
Identification of Two Isoforms of Canine Tetherin in Domestic Dogs and Characterization of Their Antiviral Activity against Canine Influenza Virus
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Cells
2.2. Plasmids
2.3. Expression of Canine Tetherin
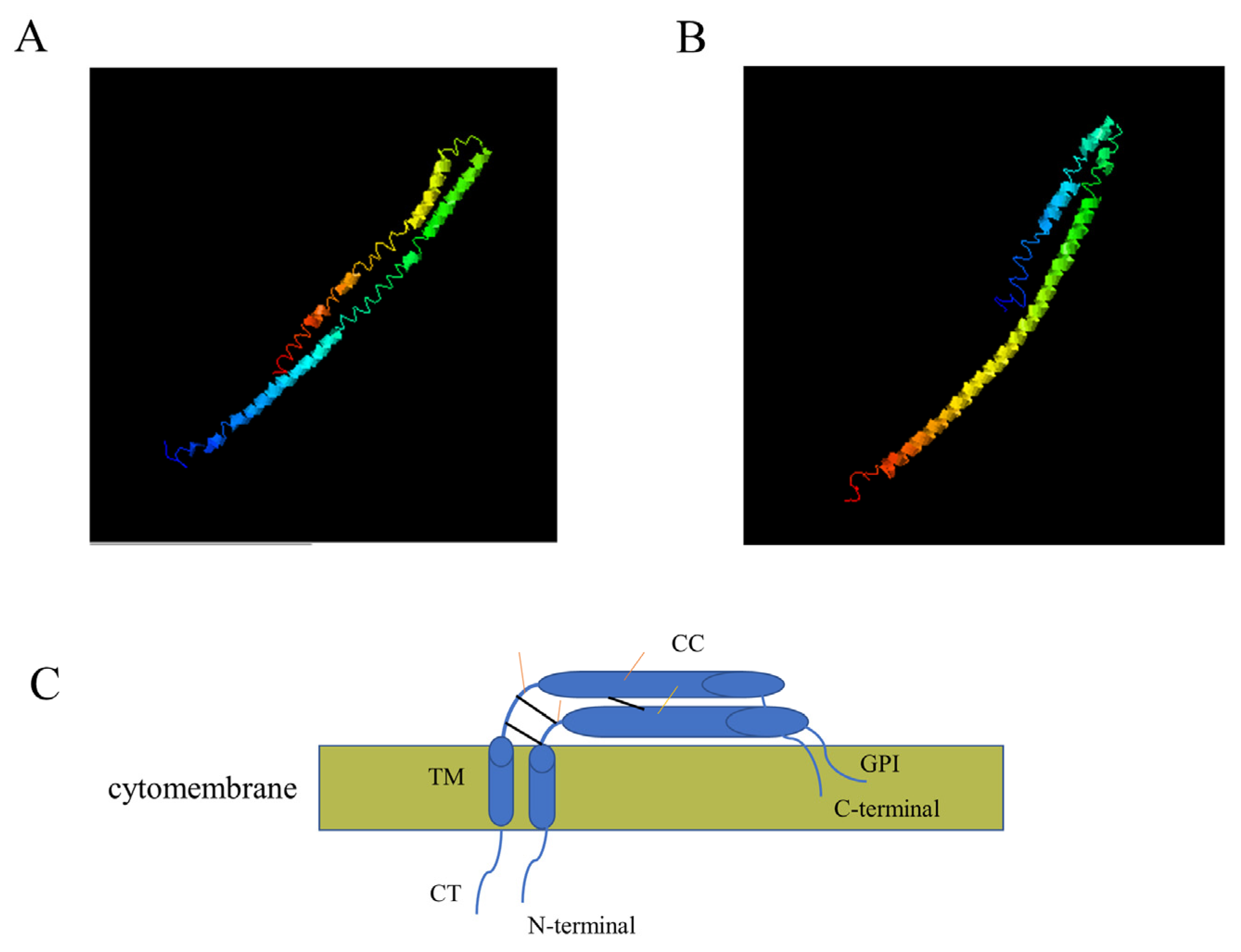
2.4. Bioinformatics Analysis
2.5. Immunofluorescence Staining and Confocal Microscopy
2.6. Assessment of Whether Canine Tetherin Restricts the Release of CIV
2.6.1. Viral Infection
2.6.2. Viral Titer Determination
2.6.3. Reverse Transcription Polymerase Chain Reaction (RT–qPCR) Analysis
2.6.4. Western Blot Analysis
3. Results
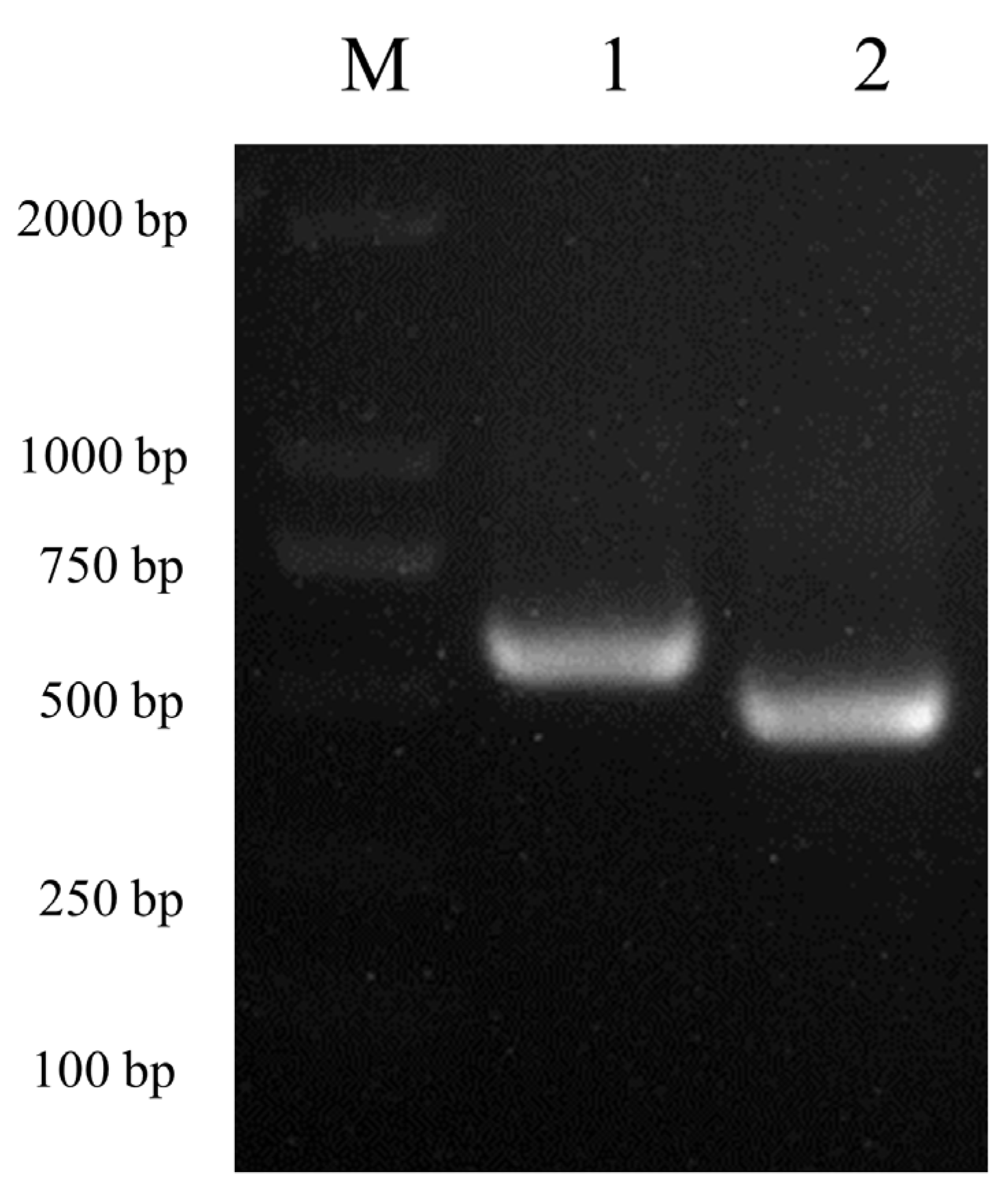
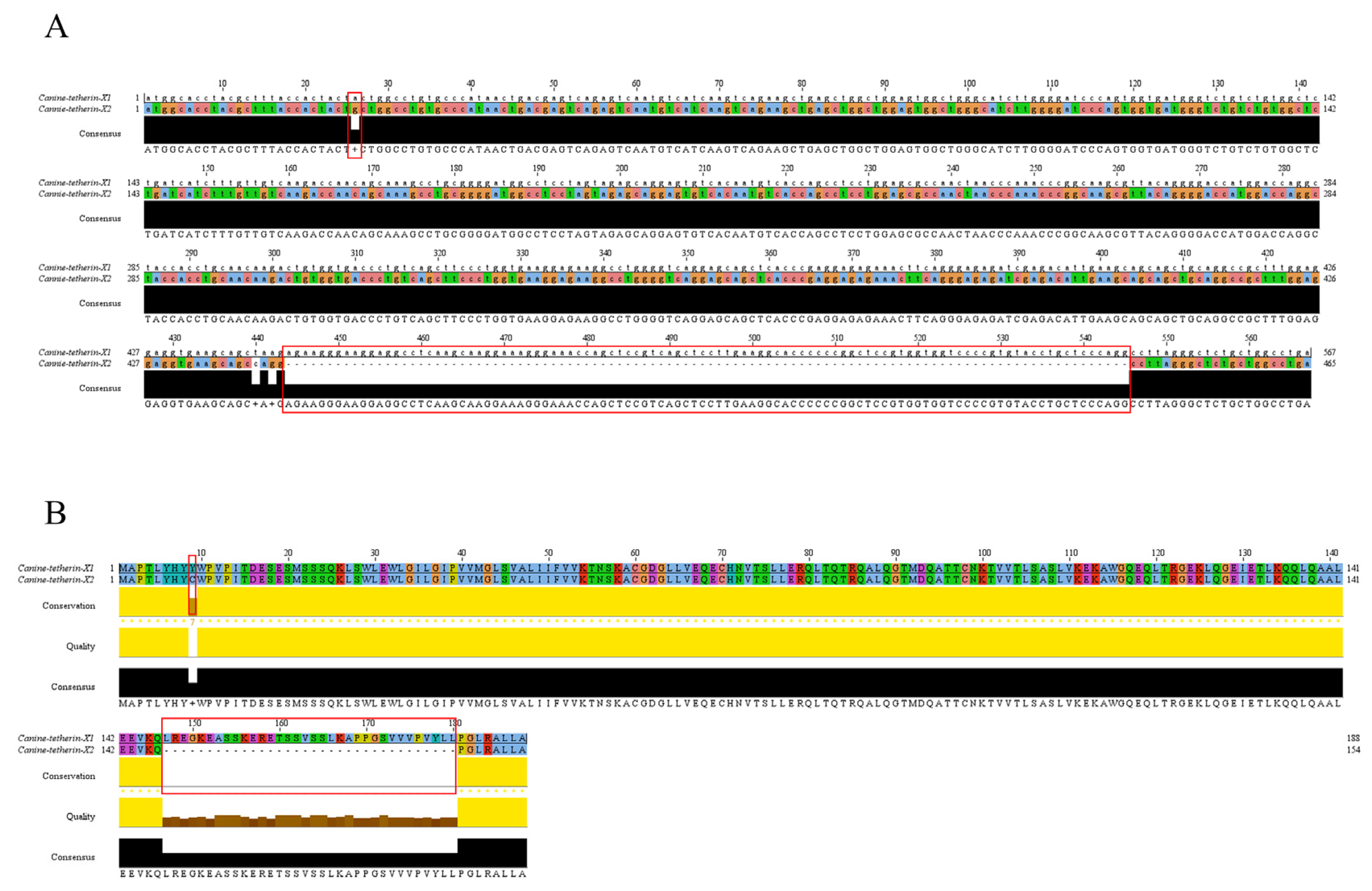
3.1. Amplification of Two Canine Tetherin Isoforms (Tetherin-X1 and Tetherin-X2)
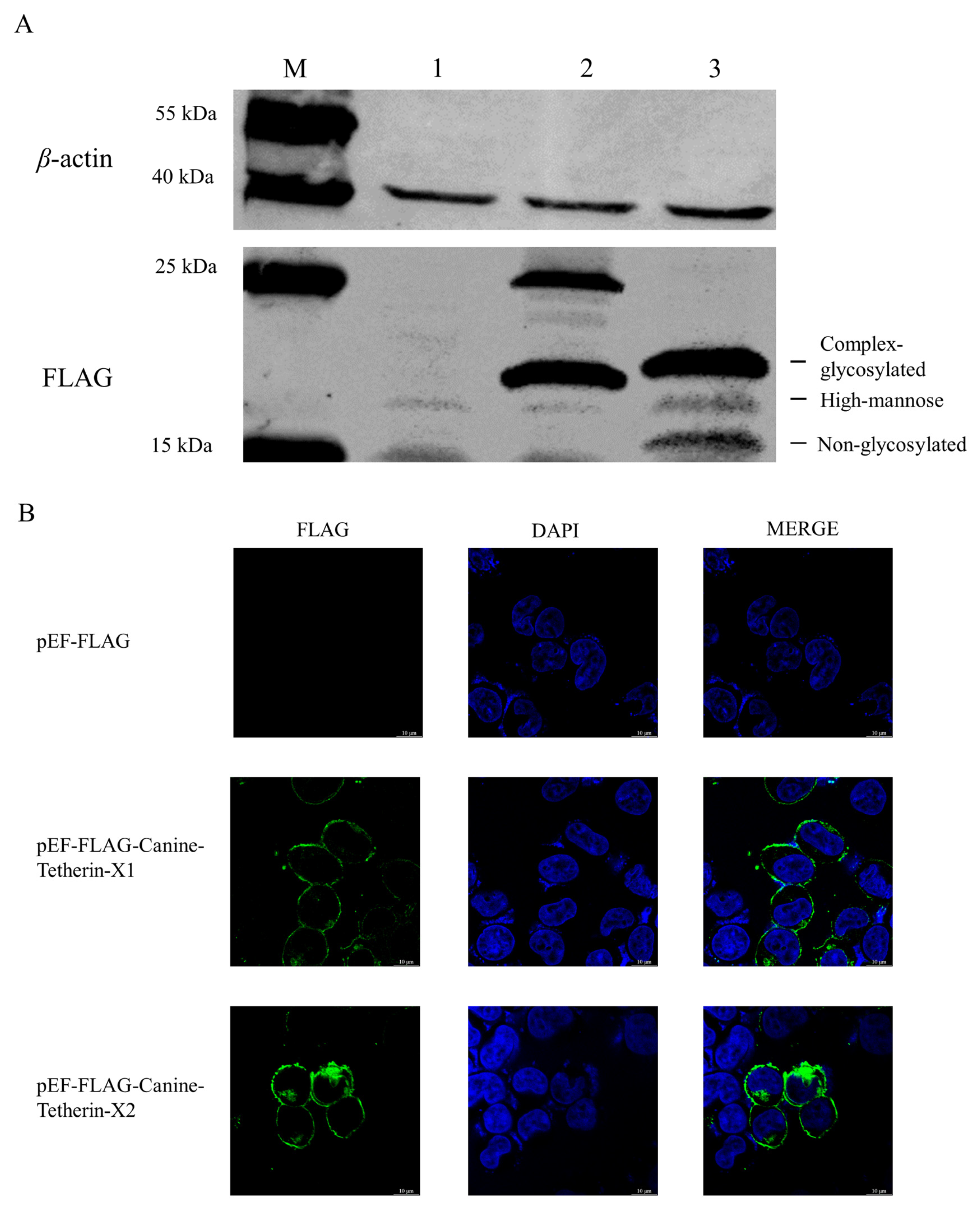
3.2. Expression and Localization of the Two Canine Tetherin Isoforms
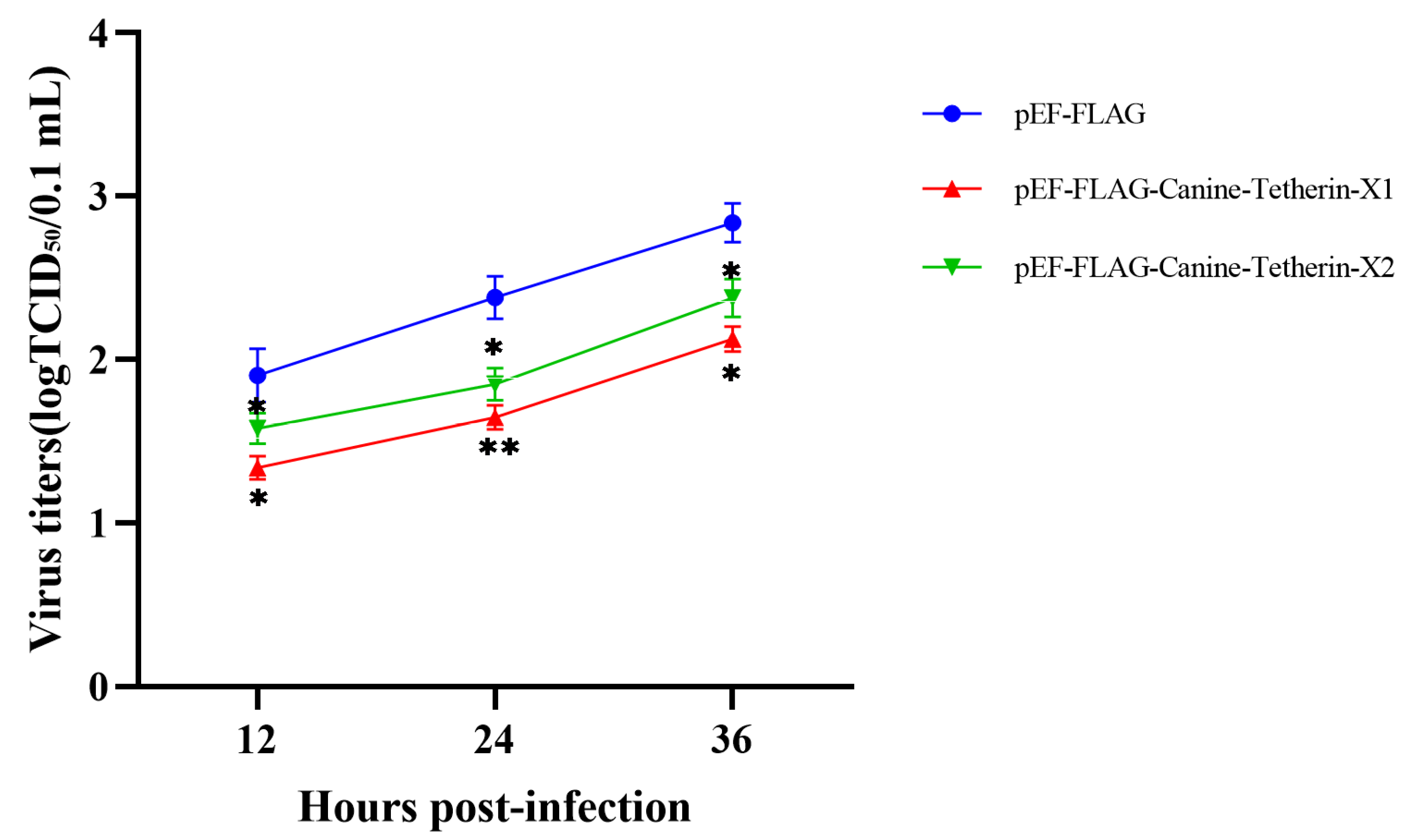
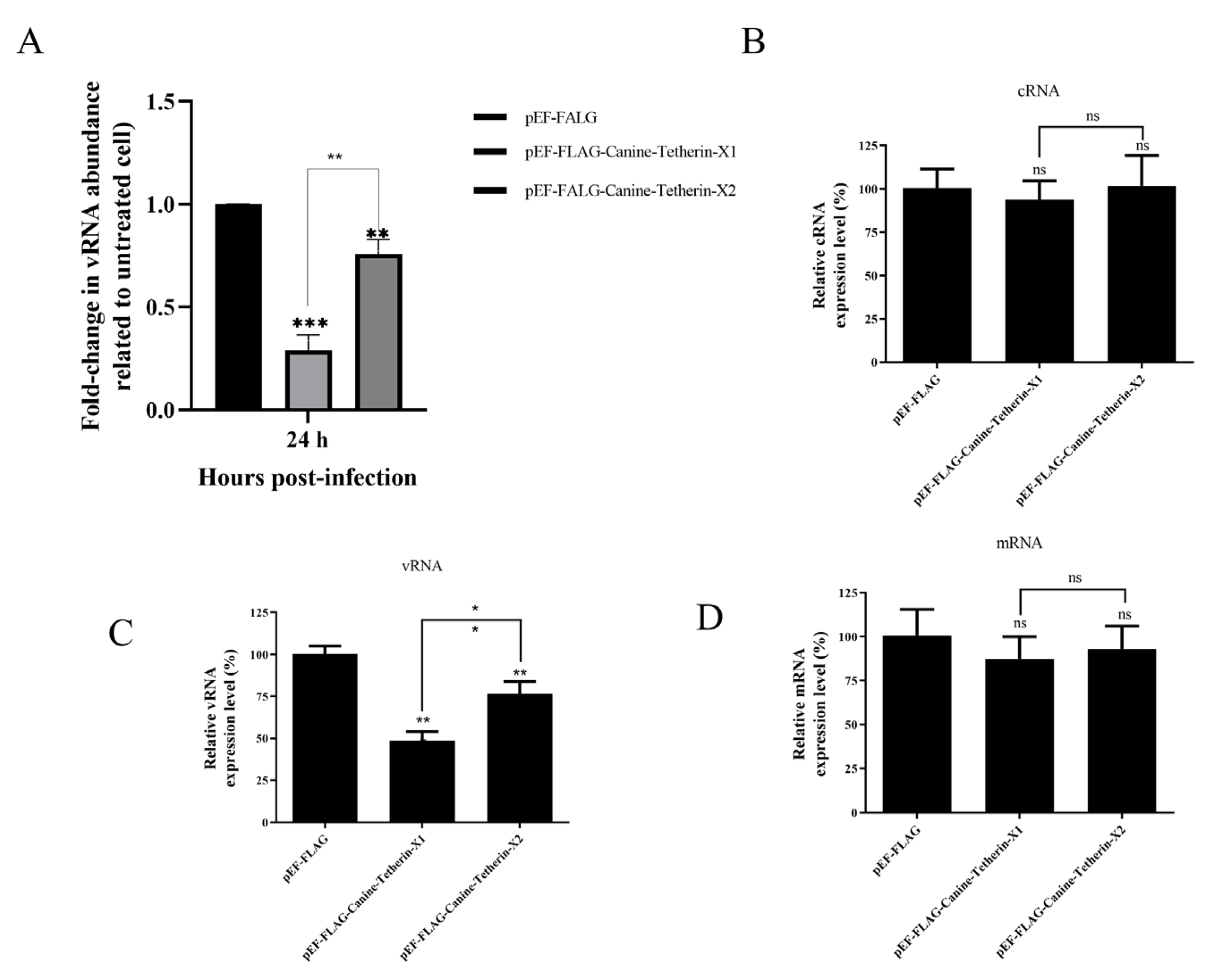
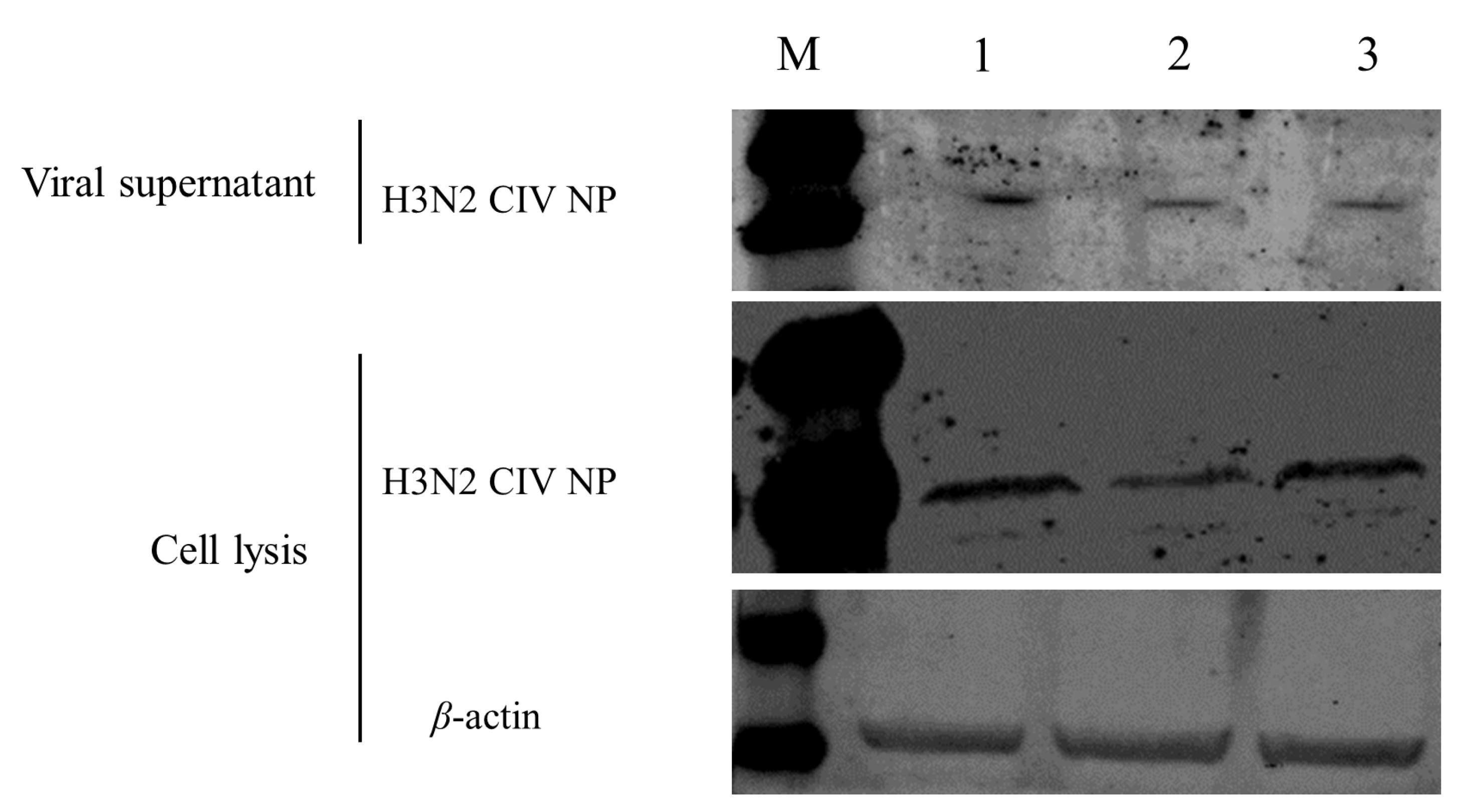
3.3. Canine Tetherin Restricts the Release of CIV
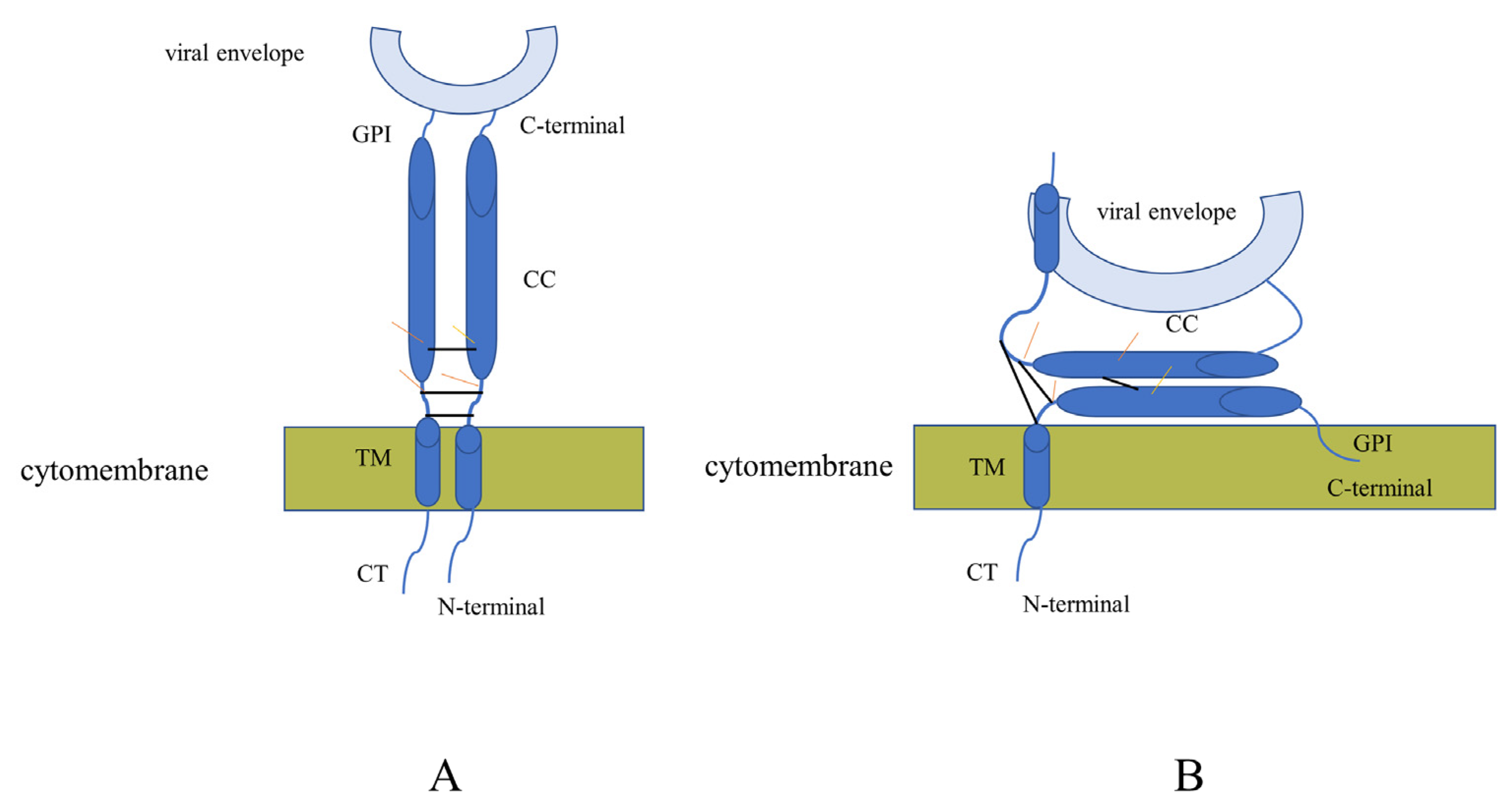
3.4. Incidence of Truncated Canine Tetherin Isoforms
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jin, S.; Tian, S.; Luo, M.; Xie, W.; Liu, T.; Duan, T.; Wu, Y.; Cui, J. Tetherin suppresses type i interferon signaling by targeting mavs for ndp52-mediated selective autophagic degradation in human cells. Mol. Cell 2017, 68, 308–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoggins, J.W.; Wilson, S.J.; Panis, M.; Murphy, M.Y.; Jones, C.T.; Bieniasz, P.; Rice, C.M. A diverse range of gene products are effectors of the type i interferon antiviral response. Nature 2011, 472, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, A. A virological view of innate immune recognition. Annu. Rev. Microbiol. 2012, 66, 177–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieniasz, P.D. Intrinsic immunity: A front-line defense against viral attack. Nat. Immunol. 2004, 5, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Yamamoto, S.P.; Misawa, N.; Yoshida, T.; Miyazawa, T.; Koyanagi, Y. Comparative study on the effect of human bst-2/tetherin on hiv-1 release in cells of various species. Retrovirology 2009, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Winkler, M.; Bertram, S.; Gnirss, K.; Nehlmeier, I.; Gawanbacht, A.; Kirchhoff, F.; Ehrhardt, C.; Ludwig, S.; Kiene, M.; Moldenhauer, A.S.; et al. Influenza a virus does not encode a tetherin antagonist with vpu-like activity and induces ifn-dependent tetherin expression in infected cells. PLoS ONE 2012, 7, e43337. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, S.; Bieniasz, P.D. Mechanism of hiv-1 virion entrapment by tetherin. PLoS Pathog. 2013, 9, e1003483. [Google Scholar] [CrossRef] [Green Version]
- Perez-Caballero, D.; Zang, T.; Ebrahimi, A.; Mcnatt, M.W.; Gregory, D.A.; Johnson, M.C.; Bieniasz, P.D. Tetherin inhibits hiv-1 release by directly tethering virions to cells. Cell 2009, 139, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Jouvenet, N.; Neil, S.J.; Zhadina, M.; Zang, T.; Kratovac, Z.; Lee, Y.; Mcnatt, M.; Hatziioannou, T.; Bieniasz, P.D. Broad-spectrum inhibition of retroviral and filoviral particle release by tetherin. J. Virol. 2009, 83, 1837–1844. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, T.; Noda, T.; Urata, S.; Kawaoka, Y.; Yasuda, J. Inhibition of lassa and marburg virus production by tetherin. J. Virol. 2009, 83, 2382–2385. [Google Scholar] [CrossRef]
- Kong, W.S.; Irie, T.; Yoshida, A.; Kawabata, R.; Kadoi, T.; Sakaguchi, T. Inhibition of virus-like particle release of sendai virus and nipah virus, but not that of mumps virus, by tetherin/cd317/bst-2. Hiroshima J. Med. Sci. 2012, 61, 59–67. [Google Scholar]
- Mansouri, M.; Viswanathan, K.; Douglas, J.L.; Hines, J.; Gustin, J.; Moses, A.V.; Fruh, K. Molecular mechanism of bst2/tetherin downregulation by k5/mir2 of kaposi’s sarcoma-associated herpesvirus. J. Virol. 2009, 83, 9672–9681. [Google Scholar] [CrossRef] [Green Version]
- Kaletsky, R.L.; Francica, J.R.; Agrawal-Gamse, C.; Bates, P. Tetherin-mediated restriction of filovirus budding is antagonized by the ebola glycoprotein. Proc. Natl. Acad. Sci. USA 2009, 106, 2886–2891. [Google Scholar] [CrossRef] [Green Version]
- Pardieu, C.; Vigan, R.; Wilson, S.J.; Calvi, A.; Zang, T.; Bieniasz, P.; Kellam, P.; Towers, G.J.; Neil, S.J. The ring-ch ligase k5 antagonizes restriction of kshv and hiv-1 particle release by mediating ubiquitin-dependent endosomal degradation of tetherin. PLoS Pathog. 2010, 6, e1000843. [Google Scholar] [CrossRef]
- Zheng, Y.; Hao, X.; Zheng, Q.; Lin, X.; Zhang, X.; Zeng, W.; Ding, S.; Zhou, P.; Li, S. Canine influenza virus is mildly restricted by canine tetherin protein. Viruses 2018, 10, 565. [Google Scholar] [CrossRef] [Green Version]
- Kupzig, S.; Korolchuk, V.; Rollason, R.; Sugden, A.; Wilde, A.; Banting, G. Bst-2/hm1. 24 is a raft-associated apical membrane protein with an unusual topology. Traffic 2003, 4, 694–709. [Google Scholar] [CrossRef]
- Heusinger, E.; Kluge, S.F.; Kirchhoff, F.; Sauter, D. Early vertebrate evolution of the host restriction factor tetherin. J. Virol. 2015, 89, 12154–12165. [Google Scholar] [CrossRef] [Green Version]
- Ozcan, K.A.; Berndsen, C.E. Bending of the bst-2 coiled-coil during viral budding. Proteins 2017, 85, 2081–2087. [Google Scholar] [CrossRef]
- Hayward, J.A.; Tachedjian, M.; Johnson, A.; Irving, A.T.; Gordon, T.B.; Cui, J.; Nicolas, A.; Smith, I.; Boyd, V.; Marsh, G.A.; et al. Unique evolution of antiviral tetherin in bats. J. Virol. 2022, 96, e115222. [Google Scholar] [CrossRef]
- Lu, G.; Ou, J.; Cai, S.; Lai, Z.; Zhong, L.; Yin, X.; Li, S. Canine interferon-inducible transmembrane protein is a host restriction factor that potently inhibits replication of emerging canine influenza virus. Front. Immunol. 2021, 12, 710705. [Google Scholar] [CrossRef]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of equine influenza virus to dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Kang, B.; Lee, C.; Jung, K.; Ha, G.; Kang, D.; Park, S.; Park, B.; Oh, J. Transmission of avian influenza virus (h3n2) to dogs. Emerg. Infect. Dis 2008, 14, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Rossman, J.S.; Lamb, R.A. Influenza virus assembly and budding. Virology 2011, 411, 229–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnirss, K.; Zmora, P.; Blazejewska, P.; Winkler, M.; Lins, A.; Nehlmeier, I.; Gartner, S.; Moldenhauer, A.S.; Hofmann-Winkler, H.; Wolff, T.; et al. Tetherin sensitivity of influenza a viruses is strain specific: Role of hemagglutinin and neuraminidase. J. Virol. 2015, 89, 9178–9188. [Google Scholar] [CrossRef] [Green Version]
- Tavakoli-Tameh, A.; Janaka, S.K.; Zarbock, K.; O’Connor, S.; Crosno, K.; Capuano, S.R.; Uno, H.; Lifson, J.D.; Evans, D.T. Loss of tetherin antagonism by nef impairs siv replication during acute infection of rhesus macaques. PLoS Pathog. 2020, 16, e1008487. [Google Scholar] [CrossRef] [Green Version]
- Mangeat, B.; Cavagliotti, L.; Lehmann, M.; Gers-Huber, G.; Kaur, I.; Thomas, Y.; Kaiser, L.; Piguet, V. Influenza virus partially counteracts restriction imposed by tetherin/bst-2. J. Biol. Chem. 2012, 287, 22015–22029. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Pang, X.; Li, J.; Cen, S.; Jin, Q.; Guo, F. The role of the structural domains of human bst-2 in inhibiting the release of xenotropic murine leukemia virus-related virus. Biochem. Biophys. Res. Commun. 2012, 428, 17–23. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Ou, J.; Hu, X.; Zheng, Y.; Ye, S.; Zhong, L.; Lai, Z.; Cai, S.; Lu, G.; Li, S. Identification of Two Isoforms of Canine Tetherin in Domestic Dogs and Characterization of Their Antiviral Activity against Canine Influenza Virus. Viruses 2023, 15, 393. https://doi.org/10.3390/v15020393
Xu L, Ou J, Hu X, Zheng Y, Ye S, Zhong L, Lai Z, Cai S, Lu G, Li S. Identification of Two Isoforms of Canine Tetherin in Domestic Dogs and Characterization of Their Antiviral Activity against Canine Influenza Virus. Viruses. 2023; 15(2):393. https://doi.org/10.3390/v15020393
Chicago/Turabian StyleXu, Liang, Jiajun Ou, Xuerui Hu, Yanhong Zheng, Shaotang Ye, Lintao Zhong, Zhiying Lai, Siqi Cai, Gang Lu, and Shoujun Li. 2023. "Identification of Two Isoforms of Canine Tetherin in Domestic Dogs and Characterization of Their Antiviral Activity against Canine Influenza Virus" Viruses 15, no. 2: 393. https://doi.org/10.3390/v15020393