
Hemorrhagic Fever with Renal Syndrome in Asia: History, Pathogenesis, Diagnosis, Treatment, and Prevention


 , ,
, , 
 and
and
Abstract
:1. Introduction
2. History of Orthohantaviruses and the Disease They Cause in Asia
3. Clinical Presentation of Orthohantavirus Disease in Asia
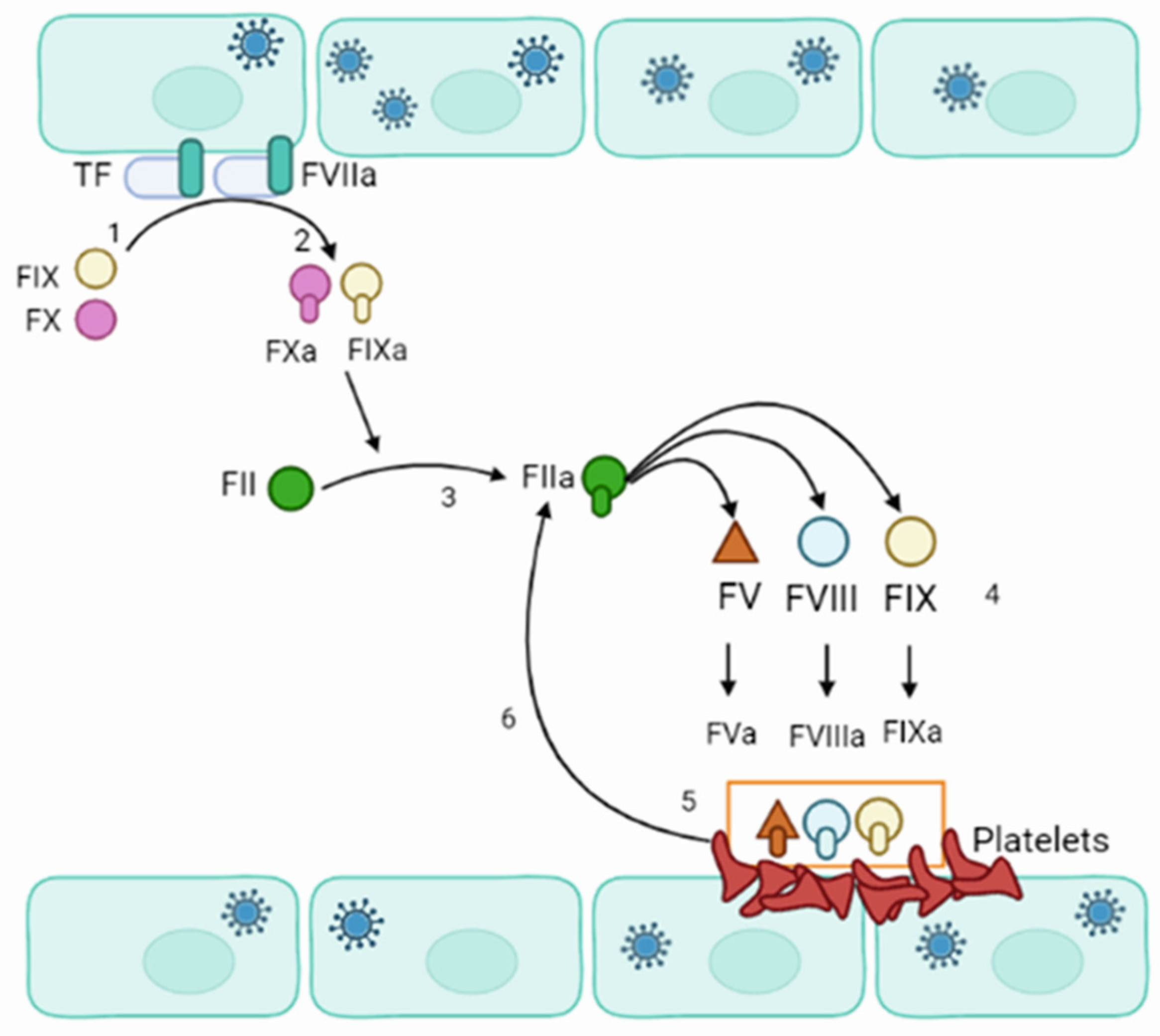
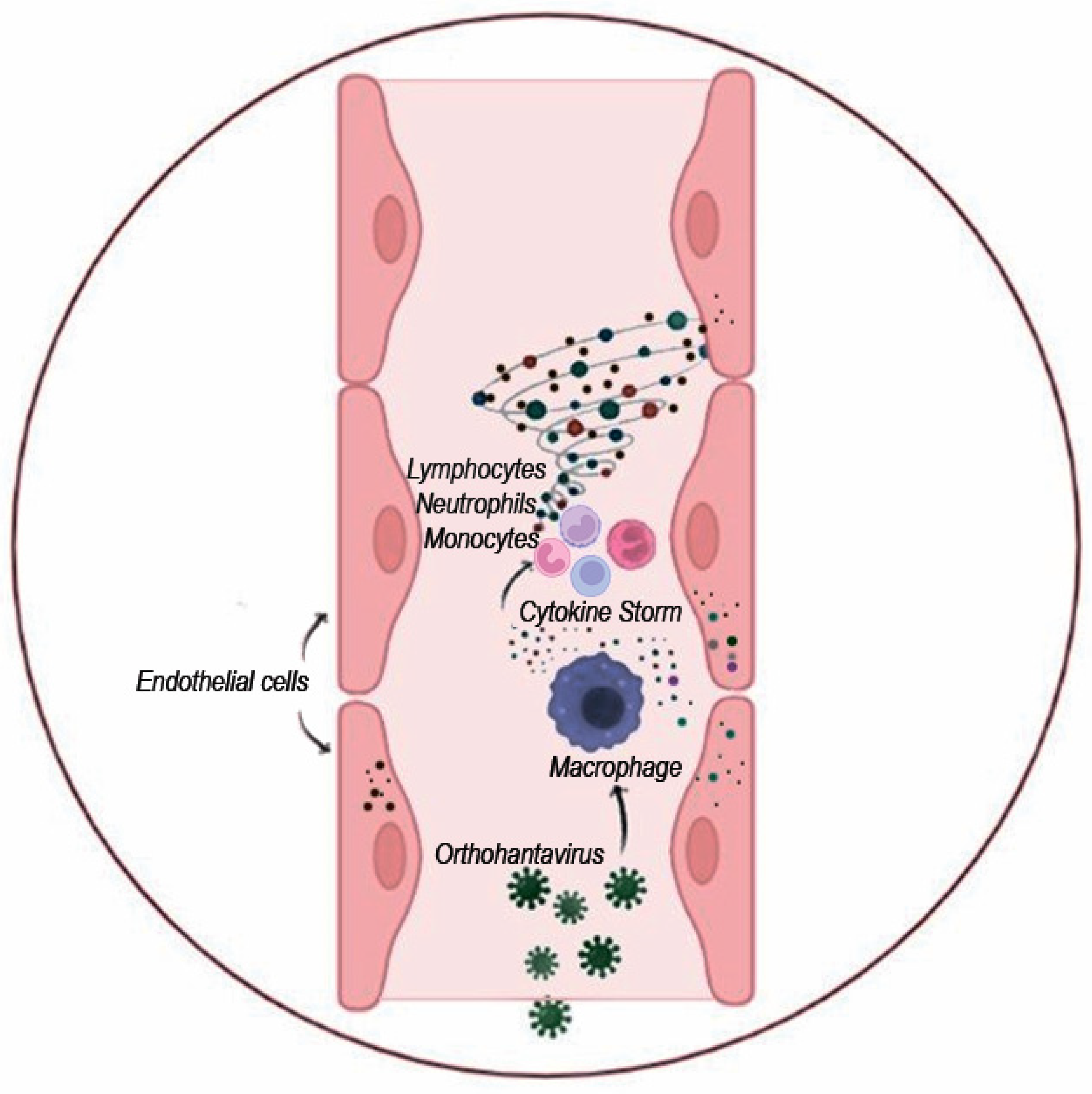
4. Pathogenesis of Orthohantavirus Infections
5. Vaccines and Prevention of Hantavirus Infections
6. Treatment of Orthohantavirus Infection
7. Challenges of Orthohantavirus Diagnostics and Treatment
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICTV Current ICTV Taxonomy Release. Available online: https://ictv.global/taxonomy (accessed on 8 August 2022).
- Laenen, L.; Vergote, V.; Calisher, C.H.; Klempa, B.; Klingström, J.; Kuhn, J.H.; Maes, P. Hantaviridae: Current Classification and Future Perspectives. Viruses 2019, 11, 788. [Google Scholar] [CrossRef] [Green Version]
- Noack, D.; Goeijenbier, M.; Reusken, C.B.E.M.; Koopmans, M.P.G.; Rockx, B.H.G. Orthohantavirus Pathogenesis and Cell Tropism. Front. Cell. Infect. Microbiol. 2020, 10, 399. [Google Scholar] [CrossRef]
- Kariwa, H.; Yoshimatsu, K.; Arikawa, J. Hantavirus Infection in East Asia. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 341–356. [Google Scholar] [CrossRef]
- Chandy, S.; Abraham, P.; Sridharan, G. Hantaviruses: An Emerging Public Health Threat in India? A Review. J. Biosci. 2008, 33, 495–504. [Google Scholar] [CrossRef]
- Brummer-Korvenkontio, M.; Manni, T.; Ukkonen, S.; Vaheri, A. Detection of Hemagglutination-Inhibiting Antibodies in Patients with Nephropathia Epidemica and Korean Hemorrhagic Fever by Using Puumala Virus Cell Culture Antigen. J. Infect. Dis. 1986, 153, 997–998. [Google Scholar] [CrossRef]
- Taller, A.; Xiao, S.; Godec, M.; Gligic, A. Belgrade Virus, a Cause of Hemorrhagic Fever with Renal Syndrome in the Balkans, Is Closely Related to Dobrava Virus of Field Mice. J. Infect. 1993, 3, 123–145. [Google Scholar]
- Lee, H.W.; Baek, L.J.; Johnson, K.M. Isolation of Hantaan Virus, the Etiologic Agent of Korean Hemorrhagic Fever, from Wild Urban Rats. J. Infect. Dis. 1982, 146, 638–644. [Google Scholar] [CrossRef]
- Jiang, H.; Zheng, X.; Wang, L.; Du, H.; Wang, P.; Bai, X. Hantavirus Infection: A Global Zoonotic Challenge. Virol. Sin. 2017, 32, 32–43. [Google Scholar] [CrossRef]
- Chen, H.X.; Qiu, F.X. Epidemiologic Surveillance on the Hemorrhagic Fever with Renal Syndrome in China. Chin. Med. J. (Engl.) 1993, 106, 857–863. [Google Scholar]
- Zhang, Y.Z.; Zou, Y.; Fu, Z.F.; Plyusnin, A. Hantavirus Infections in Humans and Animals, China. Emerg. Infect. Dis. 2010, 16, 1195. [Google Scholar] [CrossRef]
- Zou, L.-X.; Sun, L. Analysis of Hemorrhagic Fever with Renal Syndrome Using Wavelet Tools in Mainland China, 2004–2019. Front. Public Health 2020, 8, 571984. [Google Scholar] [CrossRef]
- Tkachenko, E.A.; Ishmukhametov, A.A.; Dzagurova, T.K.; Bernshtein, A.D.; Morozov, V.G.; Siniugina, A.A.; Kurashova, S.S.; Balkina, A.S.; Tkachenko, P.E.; Kruger, D.H.; et al. Hemorrhagic Fever with Renal Syndrome, Russia. Emerg Infect. Dis. 2019, 25, 2325–2328. [Google Scholar] [CrossRef]
- Tkachenko, E.A.; Dzagurova, T.K.; Tkachenko, P.E. Current Status of Hantavirus Vaccines Development. Nov. Technol. Vaccine Dev. 2014, 1, 113–151. [Google Scholar] [CrossRef]
- Casals, J.; Henderson, B.E.; Hoogstraal, H.; Johnson, K.M.; Shelokov, A. A Review of Soviet Viral Hemorrhagic Fevers, 1969. J. Infect. Dis. 1970, 122, 437–453. [Google Scholar] [CrossRef]
- Povalishina, T.P. On the Influence of the Human Activity upon the Existence and Detection of Natural Foci of Haemorrhagic Fever with a Renal Syndrome; Rasicky, B., Heyberger, K., Eds.; Theoretical Questions of Natural Foci of Diseases; Czechoslovak Academy of Sciences: Prague, Czech Republic, 1965. [Google Scholar]
- Johnson, K.M. Hantaviruses: History and Overview. Curr. Top. Microbiol. Immunol. 2001, 256, 1–14. [Google Scholar]
- Smadel, J.E. Epidemic Hemorrhagic Fever. Am. J. Public Health Nations Health 1953, 43, 1327–1330. [Google Scholar] [CrossRef] [Green Version]
- Myhrman, G. Nephropathia Epidemica a New Infectious Disease in Northern Scandinavia. Acta Med. Scand. 1951, 140, 52–56. [Google Scholar] [CrossRef]
- Smorodintsev, A.A.; Chudakov, V.G.; Churilov, A.V. Haemorrhagic Nephroso-Nephritis; Pergamon Press Ltd.: London, UK; New York, NY, USA; Paris, France; Los Angeles, CA, USA, 1959. [Google Scholar]
- Lokugamage, K.; Kariwa, H.; Hayasaka, D.; Cui, B.Z.; Iwasaki, T.; Lokugamage, N.; Ivanov, L.I.; Volkov, V.I.; Demenev, V.A.; Slonova, R.; et al. Genetic Characterization of Hantaviruses Transmitted by the Korean Field Mouse (Apodemus peninsulae), Far East Russia. Emerg. Infect. Dis. 2002, 8, 768–776. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the Etiologic Agent of Korean Hemorrhagic Fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef]
- McCormick, J.B.; Palmer, E.L.; Sasso, D.R.; Kiley, M.P. Morphological identification of the agent of korean haemorrhagic fever (Hantaan virus) as a member of the bunyaviridae. Lancet 1982, 319, 765–768. [Google Scholar] [CrossRef]
- Brummer-Korvenkontio, M.; Vaheri, A.; Hovi, T.; von Bonsdorff, C.-H.; Vuorimies, J.; Manni, T.; Penttinen, K.; Oker-Blom, N.; Lahdevirta, J. Nephropathia Epidemica: Detection of Antigen in Bank Voles and Serologic Diagnosis of Human Infection. J. Infect. Dis. 1980, 141, 131–134. [Google Scholar] [CrossRef]
- Plyusnin, A.; Elliott Richard, M. Bunyaviridae: Molecular and Cellular Biology; Caister Academic Press: Norwich, UK, 2011; ISBN 978-1-912530-52-6. [Google Scholar]
- Song, J.W.; Baek, L.J.; Schmaljohn, C.S.; Yanagihara, R. Thottapalayam Virus, a Prototype Shrewborne Hantavirus. Emerg. Infect. Dis. 2007, 13, 980. [Google Scholar] [CrossRef]
- Carey, D.E.; Reuben, R.; Panicker, K.N.; Shope, R.E.; Myers, R.M. Thottapalayam Virus: A Presumptive Arbovirus Isolated from a Shrew in India. Indian J. Med. Res. 1971, 59, 1758–1760. [Google Scholar]
- Clement, J.; Maes, P.; Muthusethupathi, M.; Nainan, G.; van Ranst, M. First Evidence of Fatal Hantavirus Nephropathy in India, Mimicking Leptospirosis. Nephrol. Dial. Transplant. 2006, 21, 826–827. [Google Scholar] [CrossRef] [Green Version]
- Chandy, S.; Mitra, S.; Sathish, N.; Vijayakumar, T.S.; Abraham, O.C.; Jesudason, M.V.; Abraham, P.; Yoshimatsu, K.; Arikawa, J.; Sridharan, G. A Pilot Study for Serological Evidence of Hantavirus Infection in Human Population in South India. Indian J. Med. Res. 2005, 122, 211–215. [Google Scholar]
- Chin, C.; Chiueh, T.-S.; Yang, W.-C.; Yang, T.-H.; Shih, C.-M.; Lin, H.-T.; Lin, K.-C.; Lien, J.-C.; Tsai, T.F.; Ruo, S.L.; et al. Hantavirus Infection in Taiwan: The Experience of a Geographically Unique Area. J. Med. Virol. 2000, 60, 237–247. [Google Scholar] [CrossRef]
- Elwell, M.; Ward, G. Serologic Evidence of Hantaan-like Virus in Rodents and Man in Thailand. Southeast Asian J. Trop. 1985, 16, 349–354. [Google Scholar]
- Song G Epidemiological Progresses of Hemorrhagic Fever with Renal Syndrome in China. Chin. Med. J. (Engl.) 1999, 112, 472–477.
- Jameson, L.J.; Taori, S.K.; Atkinson, B.; Levick, P.; Featherstone, C.A.; van der Burgt, G.; McCarthy, N.; Hart, J.; Osborne, J.C.; Walsh, A.L.; et al. Pet Rats as a Source of Hantavirus in England and Wales, 2013. Eurosurveillance 2013, 18, 20415. [Google Scholar] [CrossRef]
- Fang, L.Z.; Zhao, L.; Wen, H.L.; Zhang, Z.T.; Liu, J.W.; He, S.T.; Xue, Z.F.; Ma, D.Q.; Zhang, X.S.; Zhang, Y.; et al. Reservoir Host Expansion of Hantavirus, China. Emerg. Infect. Dis. 2015, 21, 170. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yoshimatsu, K.; Ebihara, H.; Ogino, M.; Araki, K.; Kariwa, H.; Wang, Z.; Luo, Z.; Li, D.; Hang, C.; et al. Genetic Diversity of Hantaviruses Isolated in China and Characterization of Novel Hantaviruses Isolated from Niviventer Confucianus and Rattus Rattus. Virology 2000, 278, 332–345. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.-Y.; Yang, W.-H.; Pan, H.; Zhou, J.-H.; Han, X.; Zhu, G.-J.; Desmond, J.S.; Daszak, P.; Shi, Z.-L.; Zhang, Y.-Z. Fugong Virus, a Novel Hantavirus Harbored by the Small Oriental Vole (Eothenomys eleusis) in China. Virol. J. 2016, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Wang, J.-B.; Gaowa, H.-S.; Yao, L.-S.; Hu, G.-W.; Li, M.-H.; Chen, H.-X.; Plyusnin, A.; Shao, R.; Zhang, Y.-Z. Isolation and Genetic Characterization of Hantaviruses Carried ByMicrotus Voles in China. J. Med. Virol. 2008, 80, 680–688. [Google Scholar] [CrossRef]
- Lin, X.-D.; Zhou, R.-H.; Fan, F.-N.; Ying, X.-H.; Sun, X.-Y.; Wang, W.; Holmes, E.C.; Zhang, Y.-Z. Biodiversity and Evolution of Imjin Virus and Thottapalayam Virus in Crocidurinae Shrews in Zhejiang Province, China. Virus Res. 2014, 189, 114–120. [Google Scholar] [CrossRef]
- Arai, S.; Gu, S.H.; Baek, L.J.; Tabara, K.; Bennett, S.N.; Oh, H.S.; Takada, N.; Kang, H.J.; Tanaka-Taya, K.; Morikawa, S.; et al. Divergent ancestral lineages of newfound hantaviruses harbored by phylogenetically related crocidurine shrew species in Korea. Virology 2012, 424, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.-F.; Zhang, W.-Y.; Wu, X.-M.; Zhang, P.-H.; Cao, W.-C. Soochong Virus and Amur Virus Might Be the Same Entities of Hantavirus. J. Med. Virol. 2007, 79, 1792–1795. [Google Scholar] [CrossRef]
- Plyusnina, A.; Ibrahim, I.N.; Plyusnin, A. A Newly Recognized Hantavirus in the Asian House Rat (Rattus tanezumi) in Indonesia. J. Gen. Virol. 2009, 90, 205–209. [Google Scholar] [CrossRef]
- Kutanan, W.; Kampuansai, J.; Brunelli, A.; Ghirotto, S.; Pittayaporn, P.; Ruangchai, S.; Schröder, R.; MacHoldt, E.; Srikummool, M.; Kangwanpong, D.; et al. New Insights from Thailand into the Maternal Genetic History of Mainland Southeast Asia. Eur. J. Hum. Genet. 2018, 26, 898–911. [Google Scholar] [CrossRef] [Green Version]
- Garanina, S.B.; Platonov, A.E.; Zhuravlev, V.I.; Murashkina, A.N.; Yakimenko, V.V.; Korneev, A.G.; Shipulin, G.A. Genetic Diversity and Geographic Distribution of Hantaviruses in Russia. Zoonoses Public Health 2009, 56, 297–309. [Google Scholar] [CrossRef]
- Abu Daud, N.H.; Kariwa, H.; Tanikawa, Y.; Nakamura, I.; Seto, T.; Miyashita, D.; Yoshii, K.; Nakauchi, M.; Yoshimatsu, K.; Arikawa, J.; et al. Mode of Infection of Hokkaido Virus (Genus hantavirus) among Grey Red-Backed Voles, Myodes Rufocanus, in Hokkaido, Japan. Microbiol. Immunol. 2007, 51, 1081–1090. [Google Scholar] [CrossRef]
- Song, K.J.; Baek, L.J.; Moon, S.; Ha, S.J.; Kim, S.H.; Park, K.S.; Klein, T.A.; Sames, W.; Kim, H.C.; Lee, J.S.; et al. Muju Virus, a Novel Hantavirus Harboured by the Arvicolid Rodent Myodes Regulus in Korea. J. Gen. Virol. 2007, 88, 3121. [Google Scholar] [CrossRef]
- Zhang, Y.; Yuan, J.; Yang, X.; Zhou, J.; Yang, W.; Peng, C.; Zhang, H.-L.; Shi, Z. A Novel Hantavirus Detected in Yunnan Red-Backed Vole (Eothenomys miletus) in China. J. Gen. Virol. 2011, 92, 1454–1457. [Google Scholar] [CrossRef]
- Song, J.-W.; Kang, H.J.; Song, K.-J.; Truong, T.T.; Bennett, S.N.; Arai, S.; Truong, N.U.; Yanagihara, R. Newfound Hantavirus in Chinese Mole Shrew, Vietnam. Emerg. Infect. Dis. 2007, 13, 1784–1787. [Google Scholar] [CrossRef]
- Guo, W.P.; Lin, X.D.; Wang, W.; Tian, J.H.; Cong, M.L.; Zhang, H.L.; Wang, M.R.; Zhou, R.H.; Wang, J.B.; Li, M.H.; et al. Phylogeny and Origins of Hantaviruses Harbored by Bats, Insectivores, and Rodents. PLoS Pathog. 2013, 9, e1003159. [Google Scholar] [CrossRef] [Green Version]
- Arai, S.; Ohdachi, S.D.; Asakawa, M.; Kang, H.J.; Mocz, G.; Arikawa, J.; Okabe, N.; Yanagihara, R. Molecular Phylogeny of a Newfound Hantavirus in the Japanese Shrew Mole (Urotrichus talpoides). Proc. Natl. Acad. Sci. USA 2008, 105, 16296–16301. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Q.; Gao, J.-H.; Li, M.; Guo, W.-P.; Lu, M.-Q.; Wang, W.; Hu, M.-X.; Li, M.-H.; Yang, J.; Liang, H.-J.; et al. Co-Circulation of Hantaan, Kenkeme, and Khabarovsk Hantaviruses in Bolshoy Ussuriysky Island, China. Virus Res. 2014, 191, 51–58. [Google Scholar] [CrossRef]
- Krautkrämer, E.; Zeier, M. Old World Hantaviruses: Aspects of Pathogenesis and Clinical Course of Acute Renal Failure. Virus Res. 2014, 187, 59–64. [Google Scholar] [CrossRef]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A Global Perspective on Hantavirus Ecology, Epidemiology, and Disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Du, H.; Wang, L.M.; Wang, P.Z.; Bai, X.F. Hemorrhagic Fever with Renal Syndrome: Pathogenesis and Clinical Picture. Front. Cell. Infect. Microbiol. 2016, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Guang, M.Y.; Liu, G.Z.; Cosgriff, T.M. Hemorrhage in Hemorrhagic Fever with Renal Syndrome in China. Clin. Infect. Dis. 1989, 11, S884–S890. [Google Scholar] [CrossRef]
- Krüger, D.H.; Ulrich, R.; Lundkvist, Å. Hantavirus Infections and Their Prevention. Microbes Infect. 2001, 3, 1129–1144. [Google Scholar] [CrossRef]
- Vaheri, A.; Henttonen, H.; Voutilainen, L.; Mustonen, J.; Sironen, T.; Vapalahti, O. Hantavirus Infections in Europe and Their Impact on Public Health. Rev. Med. Virol. 2012, 23, 35–49. [Google Scholar] [CrossRef]
- Wang, M.; Wang, J.; Wang, T.; Li, J.; Hui, L.; Ha, X. Thrombocytopenia as a Predictor of Severe Acute Kidney Injury in Patients with Hantaan Virus Infections. PLoS ONE 2013, 8, e53236. [Google Scholar] [CrossRef]
- Linderholm, M.; Elgh, F. Clinical Characteristics of Hantavirus Infections on the Eurasian Continent. Curr. Top. Microbiol. Immunol. 2000, 256, 135–151. [Google Scholar] [CrossRef]
- Shastri, B.; Kofman, A.; Hennenfent, A.; Klena, J.D.; Nicol, S.; Graziano, J.C.; Morales-Betoulle, M.; Cannon, D.; Maradiaga, A.; Tran, A.; et al. Domestically Acquired Seoul Virus Causing Hemophagocytic Lymphohistiocytosis—Washington, DC, 2018. In Open Forum Infectious Diseases 2019; Oxford University Press: New York, NY, USA, 2019; Volume 6. [Google Scholar] [CrossRef] [Green Version]
- Bergstedt Oscarsson, K.; Brorstad, A.; Baudin, M.; Lindberg, A.; Forssén, A.; Evander, M.; Eriksson, M.; Ahlm, C. Human Puumala Hantavirus Infection in Northern Sweden; Increased Seroprevalence and Association to Risk and Health Factors. BMC Infect. Dis. 2016, 16, 566. [Google Scholar] [CrossRef] [Green Version]
- Laine, O.; Mäkelä, S.; Mustonen, J.; Helminen, M.; Vaheri, A.; Lassila, R.; Joutsi-Korhonen, L. Platelet Ligands and ADAMTS13 during Puumala Hantavirus Infection and Associated Thrombocytopenia. Blood Coagul. Fibrinolysis 2011, 22, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Dheerasekara, K.; Sumathipala, S.; Muthugala, R. Hantavirus Infections—Treatment and Prevention. Curr. Treat. Options Infect. Dis. 2020, 12, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Hägele, S.; Nusshag, C.; Müller, A.; Baumann, A.; Zeier, M.; Krautkrämer, E. Cells of the Human Respiratory Tract Support the Replication of Pathogenic Old World Orthohantavirus Puumala. Virol. J. 2021, 18, 169. [Google Scholar] [CrossRef] [PubMed]
- Galeno, H.; Mora, J.; Villagra, E.; Fernandez, J.; Hernandez, J.; Mertz, G.J.; Ramirez, E. First Human Isolate of Hantavirus (Andes virus) in the Americas. Emerg. Infect. Dis. 2002, 8, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Rasmuson, J.; Andersson, C.; Norrman, E.; Haney, M.; Evander, M.; Ahlm, C. Time to Revise the Paradigm of Hantavirus Syndromes? Hantavirus Pulmonary Syndrome Caused by European Hantavirus. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 30, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Ferrés, M.; Vial, P.; Marco, C.; Yañez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.J.; Mertz, G.J. Prospective Evaluation of Household Contacts of Persons with Hantavirus Cardiopulmonary Syndrome in Chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef] [Green Version]
- Pensiero, M.N.; Sharefkin, J.B.; Dieffenbach, C.W.; Hay’, J. Hantaan Virus Infection of Human Endothelial Cells. J. Virol. 1992, 66, 5929–5936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanagihara, R.; Silverman, D.J. Experimental Infection of Human Vascular Endothelial Cells by Pathogenic and Nonpathogenic Hantaviruses. Arch. Virol. 1990, 111, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Antoine, M.; Langlois, M.E.; Bres, E.; Rabeyrin, M.; Reynes, J.M.; Deeb, A. Imported Haemorrhagic Fever with Renal Syndrome Caused by Dobrava-Belgrade Hantavirus in France. Clin. Kidney J. 2021, 14, 1014–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaheri, A.; Strandin, T.; Hepojoki, J.; Sironen, T.; Henttonen, H.; Mäkelä, S.; Mustonen, J. Uncovering the Mysteries of Hantavirus Infections. Nat. Rev. Microbiol. 2013, 11, 539–550. [Google Scholar] [CrossRef]
- Romero, M.G.; Anjum, F. Hemorrhagic Fever Renal Syndrome; StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2020. Available online: https://www.ncbi.nlm.nih.gov/books/NBK560660/ (accessed on 1 January 2022).
- Yi, J.; Xu, Z.; Zhuang, R.; Wang, J.; Zhang, Y.; Ma, Y.; Liu, B.; Zhang, Y.; Zhang, C.; Yan, G.; et al. Hantaan Virus RNA Load in Patients Having Hemorrhagic Fever with Renal Syndrome: Correlation with Disease Severity. J. Infect. Dis. 2013, 207, 1457–1461. [Google Scholar] [CrossRef] [Green Version]
- Saksida, A.; Duh, D.; Korva, M.; Avsic-Zupanc, T. Dobrava Virus RNA Load in Patients Who Have Hemorrhagic Fever with Renal Syndrome. J. Infect. Dis. 2008, 197, 681–685. [Google Scholar] [CrossRef] [Green Version]
- Avšič-Županc, T.; Saksida, A.; Korva, M. Hantavirus Infections. Clin. Microbiol. Infect. 2019, 21, e6–e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosgriff, T.M. Mechanisms of Disease in Hantavirus Infection: Pathophysiology of Hemorrhagic Fever with Renal Syndrome. Rev. Infect. Dis. 1990, 13, 97–107. [Google Scholar] [CrossRef]
- Ferluga, D.; Vizjak, A. Hantavirus Nephropathy. J. Am. Soc. Nephrol. 2008, 19, 1653–1658. [Google Scholar] [CrossRef] [Green Version]
- Gavrilovskaya, I.N.; Peresleni, T.; Geimonen, E.; Mackow, E.R. Pathogenic Hantaviruses Selectively Inhibit Β3 Integrin Directed Endothelial Cell Migration. Arch. Virol. 2002, 147, 1913–1931. [Google Scholar] [CrossRef] [PubMed]
- Leavesley, D.I.; Schwartz, M.A.; Rosenfeld, M.; Cheresh, D.A. Integrin Β1- and Β3-Mediated Endothelial Cell Migration Is Triggered through Distinct Signaling Mechanisms. J. Cell Biol. 1993, 121, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Gavrilovskaya, I.N.; Brown, E.J.; Ginsberg, M.H.; Mackow, E.R. Cellular Entry of Hantaviruses Which Cause Hemorrhagic Fever with Renal Syndrome Is Mediated by β 3 Integrins. J. Virol. 1999, 73, 3951–3959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Kang, E.T.; Kim, Y.G.; Han, J.S.; Lee, J.S.; Kim, Y.I.; Hall, W.C.; Dalrymple, J.M.; Peters, C.J. Localization of Hantaan Viral Envelope Glycoproteins by Monoclonal Antibodies in Renal Tissues from Patients with Korean Hemorrhagic Fever H. Am. J. Clin. Pathol. 1993, 100, 398–403. [Google Scholar] [CrossRef] [Green Version]
- Gorbunova, E.; Gavrilovskaya, I.N.; Mackow, E.R. Pathogenic Hantaviruses Andes Virus and Hantaan Virus Induce Adherens Junction Disassembly by Directing Vascular Endothelial Cadherin Internalization in Human Endothelial Cells. J. Virol. 2010, 84, 7405–7411. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhang, Y.; Li, Y.; Pan, L.; Bai, L.; Zhuang, Y.; Huang, C.X.; Wang, J.P.; Yu, H.T.; Wei, X.; et al. Dysregulation of the Β3 Integrin-VEGFR2 Complex in Hantaan Virus-Directed Hyperpermeability upon Treatment with VEGF. Arch. Virol. 2012, 157, 1051–1061. [Google Scholar] [CrossRef] [PubMed]
- Furuse, M.; Fujita, K.; Hiiragi, T.; Fujimoto, K.; Tsukita, S. Claudin-1 and -2: Novel Integral Membrane Proteins Localizing at Tight Junctions with No Sequence Similarity to Occludin. J. Cell Biol. 1998, 141, 1539–1550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Wang, W.; Wang, J.P.; Pan, L.; Zhang, Y.; Yu, H.T.; Jiang, W.; Wang, P.Z.; Bai, X.F. Elevated Vascular Endothelial Growth Factor Levels Induce Hyperpermeability of Endothelial Cells in Hantavirus Infection. J. Int. Med. Res. 2012, 40, 1812–1821. [Google Scholar] [CrossRef]
- Krautkramer, E.; Grouls, S.; Stein, N.; Reiser, J.; Zeier, M. Pathogenic Old World Hantaviruses Infect Renal Glomerular and Tubular Cells and Induce Disassembling of Cell-to-Cell Contacts. J. Virol. 2011, 85, 9811–9823. [Google Scholar] [CrossRef] [Green Version]
- Dieterle, M.E.; Solà-Riera, C.; Ye, C.; Goodfellow, S.M.; Mittler, E.; Kasikci, E.; Bradfute, S.B.; Klingström, J.; Jangra, R.K.; Chandran, K. Genetic Depletion Studies Inform Receptor Usage by Virulent Hantaviruses in Human Endothelial Cells. Elife 2021, 10, e69708. [Google Scholar] [CrossRef]
- Jangra, R.K.; Herbert, A.S.; Li, R.; Jae, L.T.; Kleinfelter, L.M.; Slough, M.M.; Barker, S.L.; Guardado-Calvo, P.; Román-Sosa, G.; Dieterle, M.E.; et al. Protocadherin-1 Is Essential for Cell Entry by New World Hantaviruses. Nature 2018, 563, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Connolly-Andersen, A.M.; Thunberg, T.; Ahlm, C. Endothelial Activation and Repair during Hantavirus Infection: Association with Disease Outcome. In Open Forum Infectious Diseases; Oxford University Press: New York, NY, USA, 2014; Volume 1. [Google Scholar] [CrossRef] [Green Version]
- Nusshag, C.; Osberghaus, A.; Baumann, A.; Schnitzler, P.; Zeier, M.; Krautkrämer, E. Deregulation of Levels of Angiopoietin-1 and Angiopoietin-2 Is Associated with Severe Courses of Hantavirus Infection. J. Clin. Virol. 2017, 94, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.R.; Hwang, J.Y.; Park, H.S. Hantaan Virus Reduces the von Willebrand Factor in Human Umbilical Vein Endothelial Cells. J. Bacteriol. Virol. 2007, 37, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Khaiboullina, S.F.; Morzunov, S.P.; St. Jeor, S.C.; Rizvanov, A.A.; Lombardi, V.C. Hantavirus Infection Suppresses Thrombospondin-1 Expression in Cultured Endothelial Cells in a Strain-Specific Manner. Front. Microbiol. 2016, 7, 1077. [Google Scholar] [CrossRef] [Green Version]
- McPherson, J.; Sage, H.; Bornstein, P. Isolation and Characterization of a Glycoprotein Secreted by Aortic Endothelial Cells in Culture. Apparent Identity with Platelet Thrombospondin. J. Biol. Chem. 1981, 256, 11330–11336. [Google Scholar] [CrossRef]
- Silverstein, R.L.; Harpel, P.C.; Nachman, R.L. Tissue Plasminogen Activator and Urokinase Enhance the Binding of Plasminogen to Thrombospondin. J. Biol. Chem. 1986, 261, 9959–9965. [Google Scholar] [CrossRef] [PubMed]
- Stockschlaeder, M.; Schneppenheim, R.; Budde, U. Update on von Willebrand Factor Multimers: Focus on High-Molecular-Weight Multimers and Their Role in Hemostasis. Blood Coagul. Fibrinolysis 2014, 25, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Outinen, T.K.; Laine, O.K.; Mäkelä, S.; Pörsti, I.; Huhtala, H.; Vaheri, A.; Mustonen, J. Thrombocytopenia Associates with the Severity of Inflammation and Variables Reflecting Capillary Leakage in Puumala Hantavirus Infection, an Analysis of 546 Finnish Patients. Infect. Dis. 2016, 48, 682–687. [Google Scholar] [CrossRef]
- Koskela, S.; Mäkelä, S.; Strandin, T.; Vaheri, A.; Outinen, T.; Joutsi-Korhonen, L.; Pörsti, I.; Mustonen, J.; Laine, O. Coagulopathy in Acute Puumala Hantavirus Infection. Viruses 2021, 13, 1553. [Google Scholar] [CrossRef]
- Laine, O.; Joutsi-Korhonen, L.; Lassila, R.; Koski, T.; Huhtala, H.; Vaheri, A.; Mäkelä, S.; Mustonen, J. Hantavirus Infection-Induced Thrombocytopenia Triggers Increased Production but Associates with Impaired Aggregation of Platelets except for Collagen. Thromb. Res. 2015, 136, 1126–1132. [Google Scholar] [CrossRef]
- Lee, M.; Kim, B.K.; Suhnggwon; Park, S.; Han, J.Ŝ.; Kim, S.T.; Lee, J.S. Coagulopathy in Hemorrhagic Fever with Renal Syndrome (Korean Hemorrhagic Fever). Rev. Infect. Dis. 1989, 11, S877–S883. [Google Scholar] [CrossRef] [PubMed]
- Rasche, F.M.; Uhel, B.; Ulrich, R.; Krüger, D.H.; Karges, W.; Czock, D.; Hampl, W.; Keller, F.; Meisel, H.; Von Müller, L. Thrombocytopenia and Acute Renal Failure in Puumala Hantavirus Infections. Emerg. Infect. Dis. 2004, 10, 1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laine, O.; Mäkelä, S.; Mustonen, J.; Huhtala, H.; Szanto, T.; Vaheri, A.; Lassila, R.; Joutsi-Korhonen, L. Enhanced Thrombin Formation and Fibrinolysis during Acute Puumala Hantavirus Infection. Thromb. Res. 2010, 126, 154–158. [Google Scholar] [CrossRef]
- Sundberg, E.; Hultdin, J.; Nilsson, S.; Ahlm, C. Evidence of Disseminated Intravascular Coagulation in a Hemorrhagic Fever with Renal Syndrome—Scoring Models and Severe Illness. PLoS ONE 2011, 6, e21134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, S.; Wada, H.; Abe, Y.; Yamada, E.; Sakaguchi, A.; Nishioka, J.; Hatada, T.; Ishikura, K.; Yamada, N.; Sudo, A.; et al. Elevated Levels of Prothrombin Fragment 1 + 2 Indicate High Risk of Thrombosis. Clin. Appl. Thromb./Hemost. 2008, 14, 279–285. [Google Scholar] [CrossRef]
- Connolly-Andersen, A.M.; Whitaker, H.; Klingström, J.; Ahlm, C. Risk of Venous Thromboembolism Following Hemorrhagic Fever with Renal Syndrome: A Self-Controlled Case Series Study. Clin. Infect. Dis. 2018, 66, 268–273. [Google Scholar] [CrossRef] [Green Version]
- De Caterina, R.; Husted, S.; Wallentin, L.; Andreotti, F.; Arnesen, H.; Bachmann, F.; Baigent, C.; Huber, K.; Jespersen, J.; Kristensen, S.D.; et al. General Mechanisms of Coagulation and Targets of Anticoagulants (Section I): Position Paper of the ESC Working Group on Thrombosis—Task Force on Anticoagulants in Heart Disease. Thromb. Haemost. 2013, 109, 569–579. [Google Scholar] [CrossRef]
- Kohler, H.P.; Grant, P.J. Plasminogen-Activator Inhibitor Type 1 and Coronary Artery Disease. N. Engl. J. Med. 2000, 342, 1792–1801. [Google Scholar] [CrossRef]
- Van Hinsbergh, V.W.M. Endothelium—Role in Regulation of Coagulation and Inflammation. Semin. Immunopathol. 2012, 34, 93. [Google Scholar] [CrossRef] [Green Version]
- Yau, J.W.; Teoh, H.; Verma, S. Endothelial Cell Control of Thrombosis. BMC Cardiovasc. Disord 2015, 15, 130. [Google Scholar] [CrossRef] [Green Version]
- Kaneto, T.; Fujii, S.; Matsumoto, A.; Goto, D.; Makita, N.; Hamada, J.; Moriuchi, T.; Kitabatake, A. Induction of Tissue Factor Expression in Endothelial Cells by Basic Fibroblast Growth Factor and Its Modulation by Fenofibric Acid. Thromb. J. 2003, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Goeijenbier, M.; Meijers, J.C.M.; Anfasa, F.; Roose, J.M.; van de Weg, C.A.M.; Bakhtiari, K.; Henttonen, H.; Vaheri, A.; Osterhaus, A.D.M.E.; van Gorp, E.C.M.; et al. Effect of Puumala Hantavirus Infection on Human Umbilical Vein Endothelial Cell Hemostatic Function: Platelet Interactions, Increased Tissue Factor Expression and Fibrinolysis Regulator Release. Front. Microbiol. 2015, 6, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmedes, C.M.; Grover, S.P.; Hisada, Y.M.; Goeijenbier, M.; Hultdin, J.; Nilsson, S.; Thunberg, T.; Ahlm, C.; MacKman, N.; Connolly, A.M.F. Circulating Extracellular Vesicle Tissue Factor Activity during Orthohantavirus Infection Is Associated with Intravascular Coagulation. J. Infect. Dis. 2020, 222, 1392–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, A.P.; Mackman, N. Tissue Factor and Thrombosis: The Clot Starts Here. Thromb. Haemost. 2010, 104, 432–439. [Google Scholar] [CrossRef]
- Göbel, K.; Eichler, S.; Wiendl, H.; Chavakis, T.; Kleinschnitz, C.; Meuth, S.G. The Coagulation Factors Fibrinogen, Thrombin, and Factor XII in Inflammatory Disorders—A Systematic Review. Front. Immunol. 2018, 9, 1731. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhang, C.; Zhuang, R.; Ma, Y.; Zhang, Y.; Yi, J.; Yang, A.; Jin, B. IL-33/ST2 Correlates with Severity of Haemorrhagic Fever with Renal Syndrome and Regulates the Inflammatory Response in Hantaan Virus-Infected Endothelial Cells. PLoS Negl. Trop. Dis. 2015, 9, e0003514. [Google Scholar] [CrossRef] [PubMed]
- Sundstrom, J.B.; McMullan, L.K.; Spiropoulou, C.F.; Hooper, W.C.; Ansari, A.A.; Peters, C.J.; Rollin, P.E. Hantavirus Infection Induces the Expression of RANTES and IP-10 without Causing Increased Permeability in Human Lung Microvascular Endothelial Cells. J. Virol. 2001, 75, 6070–6085. [Google Scholar] [CrossRef] [Green Version]
- Khaiboullina, S.F.; Netski, D.M.; Krumpe, P.; St Jeor, S.C. Effects of Tumor Necrosis Factor Alpha on Sin Nombre Virus Infection In Vitro. J. Virol. 2000, 74, 11966–11971. [Google Scholar] [CrossRef] [Green Version]
- Raftery, M.J.; Kraus, A.A.; Ulrich, R.; Krüger, D.H.; Schönrich, G. Hantavirus Infection of Dendritic Cells. J. Virol. 2002, 76, 10724–10733. [Google Scholar] [CrossRef] [Green Version]
- Nagai, T.; Tanishita, O.; Takahashi, Y.; Yamanouchi, T.; Domae, K.; Kondo, K.; Dantas, J.R.; Yamanishi, K. Isolation of Haemorrhagic Fever with Renal Syndrome Virus from Leukocytes of Rats and Virus Replication in Cultures of Rat and Human Macrophages. J. Gen. Virol. 1985, 66, 1271–1278. [Google Scholar] [CrossRef]
- Nourshargh, S.; Alon, R. Leukocyte Migration into Inflamed Tissues. Immunity 2014, 41, 694–707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martynova, E.; Davidyuk, Y.; Kabwe, E.; Garanina, E.E.; Shakirova, V.; Pavelkina, V.; Uskova, Y.; Stott, R.J.; Foster, T.L.; Markelova, M.; et al. Cytokine, Chemokine, and Metalloprotease Activation in the Serum of Patients with Nephropathia Epidemica from the Republic of Tatarstan and the Republic of Mordovia, Russia. Pathogens 2021, 10, 527. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, M.; Eckerle, I.; Daniel, V.; Burkhardt, U.; Opelz, G.; Schnitzler, P. Cytokine Expression during Early and Late Phase of Acute Puumala Hantavirus Infection. BMC Immunol. 2011, 12, 65. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Guo, X.; Wang, Y.; Tian, F.; Luo, W.; Zou, Y. Cytokine Response to Hantaan Virus Infection in Patients with Hemorrhagic Fever with Renal Syndrome. J. Med. Virol. 2017, 89, 1139–1145. [Google Scholar] [CrossRef]
- Khaiboullina, S.F.; Levis, S.; Morzunov, S.P.; Martynova, E.V.; Anokhin, V.A.; Gusev, O.A.; St Jeor, S.C.; Lombardi, V.C.; Rizvanov, A.A. Serum Cytokine Profiles Differentiating Hemorrhagic Fever with Renal Syndrome and Hantavirus Pulmonary Syndrome. Front. Immunol. 2017, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Brebner, K.; Hayley, S.; Zacharko, R.; Merali, Z.; Anisman, H. Synergistic Effects of Interleukin-1β, Interleukin-6, and Tumor Necrosis Factor-α: Central Monoamine, Corticosterone, and Behavioral Variations. Neuropsychopharmacology 2000, 22, 566–580. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Kullberg, B.J.; Van der Meer, J.W.M. Circulating Cytokines as Mediators of Fever. Clin. Infect. Dis. 2000, 31, S178–S184. [Google Scholar] [CrossRef] [Green Version]
- Sundgren-Andersson, A.K.; Ostlund, P.; Bartfai, T. IL-6 Is Essential in TNF-α-Induced Fever. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, R2028–R2034. [Google Scholar] [CrossRef] [PubMed]
- Puhlmann, M.; Weinreich, D.M.; Farma, J.M.; Carroll, N.M.; Turner, E.M.; Alexander, H.R. Interleukin-1β Induced Vascular Permeability Is Dependent on Induction of Endothelial Tissue Factor (TF) Activity. J. Transl. Med. 2005, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Sawant, D.A.; Tharakan, B.; Wilson, R.L.; Stagg, H.W.; Hunter, F.A.; Childs, E.W. Regulation of TNF-α-Induced Microvascular Endothelial Cell Hyperpermeability by Recombinant Bcl-XL. J. Surg. Res. 2013, 184, 628. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.L.; Jiang, Z.Y.; Dong, J.; Liu, X.C.; Cai, S.Y.; Xiao, R.; Lu, Y.R. Expression of Tissue Factor Induced by IL-6 in HUVEC. Sichuan Da Xue Xue Bao Yi Xue Ban 2006, 37, 234–237. [Google Scholar] [PubMed]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Klingström, J.; Plyusnin, A.; Vaheri, A.; Lundkvist, Å. Wild-Type Puumala Hantavirus Infection Induces Cytokines, C-Reactive Protein, Creatinine, and Nitric Oxide in Cynomolgus Macaques. J. Virol. 2002, 76, 444–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.Y.; Song, C.H.; Choi, S.O. Acute Polyarthritis Associated with Hantavirus Infection. Nephrol. Dial. Transplant. 1999, 14, 2204–2205. [Google Scholar] [CrossRef]
- Martin-Sanchez, F.; Diamond, C.; Zeitler, M.; Gomez, A.I.; Baroja-Mazo, A.; Bagnall, J.; Spiller, D.; White, M.; Daniels, M.J.D.; Mortellaro, A.; et al. Inflammasome-Dependent IL-1β Release Depends upon Membrane Permeabilisation. Cell Death Differ. 2016, 23, 1219–1231. [Google Scholar] [CrossRef] [Green Version]
- Hatscher, L.; Lehmann, C.H.K.; Purbojo, A.; Onderka, C.; Liang, C.; Hartmann, A.; Cesnjevar, R.; Bruns, H.; Gross, O.; Nimmerjahn, F.; et al. Select Hyperactivating NLRP3 Ligands Enhance the TH1-and TH17-Inducing Potential of Human Type 2 Conventional Dendritic Cells. Sci. Signal. 2021, 14, eabe1757. [Google Scholar] [CrossRef]
- Tominaga, K.; Yoshimoto, T.; Torigoe, K.; Kurimoto, M.; Matsui, K.; Hada, T.; Okamura, H.; Nakanishi, K. IL-12 Synergizes with IL-18 or IL-1β for IFN-γ Production from Human T Cells. Int. Immunol. 2000, 12, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; Levy, D.E.; Leonard, W.J.; Littman, D.R. IL-6 Programs TH-17 Cell Differentiation by Promoting Sequential Engagement of the IL-21 and IL-23 Pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef]
- Qin, H.; Wang, L.; Feng, T.; Elson, C.O.; Niyongere, S.A.; Lee, S.J.; Reynolds, S.L.; Weaver, C.T.; Roarty, K.; Serra, R.; et al. TGF-β Promotes Th17 Cell Development through Inhibition of SOCS3. J. Immunol. 2009, 183, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Friederichs, K.; Schmitz, J.; Weissenbach, M.; Heinrich, P.C.; Schaper, F. Interleukin-6-Induced Proliferation of Pre-B Cells Mediated by Receptor Complexes Lacking the SHP2/SOCS3 Recruitment Sites Revisited. Eur. J. Biochem. 2001, 268, 6401–6407. [Google Scholar] [CrossRef]
- Kaser, A.; Brandacher, G.; Steurer, W.; Kaser, S.; Offner, F.A.; Zoller, H.; Theurl, I.; Widder, W.; Molnar, C.; Ludwiczek, O.; et al. Interleukin-6 Stimulates Thrombopoiesis through Thrombopoietin: Role in Inflammatory Thrombocytosis. Blood 2001, 98, 2720–2725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Cao, S.; Zhang, Q.; Li, J.; Zhang, S.; Wu, W.; Qu, J.; Li, C.; Liang, M.; Li, D. Comparison of Serological Assays to Titrate Hantaan and Seoul Hantavirus-Specific Antibodies. Virol. J. 2017, 14, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connolly-Andersen, A.M.; Sundberg, E.; Ahlm, C.; Hultdin, J.; Baudin, M.; Larsson, J.; Dunne, E.; Kenny, D.; Lindahl, T.L.; Ramstrom, S.; et al. Increased Thrombopoiesis and Platelet Activation in Hantavirus-Infected Patients. J. Infect. Dis. 2015, 212, 1061–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saksida, A.; Wraber, B.; Avšič-Županc, T. Serum Levels of Inflammatory and Regulatory Cytokines in Patients with Hemorrhagic Fever with Renal Syndrome. BMC Infect. Dis. 2011, 11, 142. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.Z.; Li, Z.D.; Yu, H.T.; Zhang, Y.; Wang, W.; Jiang, W.; Bai, X.F. Elevated Serum Concentrations of Inflammatory Cytokines and Chemokines in Patients with Haemorrhagic Fever with Renal Syndrome. J. Int. Med. Res. 2012, 40, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Sokol, C.L.; Luster, A.D. The Chemokine System in Innate Immunity. Cold Spring Harb. Perspect. Biol. 2015, 7, a016303. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, A.C.; Barsante, M.M.; Esteves Arantes, R.M.; Bernard, C.C.A.; Teixeira, M.M.; Carvalho-Tavares, J. CCL2 and CCL5 Mediate Leukocyte Adhesion in Experimental Autoimmune Encephalomyelitis—An Intravital Microscopy Study. J. Neuroimmunol. 2005, 162, 122–129. [Google Scholar] [CrossRef]
- Groom, J.R.; Richmond, J.; Murooka, T.T.; Sorensen, E.W.; Sung, J.H.; Bankert, K.; von Andrian, U.H.; Moon, J.J.; Mempel, T.R.; Luster, A.D. CXCR3 Chemokine Receptor-Ligand Interactions in the Lymph Node Optimize CD4+ T Helper 1 Cell Differentiation. Immunity 2012, 37, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Thapa, M.; Welner, R.S.; Pelayo, R.; Carr, D.J.J. CXCL9 and CXCL10 Expression Are Critical for Control of Genital Herpes Simplex Virus Type 2 Infection through Mobilization of HSV-Specific CTL and NK Cells to the Nervous System. J. Immunol. 2008, 180, 1098–1106. [Google Scholar] [CrossRef] [Green Version]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of Natural Killer Cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Schmitz, J.E.; Kuroda, M.J.; Santra, S.; Sasseville, V.G.; Simon, M.A.; Lifton, M.A.; Racz, P.; Tenner-Racz, K.; Dalesandro, M.; Scallon, B.J.; et al. Control of Viremia in Simian Immunodeficiency Virus Infection by CD8+ Lymphocytes. Science 1999, 283, 857–860. [Google Scholar] [CrossRef] [Green Version]
- Schuch, A.; Hoh, A.; Thimme, R. The Role of Natural Killer Cells and CD8+ T Cells in Hepatitis B Virus Infection. Front. Immunol. 2014, 5, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Epps, H.L.; Schmaljohn, C.S.; Ennis, F.A. Human Memory Cytotoxic T-Lymphocyte (CTL) Responses to Hantaan Virus Infection: Identification of Virus-Specific and Cross-Reactive CD8+ CTL Epitopes on Nucleocapsid Protein. J. Virol. 1999, 73, 5301–5308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Epps, H.L.; Terajima, M.; Mustonen, J.; Arstila, T.P.; Corey, E.A.; Vaheri, A.; Ennis, F.A. Long-Lived Memory T Lymphocyte Responses after Hantavirus Infection. J. Exp. Med. 2002, 196, 579–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmuson, J.; Pourazar, J.; Mohamed, N.; Lejon, K.; Evander, M.; Blomberg, A.; Ahlm, C. Cytotoxic Immune Responses in the Lungs Correlate to Disease Severity in Patients with Hantavirus Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 713–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temonen, M.; Mustonen, J.; Helin, H.; Pasternack, A.; Vaheri, A.; Holthöfer, H. Cytokines, Adhesion Molecules, and Cellular Infiltration in Nephropathia Epidemica Kidneys: An Immunohistochemical Study. Clin. Immunol. Immunopathol. 1996, 78, 47–55. [Google Scholar] [CrossRef]
- Wang, M.; Wang, J.; Zhu, Y.; Xu, Z.; Yang, K.; Yang, A.; Jin, B. Cellular Immune Response to Hantaan Virus Nucleocapsid Protein in the Acute Phase of Hemorrhagic Fever with Renal Syndrome: Correlation with Disease Severity. J. Infect. Dis. 2009, 199, 188–195. [Google Scholar] [CrossRef]
- Tang, K.; Cheng, L.; Zhang, C.; Zhang, Y.; Zheng, X.; Zhang, Y.; Zhuang, R.; Jin, B.; Zhang, F.; Ma, Y. Novel Identified HLA-A*0201-Restricted Hantaan Virus Glycoprotein Cytotoxic T-Cell Epitopes Could Effectively Induce Protective Responses in HLA-A2.1/Kb Transgenic Mice May Associate with the Severity of Hemorrhagic Fever with Renal Syndrome. Front. Immunol. 2017, 8, 1797. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, T.; Ahlm, C.; Mohamed, N.; Evander, M.; Ljunggren, H.-G.; Björkström, N.K. Longitudinal Analysis of the Human T Cell Response during Acute Hantavirus Infection. J. Virol. 2011, 85, 10252–10260. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention Technical/Clinical Information: HPS Treatment—Hantavirus. Available online: https://www.cdc.gov/hantavirus/technical/hps/treatment.html (accessed on 12 July 2022).
- Centers for Disease Control and Prevention Facts about Hantaviruses. What You Need to Know to Prevent the Disease Pulmonary Syndrome (HPS). Available online: https://www.cdc.gov/hantavirus/pdf/hps_brochure.pdf (accessed on 23 August 2022).
- Zhang, Y.Z.; Xiao, D.L.; Wang, Y.; Wang, H.X.; Sun, L.; Tao, X.X.; Qu, Y.G. The Epidemic Characteristics and Preventive Measures of Hemorrhagic Fever with Syndromes in China. Zhonghua Liu Xing Bing Xue Za Zhi 2004, 25, 466–469. [Google Scholar]
- PAHO/WHO Hantavirus. Available online: https://www3.paho.org/hq/index.php?option=com_content&view=article&id=14911:hantavirus&Itemid=40721&lang=en#:~:text=Hantavirus%20(HV)%20is%20an%20emerging,of%20respiratory%20distress%20and%20hypotension (accessed on 13 October 2022).
- Centers for Disease Control and Prevention Cleaning Up after Rodents|Rodents|CDC. Available online: https://www.cdc.gov/rodents/prevent_infestations/clean_up.html#:~:text=Clean%20up%20rodent%20urine%20and,or%20droppings%20and%20cleaning%20product (accessed on 15 September 2022).
- Tian, H.; Stenseth, N.C. The Ecological Dynamics of Hantavirus Diseases: From Environmental Variability to Disease Prevention Largely Based on Data from China. PLoS Negl. Trop. Dis. 2019, 13, e0006901. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; Kim, W.K.; Lee, S.H.; Kim, J.; Lee, J.; Cho, S.; Lee, G.Y.; Noid, J.S.; Lee, K.H.; Song, J.W. A Novel Genotype of Hantaan Orthohantavirus Harbored by Apodemus Agrarius Chejuensis as a Potential Etiologic Agent of Hemorrhagic Fever with Renal Syndrome in Republic of Korea. PLoS Negl. Trop. Dis. 2021, 15, e0009400. [Google Scholar] [CrossRef]
- Ling, J.; Verner-Carlsson, J.; Eriksson, P.; Plyusnina, A.; Löhmus, M.; Järhult, J.D.; van de Goot, F.; Plyusnin, A.; Lundkvist, Å.; Sironen, T. Genetic Analyses of Seoul Hantavirus Genome Recovered from Rats (Rattus norvegicus) in the Netherlands Unveils Diverse Routes of Spread into Europe. J. Med. Virol. 2019, 91, 724–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynes, J.M.; Razafindralambo, N.K.; Lacoste, V.; Olive, M.M.; Barivelo, T.A.; Soarimalala, V.; Heraud, J.M.; Lavergne, A. Anjozorobe Hantavirus, a New Genetic Variant of Thailand Virus Detected in Rodents from Madagascar. Vector-Borne Zoonotic Dis. 2014, 14, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raharinosy, V.; Olive, M.M.; Andriamiarimanana, F.M.; Andriamandimby, S.F.; Ravalohery, J.P.; Andriamamonjy, S.; Filippone, C.; Rakoto, D.A.D.; Telfer, S.; Heraud, J.M. Geographical Distribution and Relative Risk of Anjozorobe Virus (Thailand orthohantavirus) Infection in Black Rats (Rattus rattus) in Madagascar. Virol. J. 2018, 15, 83. [Google Scholar] [CrossRef] [Green Version]
- Olsson, G.E.; White, N.; Ahlm, C.; Elgh, F.; Verlemyr, A.C.; Juto, P.; Thomas Palo, R. Demographic Factors Associated with Hantavirus Infection in Bank Voles (Clethrionomys glareolus). Emerg. Infect. Dis. 2002, 8, 924. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N. Regulation of Rodent-Borne Viruses in the Natural Host: Implications for Human Disease. In Infectious Diseases from Nature: Mechanisms of Viral Emergence and Persistence; Springer: Vienna, Austria, 2005; pp. 45–57. [Google Scholar]
- Tian, H.; Yu, P.; Bjørnstad, O.N.; Cazelles, B.; Yang, J.; Tan, H.; Huang, S.; Cui, Y.; Dong, L.; Ma, C.; et al. Anthropogenically Driven Environmental Changes Shift the Ecological Dynamics of Hemorrhagic Fever with Renal Syndrome. PLoS Pathog. 2017, 13, e1006198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boone, J.D.; Otteson, E.W.; McGWIRE, K.C.; Villard, P.; Rowe, J.E.; St Jeor, S.C. Ecology and Demographics of Hantavirus Infections in Rodent Populations in the Walker River Basin of Nevada and California. Citeseer 1998, 59, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.; Hörnfeldt, B.; Evander, M.; Magnusson, M.; Olsson, G.; Ecke, F. Dynamics and Drivers of Hantavirus Prevalence in Rodent Populations. Vector-Borne Zoonotic Dis. 2014, 14, 537–551. [Google Scholar] [CrossRef]
- Schmaljohn, C.S. Vaccines for Hantaviruses: Progress and Issues. Expert Rev. Vaccines 2014, 11, 511–513. [Google Scholar] [CrossRef]
- Liu, R.; Ma, H.; Shu, J.; Zhang, Q.; Han, M.; Liu, Z.; Jin, X.; Zhang, F.; Wu, X. Vaccines and Therapeutics Against Hantaviruses. Front. Microbiol. 2020, 10, 2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, G.; Han, L.; An, Q.; Liu, W.; Kong, Y. Immunization Effect of Purified Bivalent Vaccine to Haemorrhagic Fever with Renal Syndrome Manufactured from Primary Cultured Hamster Kidney Cells. Chin. Med. 2005, 5, 1554–1587. [Google Scholar]
- Chen, H.X.; Luo, Z.Z.; Zhang, J.J. Hantavirus Vaccine Efficacy Evaluation Working Group. Large Scale Field Evaluation on Vaccines of Hemorrhagic Fever with Renal Syndrome in China. Chin. J. Epidemiol. 2002, 23, 145–147. [Google Scholar]
- Cho, H.W.; Howard, C.R.; Lee, H.W. Review of an Inactivated Vaccine against Hantaviruses. Intervirology 2002, 45, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Kim, C.S.; Moon, K.T. Protective Effectiveness of Hantavirus Vaccine. Emerg. Infect. Dis. 2004, 10, 2218. [Google Scholar] [CrossRef]
- Yamanishi, K.; Tanishita, O.; Tamura, M.; Asada, H.; Kondo, K.; Takagi, M.; Yoshida, I.; Konobe, T.; Fukai, K. Development of Inactivated Vaccine against Virus Causing Haemorrhagic Fever with Renal Syndrome. Vaccine 1988, 6, 278–282. [Google Scholar] [CrossRef]
- Cho, H.W.; Howard, C.R. Antibody Responses in Humans to an Inactivated Hantavirus Vaccine (Hantavax®). Vaccine 1999, 17, 2569–2575. [Google Scholar] [CrossRef]
- Song, J.Y.; Woo, H.J.; Cheong, H.J.; Noh, J.Y.; Baek, L.J.; Kim, W.J. Long-Term Immunogenicity and Safety of Inactivated Hantaan Virus Vaccine (HantavaxTM) in Healthy Adults. Vaccine 2016, 34, 1289–1295. [Google Scholar] [CrossRef]
- Sohn, Y.M.; Rho, H.O.; Park, M.S.; Kim, J.S.; Summers, P.L. Primary Humoral Immune Responses to Formalin Inactivated Hemorrhagic Fever with Renal Syndrome Vaccine (Hantavax®): Consideration of Active Immunization in South Korea. Yonsei Med. J. 2009, 42, 278–284. [Google Scholar] [CrossRef]
- Song, J.Y.; Jeong, H.W.; Yun, J.W.; Lee, J.; Woo, H.J.; Bae, J.Y.; Park, M.S.; Choi, W.S.; Park, D.W.; Noh, J.Y.; et al. Immunogenicity and Safety of a Modified Three-Dose Priming and Booster Schedule for the Hantaan Virus Vaccine (Hantavax): A Multi-Center Phase III Clinical Trial in Healthy Adults. Vaccine 2020, 38, 8016–8023. [Google Scholar] [CrossRef]
- Yi, Y.; Park, H.; Jung, J. Effectiveness of Inactivated Hantavirus Vaccine on the Disease Severity of Hemorrhagic Fever with Renal Syndrome. Kidney Res. Clin. Pr. 2018, 37, 366. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.K.; Gligic, A.; Tomanovic, S.; Bozovjc, B.; Obradovic, M.; Woo, Y.D.; An, C.N.; Kim, H.; Jiang, Y.S.; Park, S.C.; et al. A Field Efficacy Trial of Inactivated Hantaan Virus Vaccine (Hantavax(TM)) Against Hemorrhagic Fever with Renal Syndrome (HFRS) in the Endemic Areas of Yugoslavia from 1996 to 1998. J. Korean Soc. Virol. 1999, 29, 55–64. [Google Scholar]
- Jung, J.; Ko, S.-J.; Oh, H.S.; Moon, S.M.; Song, J.-W.; Huh, K. Protective Effectiveness of Inactivated Hantavirus Vaccine Against Hemorrhagic Fever with Renal Syndrome. J. Infect. Dis. 2018, 217, 1417–1420. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wei, Z.; Wei, J.; Ma, C.; Dong, J.; Lu, X.; Zheng, Y.; Yu, P.; Qu, J.; Dong, L. Long Term Epidemiological Effects of Vaccination on Hemorrhagical Fever with Renal Syndrome (HFRS) in Shaanxi Provincial HFRS Epidemic Areas. Zhonghua Liu Xing Bing Xue Za Zhi 2012, 33, 309–312. [Google Scholar] [PubMed]
- Zheng, Y.; Zhou, B.-Y.; Wei, J.; Xu, Y.; Dong, J.-H.; Guan, L.-Y.; Ma, P.; Yu, P.-B.; Wang, J.-J. Persistence of Immune Responses to Vaccine against Haemorrhagic Fever with Renal Syndrome in Healthy Adults Aged 16–60 Years: Results from an Open-Label2-Year Follow-up Study. Infect. Dis. 2018, 50, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Clement, J.; van Ranst, M. Recent Approaches in Hantavirus Vaccine Development. Expert Rev. Vaccines 2009, 8, 67–76. [Google Scholar] [CrossRef]
- Li, Z.; Zeng, H.; Wang, Y.; Zhang, Y.; Cheng, L.; Zhang, F.; Lei, Y.; Jin, B.; Ma, Y.; Chen, L. The Assessment of Hantaan Virus-Specific Antibody Responses after the Immunization Program for Hemorrhagic Fever with Renal Syndrome in Northwest China. Hum. Vaccines Immunother. 2017, 13, 802–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dzagurova, T.K.; Siniugina, A.A.; Ishmukhametov, A.A.; Egorova, M.S.; Kurashova, S.S.; Balovneva, M.V.; Deviatkin, A.A.; Tkachenko, P.E.; Leonovich, O.A.; Tkachenko, E.A. Pre-Clinical Studies of Inactivated Polyvalent HFRS Vaccine. Front. Cell. Infect. Microbiol. 2020, 10, 545372. [Google Scholar] [CrossRef]
- Wagenknecht, H.-A. Book Review. Protein Sci. 2009, 13, 3331–3332. [Google Scholar] [CrossRef]
- Acuna, R.; Cifuentes-Munoz, N.; Marquez, C.L.; Bulling, M.; Klingstrom, J.; Mancini, R.; Lozach, P.-Y.; Tischler, N.D. Hantavirus Gn and Gc Glycoproteins Self-Assemble into Virus-Like Particles. J. Virol. 2014, 88, 2344–2348. [Google Scholar] [CrossRef] [Green Version]
- Fougeroux, C.; Goksøyr, L.; Idorn, M.; Soroka, V.; Myeni, S.K.; Dagil, R.; Janitzek, C.M.; Søgaard, M.; Aves, K.-L.; Horsted, E.W.; et al. Capsid-like Particles Decorated with the SARS-CoV-2 Receptor-Binding Domain Elicit Strong Virus Neutralization Activity. Nat. Commun. 2021, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.F.; Wang, F.; Zhang, L.; Yu, L.; Ye, W.; Liu, Z.Y.; Ying, Q.K.; Wu, X.A.; Xu, Z.K.; Zhang, F.L. Incorporation of GM-CSF or CD40L Enhances the Immunogenicity of Hantaan Virus-like Particles. Front. Cell. Infect. Microbiol. 2016, 6, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Ma, T.; Zhang, X.; Ying, Q.; Han, M.; Zhang, M.; Yang, R.; Li, Y.; Wang, F.; Liu, R.; et al. Incorporation of CD40 Ligand or Granulocyte-Macrophage Colony Stimulating Factor into Hantaan Virus (HTNV) Virus-like Particles Significantly Enhances the Long-Term Immunity Potency against HTNV Infection. J. Med. Microbiol. 2019, 68, 480–492. [Google Scholar] [CrossRef]
- Koletzki, D.; Biel, S.S.; Meisel, H.; Nugel, E.; Gelderblom, H.R.; Krüger, D.H.; Ulrich, R. HBV Core Particles Allow the Insertion and Surface Exposure of the Entire Potentially Protective Region of Puumala Hantavirus Nucleocapsid Protein. Biol. Chem. 1999, 380, 325–333. [Google Scholar] [CrossRef]
- Ulrich, R.; Koletzki, D.; Lachmann, S.; Lundkvist, Å.; Zankl, A.; Kazaks, A.; Kurth, A.; Gelderblom, H.R.; Borisova, G.; Meisel, H.; et al. New Chimaeric Hepatitis B Virus Core Particles Carrying Hantavirus (Serotype puumala) Epitopes: Immunogenicity and Protection against Virus Challenge. J. Biotechnol. 1999, 73, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Mcclain, D.J.; Summers, P.L.; Harrison, S.A.; Schmaljohn, A.L.; Schmaljohn, C.S. Clinical Evaluation of a Vaccinia-Vectored Hantaan Virus Vaccine. J. Med. Virol. 2000, 60, 77–85. [Google Scholar] [CrossRef]
- Geldmacher, A.; Skrastina, D.; Petrovskis, I.; Borisova, G.; Berriman, J.A.; Roseman, A.M.; Crowther, R.A.; Fischer, J.; Musema, S.; Gelderblom, H.R.; et al. An Amino-Terminal Segment of Hantavirus Nucleocapsid Protein Presented on Hepatitis B Virus Core Particles Induces a Strong and Highly Cross-Reactive Antibody Response in Mice. Virology 2004, 323, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C.S.; Hasty, S.E.; Dalrymple, J.M. Preparation of Candidate Vaccinia-Vectored Vaccines for Haemorrhagic Fever with Renal Syndrome. Vaccine 1992, 10, 10–13. [Google Scholar] [CrossRef]
- Chu, Y.-K.; Jennings, G.B.; Schmaljohn, C.S. A Vaccinia Virus-Vectored Hantaan Virus Vaccine Protects Hamsters from Challenge with Hantaan and Seoul Viruses but Not Puumala Virus. J. Virol. 1995, 69, 6417–6423. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, E.F.; Josleyn, M.; Ullman, D.; Fisher, D.; Dalrymple, L.; Sellers-Myers, K.; Loudon, P.; Rusnak, J.; Rivard, R.; Schmaljohn, C.; et al. A Phase 1 Clinical Trial of Hantaan Virus and Puumala Virus M-Segment DNA Vaccines for Hemorrhagic Fever with Renal Syndrome. Vaccine 2012, 30, 1951–1958. [Google Scholar] [CrossRef]
- Hooper, J.W.; Custer, D.M.; Thompson, E.; Schmaljohn, C.S. DNA Vaccination with the Hantaan Virus M Gene Protects Hamsters against Three of Four HFRS Hantaviruses and Elicits a High-Titer Neutralizing Antibody Response in Rhesus Monkeys. J. Virol. 2001, 75, 8469–8477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, J.W.; Custer, D.M.; Smith, J.; Wahl-Jensen, V. Hantaan/Andes Virus DNA Vaccine Elicits a Broadly Cross-Reactive Neutralizing Antibody Response in Nonhuman Primates. Virology 2006, 347, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klasse, P.J.; Sattentau, Q.J. Occupancy and Mechanism in Antibody-Mediated Neutralization of Animal Viruses. J. Gen. Virol. 2002, 83, 2091–2108. [Google Scholar] [CrossRef] [PubMed]
- Pantophlet, R.; Burton, D.R. GP120: Target for Neutralizing HIV-1 Antibodies. Immunol. Annu. Rev. 2006, 24, 739–769. [Google Scholar] [CrossRef] [PubMed]
- Dantas, J.R.; Okuno, Y.; Asada, H.; Tamura, M.; Takahashi, M.; Tanishita, O.; Takahashi, Y.; Kurata, T.; Yamanishi, K. Characterization of Glycoproteins of Viruses Causing Hemorrhagic Fever with Renal Syndrome (HFRS) Using Monoclonal Antibodies. Virology 1986, 151, 379–384. [Google Scholar] [CrossRef]
- Pettersson, L.; Thunberg, T.; Rocklöv, J.; Klingström, J.; Evander, M.; Ahlm, C. Viral Load and Humoral Immune Response in Association with Disease Severity in Puumala Hantavirus-Infected Patients-Implications for Treatment. Clin. Microbiol. Infect. 2014, 20, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Park, S.M.; Kim, J. A Soluble and Heat-Resistant Form of Hantavirus Nucleocapsid Protein for the Serodiagnosis of HFRS. J. Virol. Methods 2008, 147, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rissanen, I.; Krumm, S.A.; Stass, R.; Whitaker, A.; Voss, J.E.; Bruce, E.A.; Rothenberger, S.; Kunz, S.; Burton, D.R.; Huiskonen, J.T.; et al. Structural Basis for a Neutralizing Antibody Response Elicited by a Recombinant Hantaan Virus Gn Immunogen. mBio 2021, 12, e02531-20. [Google Scholar] [CrossRef]
- Lundkvist, Å.; Niklasson, B. Bank Vole Monoclonal Antibodies against Puumala Virus Envelope Glycoproteins: Identification of Epitopes Involved in Neutralization. Arch. Virol. 1992, 126, 93–105. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, Y.; Wang, J.; Lv, T.; Jin, B. Identification of Three Novel CTL Epitopes within Nucleocapsid Protein of Hantaan Virus. Viral Immunol. 2011, 24, 449–454. [Google Scholar] [CrossRef] [Green Version]
- Elgh, F.; Linderholm, M.; Wadell, G.; Tärnvik, A.; Juto, P. Development of Humoral Cross-Reactivity to the Nucleocapsid Protein of Heterologous Hantaviruses in Nephropathia Epidemica. FEMS Immunol. Med. Microbiol. 1998, 22, 309–315. [Google Scholar] [CrossRef]
- De Carvalho Nicacio, C.; Gonzalez Della Valle, M.; Padula, P.; Björling, E.; Plyusnin, A.; Lundkvist, Å. Cross-Protection against Challenge with Puumala Virus after Immunization with Nucleocapsid Proteins from Different Hantaviruses. J. Virol. 2002, 76, 6669–6677. [Google Scholar] [CrossRef] [Green Version]
- Geldmacher, A.; Schmaler, M.; Krüger, D.H.; Ulrich, R. Yeast-Expressed Hantavirus Dobrava Nucleocapsid Protein Induces a Strong, Long-Lasting, and Highly Cross-Reactive Immune Response in Mice. Viral. Immunol. 2004, 17, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Jandrig, B.; Klempa, B.; Yoshimatsu, K.; Arikawa, J.; Meisel, H.; Niedrig, M.; Pitra, C.; Krüger, D.H.; Ulrich, R. Nucleocapsid Protein of Cell Culture-Adapted Seoul Virus Strain 80-39: Analysis of Its Encoding Sequence, Expression in Yeast and Immuno-Reactivity. Virus Genes 2005, 30, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Shkair, L.; Garanina, E.; Martynova, E.; Kolesnikova, A.I. Immunogenic Properties of MVs Containing Structural Hantaviral Proteins: An Original Study. Pharmaceutics 2022, 1, 93. [Google Scholar] [CrossRef]
- Krüger, D.; Schönrich, G.; Klempa, B. Human Pathogenic Hantaviruses and Prevention of Infection. Hum. Vaccines 2011, 7, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vial, P.A.; Valdivieso, F.; Calvo, M.; Rioseco, M.L.; Riquelme, R.; Araneda, A.; Tomicic, V.; Graf, J.; Paredes, L.; Florenzano, M.; et al. A Non-Randomized Multicentre Trial of Human Immune Plasma for Treatment of Hantavirus Cardiopulmonary Syndrome Caused by Andes Virus. Antivir. Ther. 2015, 20, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.K.; Takashima, I.; Hashimoto, N. Characteristics of Passive Immunity against Hantavirus Infection in Rats. Arch. Virol. 1989, 105, 235–246. [Google Scholar] [CrossRef]
- Xu, Z.; Wei, L.; Wang, L.; Wang, H.; Jiang, S. The in Vitro and in Vivo Protective Activity of Monoclonal Antibodies Directed against Hantaan Virus: Potential Application for Immunotherapy and Passive. Biochem. Biophys. Res. Commun. 2002, 298, 552–558. [Google Scholar] [CrossRef]
- Rong, X.; Xiao, Y.Y.; Dao, F.Y.; Chang, Y.Z.; Pei, L.G.; Fan, D.Z. Phase I Evaluation of the Safety and Pharmacokinetics of a Single-Dose Intravenous Injection of a Murine Monoclonal Antibody against Hantaan Virus in Healthy Volunteers. Antimicrob. Agents Chemother. 2009, 53, 5055–5059. [Google Scholar] [CrossRef] [Green Version]
- Hall, P.; Leitão, A.; Ye, C.; Kilpatrick, K.; Hjelle, B. Small Molecule Inhibitors of Hantavirus Infection. Bioorg. Med. Chem. Lett. 2010, 20, 7085–7091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.-W.; Song, K.-J.; Baek, L.-J.; Frost, B.; Poncz, M.; Park, K. In Vivo Characterization of the Integrin Β3 as a Receptor for Hantaan Virus Cellular Entry. Exp. Mol. Med. 2005, 37, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.R.; Hjelle, B.; Brown, D.C.; Ye, C.; Bondu-Hawkins, V.; Kilpatrick, K.A.; Larson, R.S. Multivalent Presentation of Antihantavirus Peptides on Nanoparticles Enhances Infection Blockade. Antimicrob. Agents Chemother. 2008, 52, 2079–2088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huggins, J.W.; Hsiang, C.M.; Cosgriff, T.M.; Guang, M.Y.; Smith, J.I.; Wu, Z.O.; LeDuc, J.W.; Zheng, Z.M.; Meegan, J.M.; Wang, Q.N.; et al. Prospective, Double-Blind, Concurrent, Placebo-Controlled Clinical Trial of Intravenous Ribavirin Therapy of Hemorrhagic Fever with Renal Syndrome. J. Infect. Dis. 1991, 164, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Malinin, O.V.; Platonov, A.E. Insufficient Efficacy and Safety of Intravenous Ribavirin in Treatment of Haemorrhagic Fever with Renal Syndrome Caused by Puumala Virus. Infect. Dis. 2017, 49, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Severson, W.E.; Schmaljohn, C.S.; Javadian, A.; Jonsson, C.B. Ribavirin Causes Error Catastrophe during Hantaan Virus Replication. J. Virol. 2003, 77, 481–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, Y.; Komeno, T.; Nakamura, T. Favipiravir (T-705), a Broad Spectrum Inhibitor of Viral RNA Polymerase. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 449–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayor, J.; Engler, O.; Rothenberger, S. Antiviral Efficacy of Ribavirin and Favipiravir against Hantaan Virus. Microorganisms 2021, 9, 1306. [Google Scholar] [CrossRef]
- Taylor, S.L.; Wahl-Jensen, V.; Copeland, A.M.; Jahrling, P.B.; Schmaljohn, C.S. Endothelial Cell Permeability during Hantavirus Infection Involves Factor XII-Dependent Increased Activation of the Kallikrein-Kinin System. PLoS Pathog. 2013, 9, e1003470. [Google Scholar] [CrossRef]
- Antonen, J.; Leppänen, I. A Severe Case of Puumala Hantavirus Infection Successfully Treated with Bradykinin Receptor Antagonist Icatibant. J. Infect. 2013, 45, 494–496. [Google Scholar] [CrossRef]
- Laine, O.; Leppänen, I.; Koskela, S.; Antonen, J.; Mäkelä, S.; Sinisalo, M.; Vaheri, A.; Mustonen, J. Severe Puumala Virus Infection in a Patient with a Lymphoproliferative Disease Treated with Icatibant. Infect. Dis. 2015, 47, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Rhen, T.; Cidlowski, J.A. Antiinflammatory Action of Glucocorticoids—New Mechanisms for Old Drugs. N. Engl. J. Med. 2005, 353, 1711–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Easterbrook, J.D.; Klein, S.L. Corticosteroids Modulate Seoul Virus Infection, Regulatory T Cell Responses, and MMP-9 Expression in Male, but Not Female, Norway Rats. J. Gen. Virol. 2008, 89, 2723. [Google Scholar] [CrossRef] [PubMed]
- Steer, A. Pathology of Hemorrhagic Fever: A Comparison of the Findings: 1951 and 1952. Am. J. Pathol. 1955, 3, 201–221. [Google Scholar]
- Hautala, T.; Sironen, T.; Vapalahti, O.; Pääkkö, E.; Särkioja, T.; Salmela, P.I.; Vaheri, A.; Plyusnin, A.; Kauma, H. Hypophyseal Hemorrhage and Panhypopituitarism during Puumala Virus Infection: Magnetic Resonance Imaging and Detection of Viral Antigen in the Hypophysis. Clin. Infect. Dis. 2002, 35, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Seitsonen, E.; Hynninen, M.; Kolho, E.; Kallio-Kokko, H.; Pettilä, V. Corticosteroids Combined with Continuous Veno-Venous Hemodiafiltration for Treatment of Hantavirus Pulmonary Syndrome Caused by Puumala Virus Infection. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 261–266. [Google Scholar] [CrossRef]
- Dunst, R.; Mettang, T.; Kuhlmann, U. Severe Thrombocytopenia and Response to Corticosteroids in a Case of Nephropathia Epidemica. Am. J. Kidney Dis. 1998, 31, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Latus, J.; Kitterer, D.; Segerer, S.; Artunc, F.; Alscher, M.D.; Braun, N. Severe Thrombocytopenia in Hantavirus-Induced Nephropathia Epidemica. Infection 2015, 43, 83–87. [Google Scholar] [CrossRef]
- Vial, P.; Valdivieso, F.; Ferres, M.; Riquelme, R.; Rioseco, M.L.; Calvo, M.; Castillo, C.; Díaz, R.; Scholz, L.; Cuiza, A.; et al. Hantavirus Study Group in Chile. High-Dose Intravenous Methylprednisolone for Hantavirus Cardiopulmonary Syndrome in Chile: A Double-Blind, Randomized Controlled Clinical Trial. Clin. Infect. Dis. 2013, 57, 943–951. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention Hemorrhagic Fever with Renal Syndrome (HFRS)—Hantavirus. Available online: https://www.cdc.gov/hantavirus/hfrs/index.html#:~:text=Suggested%20Reading-,What%20is%20hemorrhagic%20fever%20with%20renal%20syndrome%3F,hemorrhagic%20fever%2C%20and%20nephropathia%20epidemica (accessed on 15 September 2022).
- Connolly-Andersen, A.-M.; Rasmuson, J.; Öman, M.; Ahlm, C. Mesenteric Vein Thrombosis Following Platelet Transfusion in a Patient with Hemorrhagic Fever with Renal Syndrome: A Case Report. TH Open 2018, 2, e261–e264. [Google Scholar] [CrossRef] [Green Version]
- Ying, X.; Lai, X.; Jin, X.; Cai, L.; Li, X. Continuous Renal Replacement Therapy Rescues Severe Haemorrhagic Fever with Renal Syndrome in Pregnancy: A Case Report. BMC Infect. Dis 2020, 20, 920. [Google Scholar] [CrossRef]
- Kim, H.J.; Han, S.W. Diagnostic Challenge of Hemorrhagic Fever with Renal Syndrome on Admission before Its Serological Confirmation. Korean J. Nephrol. 2004, 23, 82–91. [Google Scholar]
- Wichmann, D.; Slenczka, W.; Alter, P.; Boehm, S.; Feldmann, H. Hemorrhagic Fever with Renal Syndrome: Diagnostic Problems with a Known Disease. J. Clin. Microbiol. 2001, 39, 3414–3416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groen, J.; Gerding, M.; Jordans, J.G.M.; Clement, J.P.; Osterhaus, A.D.M.E. Class and Subclass Distribution of Hantavirus-Specific Serum Antibodies at Different Times after the Onset of Nephropathia Epidemica. J. Med. Virol. 1994, 43, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Sakhautdinov, V.G.; Galimov, O.V.; Nurtdinov, M.A.; Sibaev, I.M. Differential Diagnosis of Hemorrhagic Fever with Renal Syndrome and Acute Surgical Diseases of Abdominal Organs. Khirurgiia 2001, 3, 23–25. (In Russian) [Google Scholar]
- Dusek, J.; Pejcoch, M.; Kolsky, A.; Seeman, T.; Nemec, V.; Stejskal, J.; Vondrak, K.; Janda, J. Mild Course of Puumala Nephropathy in Children in an Area with Sporadic Occurrence Hantavirus Infection. Pediatr. Nephrol. 2006, 21, 1889–1892. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, Q.S.; Zhang, Y.; Sun, B.C.; Zhao, L.Y. Analysis of Misdiagnosed Cases of Hemorrhagic Fever with Renal Syndrome in Children: Two Cases and Literature Review. BMC Nephrol. 2019, 20, 383. [Google Scholar] [CrossRef] [PubMed]
- Mattar, S.; Guzmán, C.; Figueiredo, L.T. Diagnosis of Hantavirus Infection in Humans. Expert. Rev. Anti. Infect. 2015, 13, 939–946. [Google Scholar] [CrossRef]
- Galimzyanov, K.M.; Vasil’kova, V.V.; Cherenov, I.V.; Seidov, K.S.; Akmaeva, L.R.; Zhidovinov, A.A. Differential diagnostic criteria of kidney injury due to leptospirosis. Arch. Euromed. 2017, 7, 41–43. [Google Scholar]
- Yang, C.W. Leptospirosis Renal Disease: Understanding the Initiation by Toll-like Receptors. Kidney Int. 2007, 72, 918–925. [Google Scholar] [CrossRef] [Green Version]
- Ciferska, H.; Honsova, E.; Lodererova, A.; Hruskova, Z.; Neprasova, M.; Vachek, J.; Suchanek, M.; Zima, T.; Coppo, R.; Tesar, V.; et al. Does the Renal Expression of Toll-like Receptors Play a Role in Patients with IgA Nephropathy? J. Nephrol. 2020, 33, 307–316. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orthohantavirus Species | Natural Host Reservoir (Common Name) | Geographic Distribution | Associated Disease | References |
|---|---|---|---|---|
| Hantaan orthohantavirus (HTNV) * | Apodemus agrarius (striped field mouse) | China, Korea, Russia | HFRS | [34] |
| Dabieshan orthohantavirus (DBSV) | Niviventer confucianus (Chinese white-bellied rat) | China | Unknown | [35] |
| Fugong orthohantavirus (FUGV) | Eothenomys eleusis (small oriental vole) | China | Unknown | [36] |
| Fusong orthohantavirus (FUSV) | Microtus fortis (reed vole) | China | Unknown | [37] |
| Imjin orthohantavirus (MJNV) | Crocidura lasiura (Ussuri white-toothed shrew) | China | Unknown | [38,39] |
| Soochong orthohantavirus (SOOV) * | Apodemus peninsulae (Korean field mouse) | Korea | Unknown | [40] |
| Amur orthohantavirus (AMRV) * | Apodemus peninsulae (Korean field mouse) | Asia | HFRS | [40] |
| Seoul orthohantavirus (SEOV) | Rattus norvegicus Rattus rattus (rats) | Worldwide | HFRS | [34] |
| Serang orthohantavirus (SERV) | Rattus tanezumi (Asian house rat) | Indonesia | Mild HFRS | [41] |
| Thailand orthohantavirus (THAIV) | Bandicoot indica (Bandicoot rat) | Southeast Asia (Thailand) | HFRS | [42] |
| Puumala orthohantavirus (PUUV) ** | Myodes glareolus (Bank vole) | Eurasian continent | Mild HFRS/NE | [43] |
| Khabarovsk orthohantavirus (KHAV) | Microtus maximowiczii (Maximowicz’s vole) | Mongolia, northeast China | unknown | [37] |
| Hokkaido virus (HOKV) ** | Myodes rufocanus (Grey red-backed vole) | Japan | HFRS | [44] |
| Muju virus (MUJI) ** | Myodes regulus (Royal vole) | South Korea | unknown | [45] |
| Luxi orthohantavirus (LUXV) | Eothenomys miletus (Yunnan red-backed vole) | Asia (China) | unknown | [46] |
| Jeju orthohantavirus (JJUV) | Crocidura shantungensis (Asian lesser white-toothed shrew) | Asia (South Korea) | unknown | [39] |
| Cao Bang orthohantavirus (CBNV) | Anourosorex squamipes (Chinese mole shrew) | China and Vietnam | unknown | [47] |
| Yakeshi orthohantavirus (YKSV) | Sorex isodon (Taiga shrew) | Asia (China) | unknown | [48] |
| Asama orthohantavirus (ASAV) | Ulotrichous talpoides (Japanese shrew mole) | Asia (Japan) | unknown | [49] |
| Kenkeme orthohantavirus (KKMV) | Sorex roboratus (flat-skulled shrew) | Altai (Russia), northeast China | unknown | [50] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehgal, A.; Mehta, S.; Sahay, K.; Martynova, E.; Rizvanov, A.; Baranwal, M.; Chandy, S.; Khaiboullina, S.; Kabwe, E.; Davidyuk, Y. Hemorrhagic Fever with Renal Syndrome in Asia: History, Pathogenesis, Diagnosis, Treatment, and Prevention. Viruses 2023, 15, 561. https://doi.org/10.3390/v15020561
Sehgal A, Mehta S, Sahay K, Martynova E, Rizvanov A, Baranwal M, Chandy S, Khaiboullina S, Kabwe E, Davidyuk Y. Hemorrhagic Fever with Renal Syndrome in Asia: History, Pathogenesis, Diagnosis, Treatment, and Prevention. Viruses. 2023; 15(2):561. https://doi.org/10.3390/v15020561
Chicago/Turabian StyleSehgal, Ayushi, Sanya Mehta, Kritika Sahay, Ekaterina Martynova, Albert Rizvanov, Manoj Baranwal, Sara Chandy, Svetlana Khaiboullina, Emmanuel Kabwe, and Yuriy Davidyuk. 2023. "Hemorrhagic Fever with Renal Syndrome in Asia: History, Pathogenesis, Diagnosis, Treatment, and Prevention" Viruses 15, no. 2: 561. https://doi.org/10.3390/v15020561
APA StyleSehgal, A., Mehta, S., Sahay, K., Martynova, E., Rizvanov, A., Baranwal, M., Chandy, S., Khaiboullina, S., Kabwe, E., & Davidyuk, Y. (2023). Hemorrhagic Fever with Renal Syndrome in Asia: History, Pathogenesis, Diagnosis, Treatment, and Prevention. Viruses, 15(2), 561. https://doi.org/10.3390/v15020561