RNA-deficient adenovirus mutant in monkey cells by simian virus 40. J. Virol. 1986, 60, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thimmappaya, B.; Weinberger, C.; Schneider, R.J.; Shenk, T. Adenovirus VAI RNA is required for efficient translation of viral mRNAs at late times after infection. Cell 1982, 31, 543–551. [Google Scholar] [CrossRef]
- Machitani, M.; Yamaguchi, T.; Shimizu, K.; Sakurai, F.; Katayama, K.; Kawabata, K.; Mizuguchi, H. Adenovirus Vector-Derived VA-RNA-Mediated Innate Immune Responses. Pharmaceutics 2011, 3, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Weber, F.; Wagner, V.; Kessler, N.; Haller, O. Induction of Interferon Synthesis by the PKR-Inhibitory VA RNAs of Adenoviruses. J. Interf. Cytokine Res. 2006, 26, 1–7. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Kawabata, K.; Kouyama, E.; Ishii, K.J.; Katayama, K.; Suzuki, T.; Kurachi, S.; Sakurai, F.; Akira, S.; Mizuguchi, H. Induction of type I interferon by adenovirus-encoded small RNAs. Proc. Natl. Acad. Sci. USA 2010, 107, 17286–17291. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ortega, M.B.; Lopez, G.J.; Jimenez, G.; Garcia-Garcia, J.A.; Conde, V.; Boulaiz, H.; Carrillo, E.; Perán, M.; Marchal, J.A.; Garcia, M.A. Clinical and therapeutic potential of protein kinase PKR in cancer and metabolism. Expert Rev. Mol. Med. 2017, 19, e9. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Bennett, R.L.; Liu, X.; Byrne, M.; Stratford May, W. PKR negatively regulates leukemia progression in association with PP2A activation, Bcl-2 inhibition and increased apoptosis. Blood Cancer J. 2013, 3, e144. [Google Scholar] [CrossRef] [Green Version]
- Marchal, J.A.; Lopez, G.J.; Peran, M.; Comino, A.; Delgado, J.R.; García-García, J.A.; Conde, V.; Aranda, F.M.; Rivas, C.; Esteban, M.; et al. The impact of PKR activation: From neurodegeneration to cancer. FASEB J. 2014, 28, 1965–1974. [Google Scholar] [CrossRef]
- Roh, M.S.; Kwak, J.Y.; Kim, S.J.; Lee, H.W.; Kwon, H.C.; Hwang, T.H.; Choi, P.J.; Hong, Y.S. Expression of double-stranded RNA-activated protein kinase in small-size peripheral adenocarcinoma of the lung. Pathol. Int. 2005, 55, 688–693. [Google Scholar] [CrossRef]
- Terada, T.; Ueyama, J.; Ukita, Y.; Ohta, T. Protein expression of double-stranded RNA-activated protein kinase (PKR) in intrahepatic bile ducts in normal adult livers, fetal livers, primary biliary cirrhosis, hepatolithiasis and intrahepatic cholangiocarcinoma. Liver 2000, 20, 450–457. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Summary of virus types currently in clinical trials as oncolytic agents. (Data source: clinicaltrials.gov, accessed on 1 October 2022).
Figure 1.
Summary of virus types currently in clinical trials as oncolytic agents. (Data source: clinicaltrials.gov, accessed on 1 October 2022).

{kind=link}
Table 1.
Adenoviral vectors currently in clinical trials for cancer therapy. Replication defective viruses that are used for gene delivery only are shaded in grey.
Table 1.
Adenoviral vectors currently in clinical trials for cancer therapy. Replication defective viruses that are used for gene delivery only are shaded in grey.
| Name | Serotype Based on | E1A | Fiber | Transgene | E3 Status | Other Modification | References |
|---|---|---|---|---|---|---|---|
| H101 (oncorin) | Ad5 | untouched | Ad5 | none | del of 78.3-85.8 m.u. Probably only E3-12.5K left | deletion of E1B55K | [8] |
| ADV/HSV-tk | Ad5 viral vector | transgene in deleted E1 | Ad5 | HSV tk gene with the RSV LTR promoter | Deletion of RIDα, RIDβ and E3-14.7K | [9] | |
| MEM-288 | Ad5 | delta 24 | Ad5 | chimeric CD40 ligand and IFNb | deleted | deletion of E1B55K | [10] |
| LOAd703 | Ad5 | E2F promoter, delta 24 | chimeric Ad5/Ad35 | TMZ-CD40L + 4-1BBL | deleted E3-6.7K and gp19K | [11,12,13] | |
| CG0070 | Ad5 | E2F promoter | Ad5 | GM-CSF under control of E3 promoter | gp19K deleted | [14] | |
| AdAPT-001 | Ad5 | 50bp del in E1A promoter (tumor specific) | Ad5 | mTGFbR-IgG (TGFb trap) | Deletion of RIDα, RIDβ and E3-14.7K | deletion E1B19K | [15,16] |
| OBP-301 (Telomelysin) | Ad5 | human TERT promoter E1A-IRES-E1B in E1 deleted backbone | Ad5 | none | deleted | [17,18] | |
| Colo-Ad1 (Enadenotucirev) | Ad11/Ad3 chimera | untouched (Ad11) | Ad11 | none | near complete deletion | deletion in E4orf4 gene | [19] |
| DNX-2440 | Ad5 | delta 24 | Ad5 with RGD-motif into the H-loop | OX40 ligand | deleted | [20,21] | |
| CAdVEC | Ad5 + Ad5 gutted vector | delta24 | Ad5 | undisclosed immunomodulatory molecules in vector | deleted | ||
| CGTG-102(ONCOS-102) | Ad5 | delta 24 | Ad5 with fiber knob of Ad3 | GM-CSF under control of E3 promoter | E3-6.7K and gp19K deleted | [22,23,24] | |
| DNX-2401 | Ad5 | delta 24 | Ad5 with RGD-motif into the H-loop | none | deleted | [21] | |
| TILT-123 | Ad5 | E2F promoter, delta 24 | Ad5 with fiber knob of Ad3 | TNFa and IL-2 | transgenes in E3 unclear what is deleted | [25] | |
| Ad-E6E7 | Ad5 viral vector (vaccine) | E1 deleted | Ad5 | attenuated fusion E6E7 transgene | deleted | + Maraba virus | [26] |
| Ad-MAGEA3 | Ad5 viral vector (vaccine) | E1 deleted | Ad5 | MAGE-A3 | deleted | + Maraba virus | [27] |
| VCN-01 | Ad5 | E2F promoter, delta 24 | Ad5 with RGDK motif in shaft | soluble version of human PH20 under late promoter | WT | [28] | |
| NG-350A | Ad11/Ad3 chimera | untouched (Ad11) | Ad11 | heavy and light chains for a secreted CD40 agonist monoclonal antibody | near complete deletion | PsiOxus Therapeutics | |
| NG-641 | Ad11/Ad3 chimera | untouched (Ad11) | Ad11 | secreted Interferon alpha, the chemokines CXCL9, CXCL10 and an anti-FAP/anti-CD3 bispecific T-cell activator | near complete deletion | PsiOxus Therapeutics | |
| cells with CRAd-survivin-pk7 virus | Ad5 | human survivin promoter | Ad5 modified with polylysine | none | WT | [29] | |
| ORCA-010 | Ad5 | delta 24 | Ad5 with RGD-motif into the H-loop | none | mutated GP19K | T1 mutation in E3-19K | [30] |
Table 2.
List of clinical trials currently using oncolytic Adenoviral vectors as oncolytic agents. (Data source: clinicaltrials.gov, accessed on October 2022).
Table 2.
List of clinical trials currently using oncolytic Adenoviral vectors as oncolytic agents. (Data source: clinicaltrials.gov, accessed on October 2022).
| Trial Identifier | Phase | Conditions | Virus Name | E3 Status | Additional Treatment |
|---|---|---|---|---|---|
| NCT05051696 | N/A | Genital Neoplasms, Female | Oncorine (H101) | only 12.5K present | +/- radiotherapy |
| NCT04771676 | 2 | Refractory Malignant Ascites | Oncorine (H101) | only 12.5K present | none |
| NCT03004183 | 2 | Metastatic Non-small Cell Lung Cancer, Metastatic Triple-negative Breast Cancer | ADV/HSV-tk | deletion of RIDα/β and 14.7K | Valacyclovir, SBRT, Pembrolizumab |
| NCT05076760 | 1 | Advanced Solid Tumors | MEM-288 | deleted | MEM-288 Intratumoral Injection |
| NCT02705196 | 1/2 | Pancreatic Cancer | LOAd703 | deletion of 6.7K and gp19K | gemcitabine, nab-paclitaxel, atezolizumab |
| NCT02143804 | 2 | High Grade, Bladder Cancer, Non Muscle Invasive | CG0070 | gp19K deletion | none |
| NCT02365818 | 2 | Bladder Cancer | CG0070 | gp19K deletion | CG0070 |
| NCT05234905 | 2 | Uterine Cervical Neoplasms | Oncorine (H101) | only 12.5K present | Camrelizumab |
| NCT05113290 | 4 | Hepatocellular Carcinoma | Oncorine (H101) | only 12.5K present | Sorafenib |
| NCT05124002 | 4 | Cholangiocarcinoma, Intrahepatic | Oncorine (H101) | only 12.5K present | HAIC or FOLFOX |
| NCT04673942 | 1 | Refractory Solid Tumor, Adult | AdAPT-001 | deletion of RIDα/β and 14.7K | none |
| NCT03190824 | 2 | Melanoma Stage III and iv | OBP-301 (Telomelysin) | deleted | none |
| NCT01438112 | 2 | Non Muscle Invasive Bladder Cancer | CG0070 | gp19K deletion | none |
| NCT03916510 | 1 | Locally Advanced Rectal Cancer | Enadenotucirev (previously ColoAd-1) | near complete deletion | Capecitabine, Radiotherapy |
| NCT03714334 | 1 | Glioblastoma | DNX-2440 | deleted | none |
| NCT03740256 | 1 | Various solid tumors | CAdVEC | deleted | none |
| NCT01437280 | 1 | Solid Tumors | CGTG-102 | 6.7K and gp19K deleted | none |
| NCT05180851 | 1 | Various solid tumors | Recombinant L-IFN adenovirus | unknown | none |
| NCT02197169 | 1 | Glioblastoma or Gliosarcoma | DNX-2401 | deleted | Interferon-gamma |
| NCT02053220 | 1 | Solid Cancers | Colo-Ad1 | near complete deletion | none |
| NCT02028442 | 1/2 | Solid Tumors of Epithelial Origin | Enadenotucirev (previously ColoAd-1) | near complete deletion | none |
| NCT02028117 | 1 | Recurrent Platinum Resistant Ovarian Cancer | Enadenotucirev (previously ColoAd-1) | near complete deletion | none |
| NCT05222932 | 1 | Melanoma, Head and Neck Squamous Cell Carcinoma | TILT-123 | unknown deletion | Avelumab |
| NCT01598129 | 1 | Malignant Solid Tumor | CGTG-102 | 6.7K and gp19K deleted | low-dose metronomic cyclophosphamide. |
| NCT02798406 | 2 | Brain Cancers | DNX-2401 | deleted | pembrolizumab |
| NCT04217473 | 1 | Metastatic Melanoma | TILT-123 | unknown deletion | TILT-123 |
| NCT03618953 | 1 | HPV-Associated Cancers | Ad-E6E7, MG1-E6E7 | deleted | Atezolizumab |
| NCT04695327 | 1 | Solid Tumor | TILT-123 | unknown deletion | TILT-123 |
| NCT03773744 | 1 | Metastatic Melanoma, Squamous Cell Skin Carcinoma | Ad-MAGEA3, MG1-MAGEA3 | deleted | Pembrolizumab, Cyclophosphamide |
| NCT02045602 | 1 | Locally Advanced Solid Tumors | VCN-01 | WT | Gemcitabine, Abraxane® |
| NCT02045589 | 1 | Pancreatic Adenocarcinoma | VCN-01 | WT | Gemcitabine, Abraxane® |
| NCT02879760 | 1/2 | Non-Small Cell Lung Cancer | Ad-MAGEA3, MG1-MAGEA3 | deleted | Pembrolizumab |
| NCT04685499 | 2 | Head and Neck Squamous Cell Carcinoma With Inoperable Recurrent or Progressive Disease | OBP-301 | deleted | Pembrolizumab |
| NCT03225989 | 1/2 | Solid Cancers | LOAd703 | deletion of 6.7K and gp19K | none |
| NCT03896568 | 1 | High Grade Glioma | DNX-2401 | deleted | none |
| NCT01956734 | 1 | Glioblastoma Multiforme | DNX-2401 | deleted | Temozolomide |
| NCT03178032 | 1 | Brainstem Glioma | DNX-2401 | deleted | none |
| NCT03852511 | 1 | Metastatic Cancer | NG-350A | near complete deletion | none |
| NCT04053283 | 1 | Metastatic Cancer | NG-641 | near complete deletion | none |
| NCT03072134 | 1 | Malignant Glioma | Neural stem cells loaded with NSC-CRAd-Survivin-pk7 | WT | none |
| NCT04097002 | 1/2 | Adenocarcinoma of the Prostate | ORCA-010 | mutated gp19K | none |
| NCT05561491 | 2 | Melanoma | ONCOS-102 (Previously known as CGTG-102) | 6.7K and gp19K deleted | Balstilimab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Blanchette, P.; Teodoro, J.G. A Renaissance for Oncolytic Adenoviruses? Viruses 2023, 15, 358. https://doi.org/10.3390/v15020358
AMA Style
Blanchette P, Teodoro JG. A Renaissance for Oncolytic Adenoviruses? Viruses. 2023; 15(2):358. https://doi.org/10.3390/v15020358
Chicago/Turabian StyleBlanchette, Paola, and Jose G. Teodoro. 2023. "A Renaissance for Oncolytic Adenoviruses?" Viruses 15, no. 2: 358. https://doi.org/10.3390/v15020358
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.