
Quantitative Proteomics for the Identification of Differentially Expressed Proteins in the Extracellular Vesicles of Cervical Cancer Cells

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Extracellular Vesicle (EV) Isolation
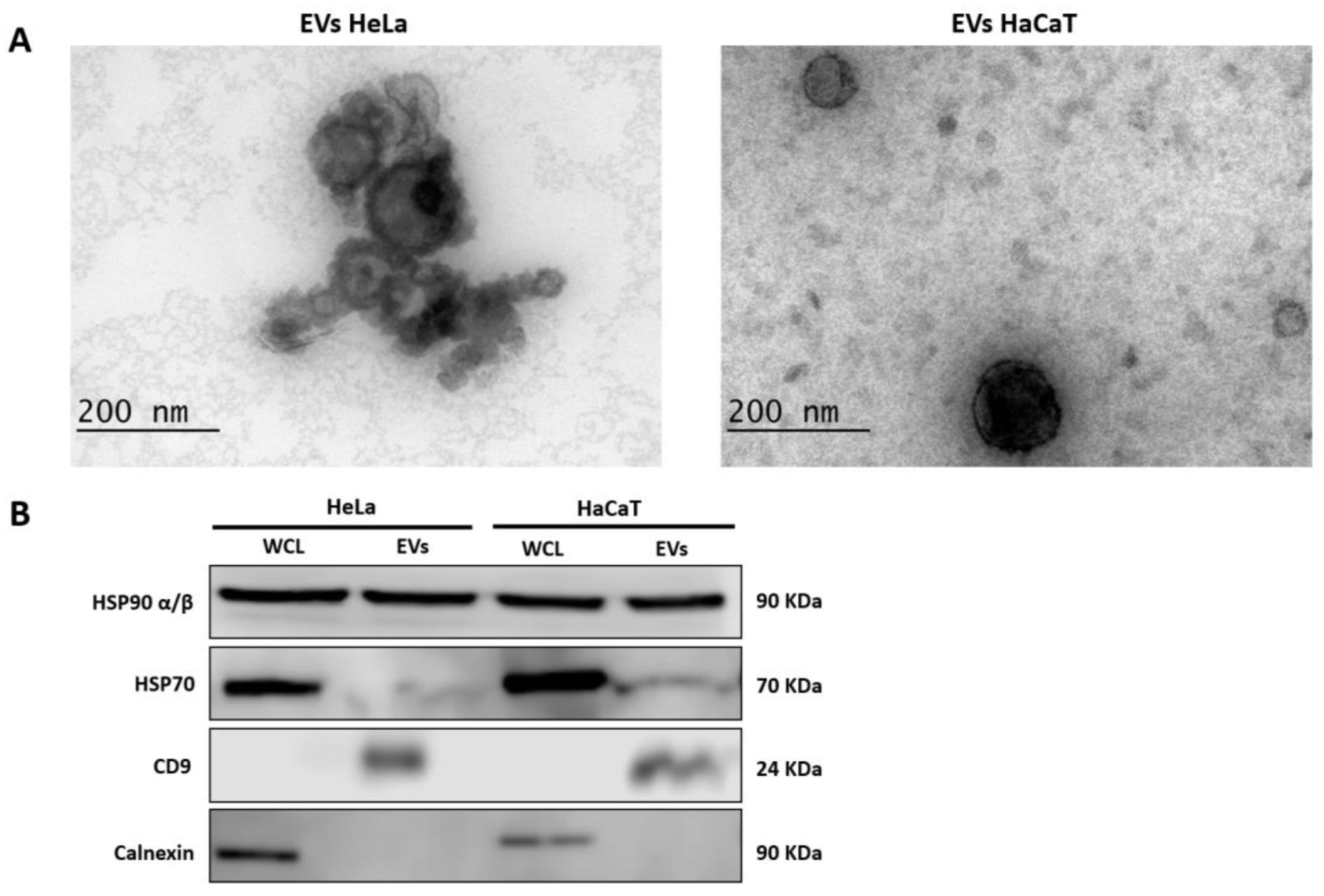
2.3. Extracellular Vesicle Identification
2.3.1. Transmission Electron Microscopy (TEM)
2.3.2. Western Blot
2.4. Protein Extraction and Quantification
2.5. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS–PAGE)
2.6. Label-Free Based Quantitative Proteomic Analysis
2.6.1. In-Gel Digestion
2.6.2. Nano LC-MS/MS Analysis
2.6.3. Analysis of LC-MS/MS Data
2.7. Bioinformatic Analysis
3. Results
3.1. Identification of Extracellular Vesicles
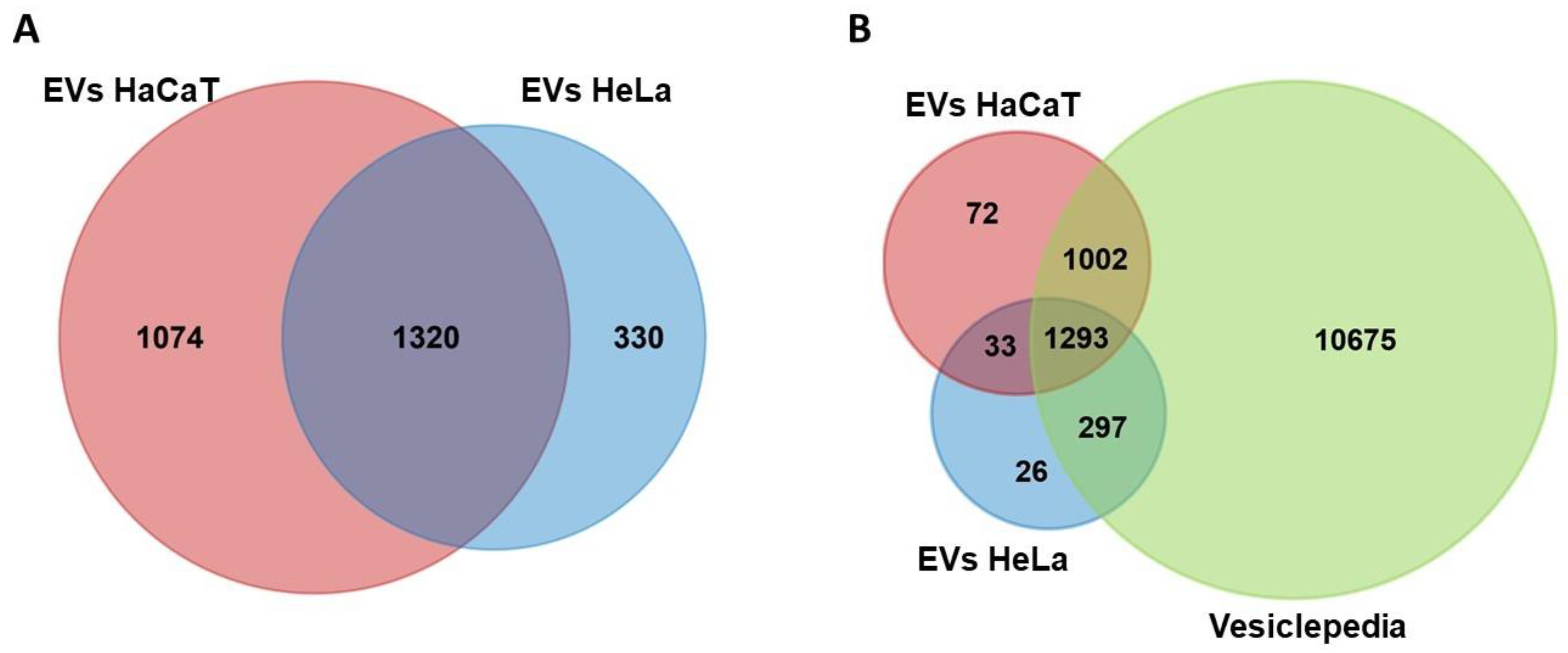
3.2. Analysis of Protein Content in HeLa and HaCaT EVs and Comparison with the Vesiclepedia Database
3.3. Differential Expression of Proteins Identified in the EVs
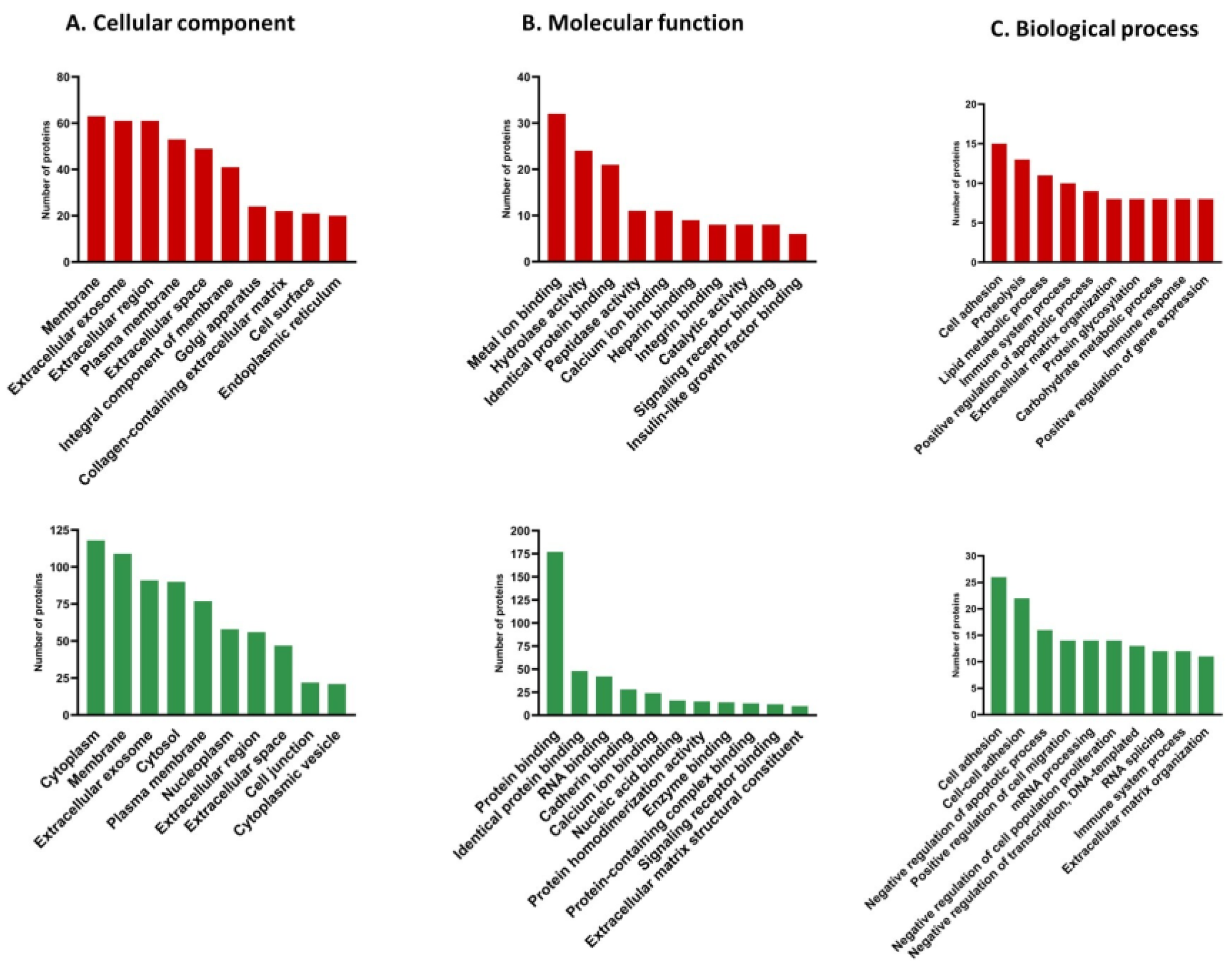
3.4. Gene Ontology (GO) Analysis of Differentially Expressed Proteins
3.5. Biological Pathway Analyses of Differentially Expressed Proteins
3.6. Interaction Network of Dysregulated Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Walboomers, J.M.; Jacobs, M.V.; Manos, M.M.; Bosch, F.X.; Kummer, J.A.; Shah, K.V.; Snijders, P.J.; Peto, J.; Meijer, C.J.; Munoz, N. Human papillomavirus is a necessary cause of invasive cervical cancer worldwide. J. Pathol. 1999, 189, 12–19. [Google Scholar] [CrossRef]
- Qiu, J.J.; Sun, S.G.; Tang, X.Y.; Lin, Y.Y.; Hua, K.Q. Extracellular vesicular Wnt7b mediates HPV E6-induced cervical cancer angiogenesis by activating the beta-catenin signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 260. [Google Scholar] [CrossRef] [PubMed]
- Castellsague, X.; Munoz, N. Chapter 3: Cofactors in human papillomavirus carcinogenesis—Role of parity, oral contraceptives, and tobacco smoking. J. Natl. Cancer Inst. Monogr. 2003, 2003, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhang, X.; Jiang, L.; Li, Y.; Zheng, Q. Tumor endothelial cell-derived extracellular vesicles contribute to tumor microenvironment remodeling. Cell Commun. Signal. 2022, 20, 97. [Google Scholar] [CrossRef]
- Lin, Y.; Zhang, C.; Xiang, P.; Shen, J.; Sun, W.; Yu, H. Exosomes derived from HeLa cells break down vascular integrity by triggering endoplasmic reticulum stress in endothelial cells. J. Extracell Vesicles 2020, 9, 1722385. [Google Scholar] [CrossRef] [Green Version]
- Luo, W. Nasopharyngeal Carcinoma Ecology Theory: Cancer as Multidimensional Spatiotemporal “Unity of Ecology and Evolution” Pathological Ecosystem. Preprints 2022, 2022100226. [Google Scholar] [CrossRef]
- Gyorgy, B.; Szabo, T.G.; Pasztoi, M.; Pal, Z.; Misjak, P.; Aradi, B.; Laszlo, V.; Pallinger, E.; Pap, E.; Kittel, A.; et al. Membrane vesicles, current state-of-the-art: Emerging role of extracellular vesicles. Cell Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [Green Version]
- Thery, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Ciardiello, C.; Cavallini, L.; Spinelli, C.; Yang, J.; Reis-Sobreiro, M.; de Candia, P.; Minciacchi, V.R.; Di Vizio, D. Focus on Extracellular Vesicles: New Frontiers of Cell-to-Cell Communication in Cancer. Int. J. Mol. Sci. 2016, 17, 175. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.Y.; Chen, C. Toward characterizing extracellular vesicles at a single-particle level. J. Biomed. Sci. 2019, 26, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, J.; Tkach, M. Dendritic cell extracellular vesicles. Int. Rev. Cell Mol. Biol. 2019, 349, 213–249. [Google Scholar] [CrossRef] [PubMed]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2019, 47, D516–D519. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Sanchez, V.; Rodriguez-Hernandez, R.M.; Aguilar-Ruiz, S.R.; Torres-Aguilar, H.; Romero-Tlalolini, M.L.A. Extracellular Vesicles in Cervical Cancer and HPV Infection. Membranes 2021, 11, 453. [Google Scholar] [CrossRef] [PubMed]
- Son, S.H.; Gangadaran, P.; Ahn, B.C. A novel strategy of transferring NIS protein to cells using extracellular vesicles leads to increase in iodine uptake and cytotoxicity. Int. J. Nanomed. 2019, 14, 1779–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandu, R.; Oh, J.W.; Kim, K.P. Mass spectrometry-based proteome profiling of extracellular vesicles and their roles in cancer biology. Exp. Mol. Med. 2019, 51, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honegger, A.; Leitz, J.; Bulkescher, J.; Hoppe-Seyler, K.; Hoppe-Seyler, F. Silencing of human papillomavirus (HPV) E6/E7 oncogene expression affects both the contents and the amounts of extracellular microvesicles released from HPV-positive cancer cells. Int. J. Cancer 2013, 133, 1631–1642. [Google Scholar] [CrossRef]
- Ranjit, S.; Kodidela, S.; Sinha, N.; Chauhan, S.; Kumar, S. Extracellular Vesicles from Human Papilloma Virus-Infected Cervical Cancer Cells Enhance HIV-1 Replication in Differentiated U1 Cell Line. Viruses 2020, 12, 239. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.; Sharma, A.; Bharti, A.C. Upstream Hedgehog signaling components are exported in exosomes of cervical cancer cell lines. Nanomedicine 2018, 13, 2127–2138. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Qasim, M.; Khan, K.; Kim, J.H. Biogenesis, Membrane Trafficking, Functions, and Next Generation Nanotherapeutics Medicine of Extracellular Vesicles. Int. J. Nanomed. 2021, 16, 3357–3383. [Google Scholar] [CrossRef]
- Guenat, D.; Hermetet, F.; Pretet, J.L.; Mougin, C. Exosomes and Other Extracellular Vesicles in HPV Transmission and Carcinogenesis. Viruses 2017, 9, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meehan, K.; Vella, L.J. The contribution of tumour-derived exosomes to the hallmarks of cancer. Crit. Rev. Clin. Lab. Sci. 2016, 53, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.; Yadav, J.; Thakur, K.; Aggarwal, N.; Chhokar, A.; Tripathi, T.; Singh, T.; Jadli, M.; Veerapandian, V.; Bharti, A.C. Transcriptome analysis of cervical cancer exosomes and detection of HPVE6*I transcripts in exosomal RNA. BMC Cancer 2022, 22, 164. [Google Scholar] [CrossRef] [PubMed]
- Cho, O.; Kim, D.W.; Cheong, J.Y. Screening Plasma Exosomal RNAs as Diagnostic Markers for Cervical Cancer: An Analysis of Patients Who Underwent Primary Chemoradiotherapy. Biomolecules 2021, 11, 1691. [Google Scholar] [CrossRef]
- Tong, F.; Andress, A.; Tang, G.; Liu, P.; Wang, X. Comprehensive profiling of extracellular RNA in HPV-induced cancers using an improved pipeline for small RNA-seq analysis. Sci. Rep. 2020, 10, 19450. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, X.; Meng, L.; Li, W.; Li, C.; Li, P.; Xu, S. Changes of miRNA Expression Profiles from Cervical-Vaginal Fluid-Derived Exosomes in Response to HPV16 Infection. Biomed. Res. Int. 2020, 2020, 7046894. [Google Scholar] [CrossRef]
- Choi, D.S.; Kim, D.K.; Kim, Y.K.; Gho, Y.S. Proteomics, transcriptomics and lipidomics of exosomes and ectosomes. Proteomics 2013, 13, 1554–1571. [Google Scholar] [CrossRef]
- Khan, S.; Jutzy, J.M.; Valenzuela, M.M.; Turay, D.; Aspe, J.R.; Ashok, A.; Mirshahidi, S.; Mercola, D.; Lilly, M.B.; Wall, N.R. Plasma-derived exosomal survivin, a plausible biomarker for early detection of prostate cancer. PLoS ONE 2012, 7, e46737. [Google Scholar] [CrossRef] [Green Version]
- Logozzi, M.; De Milito, A.; Lugini, L.; Borghi, M.; Calabro, L.; Spada, M.; Perdicchio, M.; Marino, M.L.; Federici, C.; Iessi, E.; et al. High levels of exosomes expressing CD63 and caveolin-1 in plasma of melanoma patients. PLoS ONE 2009, 4, e5219. [Google Scholar] [CrossRef] [Green Version]
- Taylor, D.D.; Gercel-Taylor, C. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol. Oncol. 2008, 110, 13–21. [Google Scholar] [CrossRef]
- Deyrieux, A.F.; Wilson, V.G. In vitro culture conditions to study keratinocyte differentiation using the HaCaT cell line. Cytotechnology 2007, 54, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glady, A.; Vandebroek, A.; Yasui, M. Human keratinocyte-derived extracellular vesicles activate the MAPKinase pathway and promote cell migration and proliferation in vitro. Inflamm. Regen. 2021, 41, 4. [Google Scholar] [CrossRef] [PubMed]
- Monguio-Tortajada, M.; Galvez-Monton, C.; Bayes-Genis, A.; Roura, S.; Borras, F.E. Extracellular vesicle isolation methods: Rising impact of size-exclusion chromatography. Cell Mol. Life Sci. 2019, 76, 2369–2382. [Google Scholar] [CrossRef]
- Peng, C.; Wang, J.; Bao, Q.; Wang, J.; Liu, Z.; Wen, J.; Zhang, W.; Shen, Y. Isolation of extracellular vesicle with different precipitation-based methods exerts a tremendous impact on the biomarker analysis for clinical plasma samples. Cancer Biomark 2020, 29, 373–385. [Google Scholar] [CrossRef]
- Flanagan, J.; Middeldorp, J.; Sculley, T. Localization of the Epstein-Barr virus protein LMP 1 to exosomes. J. Gen. Virol. 2003, 84, 1871–1879. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, N.R.; Chadha, R.; Kumar, S.; Choedon, T.; Reddy, V.S.; Kumar, V. The HBx gene of hepatitis B virus can influence hepatic microenvironment via exosomes by transferring its mRNA and protein. Virus Res. 2017, 240, 166–174. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, X.; Yu, Q.; He, J.J. Exosome-associated hepatitis C virus in cell cultures and patient plasma. Biochem. Biophys. Res. Commun. 2014, 455, 218–222. [Google Scholar] [CrossRef]
- Cone, A.S.; York, S.B.; Meckes, D.G., Jr. Extracellular Vesicles in Epstein-Barr Virus Pathogenesis. Curr. Clin. Microbiol. Rep. 2019, 6, 121–131. [Google Scholar] [CrossRef]
- Meissner, J.D. Nucleotide sequences and further characterization of human papillomavirus DNA present in the CaSki, SiHa and HeLa cervical carcinoma cell lines. J. Gen. Virol. 1999, 80 Pt 7, 1725–1733. [Google Scholar] [CrossRef]
- Kannan, A.; Hertweck, K.L.; Philley, J.V.; Wells, R.B.; Dasgupta, S. Genetic Mutation and Exosome Signature of Human Papilloma Virus Associated Oropharyngeal Cancer. Sci. Rep. 2017, 7, 46102. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.; Marczak, L.; Sharma, P.; Abramowicz, A.; Gawin, M.; Widlak, P.; Whiteside, T.L.; Pietrowska, M. Proteomes of exosomes from HPV(+) or HPV(−) head and neck cancer cells: Differential enrichment in immunoregulatory proteins. Oncoimmunology 2019, 8, 1593808. [Google Scholar] [CrossRef] [PubMed]
- Guzel, C.; Govorukhina, N.I.; Wisman, G.B.A.; Stingl, C.; Dekker, L.J.M.; Klip, H.G.; Hollema, H.; Guryev, V.; Horvatovich, P.L.; van der Zee, A.G.J.; et al. Proteomic alterations in early stage cervical cancer. Oncotarget 2018, 9, 18128–18147. [Google Scholar] [CrossRef] [Green Version]
- Higareda-Almaraz, J.C.; Enriquez-Gasca Mdel, R.; Hernandez-Ortiz, M.; Resendis-Antonio, O.; Encarnacion-Guevara, S. Proteomic patterns of cervical cancer cell lines, a network perspective. BMC Syst. Biol. 2011, 5, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappa, K.I.; Lygirou, V.; Kontostathi, G.; Zoidakis, J.; Makridakis, M.; Vougas, K.; Daskalakis, G.; Polyzos, A.; Anagnou, N.P. Proteomic Analysis of Normal and Cancer Cervical Cell Lines Reveals Deregulation of Cytoskeleton-associated Proteins. Cancer Genom. Proteom. 2017, 14, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zhu, X.; Ren, J.; Huang, S.; Xiao, Z.; Jiang, H.; Tan, Y. Quantitative proteomic analysis of cervical cancer based on TMT-labeled quantitative proteomics. J. Proteomics 2022, 252, 104453. [Google Scholar] [CrossRef]
- Zhu, X.; Lv, J.; Yu, L.; Zhu, X.; Wu, J.; Zou, S.; Jiang, S. Proteomic identification of differentially-expressed proteins in squamous cervical cancer. Gynecol. Oncol. 2009, 112, 248–256. [Google Scholar] [CrossRef]
- D’Arrigo, D.; Roffi, A.; Cucchiarini, M.; Moretti, M.; Candrian, C.; Filardo, G. Secretome and Extracellular Vesicles as New Biological Therapies for Knee Osteoarthritis: A Systematic Review. J. Clin. Med. 2019, 8, 1867. [Google Scholar] [CrossRef] [Green Version]
- Kontostathi, G.; Zoidakis, J.; Makridakis, M.; Lygirou, V.; Mermelekas, G.; Papadopoulos, T.; Vougas, K.; Vlamis-Gardikas, A.; Drakakis, P.; Loutradis, D.; et al. Cervical Cancer Cell Line Secretome Highlights the Roles of Transforming Growth Factor-Beta-Induced Protein ig-h3, Peroxiredoxin-2, and NRF2 on Cervical Carcinogenesis. Biomed. Res. Int. 2017, 2017, 4180703. [Google Scholar] [CrossRef] [Green Version]
- Pappa, K.I.; Kontostathi, G.; Makridakis, M.; Lygirou, V.; Zoidakis, J.; Daskalakis, G.; Anagnou, N.P. High Resolution Proteomic Analysis of the Cervical Cancer Cell Lines Secretome Documents Deregulation of Multiple Proteases. Cancer Genom. Proteom. 2017, 14, 507–521. [Google Scholar] [CrossRef] [Green Version]
- Koli, S.; Labelle-Dumais, C.; Zhao, Y.; Paylakhi, S.; Nair, K.S. Identification of MFRP and the secreted serine proteases PRSS56 and ADAMTS19 as part of a molecular network involved in ocular growth regulation. PLoS Genet. 2021, 17, e1009458. [Google Scholar] [CrossRef]
- Skryabin, G.O.; Komelkov, A.V.; Galetsky, S.A.; Bagrov, D.V.; Evtushenko, E.G.; Nikishin, I.I.; Zhordaniia, K.I.; Savelyeva, E.E.; Akselrod, M.E.; Paianidi, I.G.; et al. Stomatin is highly expressed in exosomes of different origin and is a promising candidate as an exosomal marker. J. Cell Biochem. 2021, 122, 100–115. [Google Scholar] [CrossRef] [PubMed]
- Skryabin, G.O.; Vinokurova, S.V.; Galetsky, S.A.; Elkin, D.S.; Senkovenko, A.M.; Denisova, D.A.; Komelkov, A.V.; Stilidi, I.S.; Peregorodiev, I.N.; Malikhova, O.A.; et al. Isolation and Characterization of Extracellular Vesicles from Gastric Juice. Cancers 2022, 14, 3314. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhao, H.; Li, X.M.; Yin, C.H.; Wu, Y.M. Use of mesothelin as a tumor-associated antigen in cervical squamous cell carcinoma. Gene 2019, 690, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chao, A.; Wang, T.H.; Lee, Y.S.; Hsueh, S.; Chao, A.S.; Chang, T.C.; Kung, W.H.; Huang, S.L.; Chao, F.Y.; Wei, M.L.; et al. Molecular characterization of adenocarcinoma and squamous carcinoma of the uterine cervix using microarray analysis of gene expression. Int. J. Cancer 2006, 119, 91–98. [Google Scholar] [CrossRef]
- Johrens, K.; Lazzerini, L.; Barinoff, J.; Sehouli, J.; Cichon, G. Mesothelin as a target for cervical cancer therapy. Arch. Gynecol. Obstet. 2019, 299, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Carter, S.A.; Foster, N.A.; Scarpini, C.G.; Chattopadhyay, A.; Pett, M.R.; Roberts, I.; Coleman, N. Lipoprotein lipase is frequently overexpressed or translocated in cervical squamous cell carcinoma and promotes invasiveness through the non-catalytic C terminus. Br. J. Cancer 2012, 107, 739–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancusi de Carvalho, J.P.; Salim, R.C.; Carvalho, F.M.; Nogueira Dias Genta, M.L.; Baracat, E.C.; Carvalho, J.P. L1 cell adhesion molecule (L1CAM) in stage IB cervical cancer: Distinct expression in squamous cell carcinomas and adenocarcinomas. J. Clin. Pathol. 2020, 73, 748–753. [Google Scholar] [CrossRef] [PubMed]
- Schrevel, M.; Corver, W.E.; Vegter, M.E.; Ter Haar, N.T.; Dreef, E.J.; Beltman, J.J.; Kenter, G.; Bosse, T.; de Kroon, C.D.; Jordanova, E.S. L1 cell adhesion molecule (L1CAM) is a strong predictor for locoregional recurrences in cervical cancer. Oncotarget 2017, 8, 87568–87581. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Yuan, J.; Rothzerg, E.; Wu, X.; Xu, H.; Zhu, S.; Xu, J. Molecular structure and the role of high-temperature requirement protein 1 in skeletal disorders and cancers. Cell Prolif. 2020, 53, e12746. [Google Scholar] [CrossRef] [Green Version]
- Stuqui, B.; Conceicao, A.L.; Termini, L.; Sichero, L.; Villa, L.L.; Rahal, P.; Calmon, M.F. The differential role of HTRA1 in HPV-positive and HPV-negative cervical cell line proliferation. BMC Cancer 2016, 16, 840. [Google Scholar] [CrossRef] [Green Version]
- Van Kilsdonk, J.W.; van Kempen, L.C.; van Muijen, G.N.; Ruiter, D.J.; Swart, G.W. Soluble adhesion molecules in human cancers: Sources and fates. Eur. J. Cell Biol. 2010, 89, 415–427. [Google Scholar] [CrossRef]
- Jimenez, L.; Yu, H.; McKenzie, A.J.; Franklin, J.L.; Patton, J.G.; Liu, Q.; Weaver, A.M. Quantitative Proteomic Analysis of Small and Large Extracellular Vesicles (EVs) Reveals Enrichment of Adhesion Proteins in Small EVs. J. Proteome Res. 2019, 18, 947–959. [Google Scholar] [CrossRef]
- Shimoda, M. Extracellular vesicle-associated MMPs: A modulator of the tissue microenvironment. Adv. Clin. Chem. 2019, 88, 35–66. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, M.; Khokha, R. Proteolytic factors in exosomes. Proteomics 2013, 13, 1624–1636. [Google Scholar] [CrossRef] [PubMed]
- Wieckowski, E.U.; Visus, C.; Szajnik, M.; Szczepanski, M.J.; Storkus, W.J.; Whiteside, T.L. Tumor-derived microvesicles promote regulatory T cell expansion and induce apoptosis in tumor-reactive activated CD8+ T lymphocytes. J. Immunol. 2009, 183, 3720–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuliano, M.; Mangino, G.; Chiantore, M.V.; Zangrillo, M.S.; Accardi, R.; Tommasino, M.; Fiorucci, G.; Romeo, G. Human Papillomavirus E6 and E7 oncoproteins affect the cell microenvironment by classical secretion and extracellular vesicles delivery of inflammatory mediators. Cytokine 2018, 106, 182–189. [Google Scholar] [CrossRef]
- Zhang, J.; Burn, C.; Young, K.; Wilson, M.; Ly, K.; Budhwani, M.; Tschirley, A.; Braithwaite, A.; Baird, M.; Hibma, M. Microparticles produced by human papillomavirus type 16 E7-expressing cells impair antigen presenting cell function and the cytotoxic T cell response. Sci. Rep. 2018, 8, 2373. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Wang, Y.; Yuan, S.; Wang, B. Dendritic cells loaded with HeLa-derived exosomes simulate an antitumor immune response. Oncol. Lett. 2018, 15, 6636–6640. [Google Scholar] [CrossRef]
- Van der Watt, P.J.; Okpara, M.O.; Wishart, A.; Parker, M.I.; Soares, N.C.; Blackburn, J.M.; Leaner, V.D. Nuclear transport proteins are secreted by cancer cells and identified as potential novel cancer biomarkers. Int. J. Cancer 2022, 150, 347–361. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position in the Top 100 | Gene Symbol | Protein Name | Average LFQ HeLa’s EVs | Average LFQ HaCaT’s EVs |
|---|---|---|---|---|
| 1 | PDCD6IP | Programmed cell death 6 interacting protein | 30.90126 | 31.82523 |
| 2 | GAPDH | Glyceraldehyde-3-phosphate dehydrogenase | 34.12177 | 34.66822 |
| 3 | HSPA8 | Heat-shock 70kDa protein 8 | 33.61965 | 34.98863 |
| 4 | ACTB | Actin, cytoplasmic 1 | 36.35891 | 36.49115 |
| 5 | ANXA2 | Annexin A2 | 32.45719 | 35.18595 |
| 6 | CD9 | CD9 antigen | 30.40349 | 30.71279 |
| 7 | PKM | Pyruvate kinase PKM | 33.28324 | 34.29517 |
| 8 | HSP90AA1 | Heat-shock protein HSP 90-alpha | 34.42612 | 34.92483 |
| 9 | ENO1 | Alpha-enolase | 33.79492 | 33.82500 |
| 10 | ANXA5 | Annexin A5 | 31.05360 | 31.50710 |
| 11 | HSP90AB1 | Heat-shock protein HSP 90-beta | 33.63702 | 33.91836 |
| 12 | CD63 | CD63 antigen | 29.42370 | 29.26211 |
| 13 | YWHAZ | 14-3-3 Protein zeta/delta | 31.30615 | 32.74208 |
| 14 | YWHAE | 14-3-3 Protein epsilon | 30.73296 | 31.84702 |
| 15 | EEF1A1 | Elongation factor 1 alpha 1 | 32.55905 | 33.68600 |
| 16 | PGK1 | Phosphoglycerate kinase 1 | 31.74001 | 31.45923 |
| 17 | CLTC | Clathrin heavy chain 1 | 26.95375 | 28.79626 |
| 18 | PPIA | Peptidyl-prolyl cis-trans isomerase A | 31.76701 | 32.57919 |
| 19 | SDCBP | Syntenin-1 | 30.96517 | 32.68455 |
| 20 | ALDOA | Fructose-bisphosphate aldolase A | 32.53601 | 33.17732 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Acevedo-Sánchez, V.; Martínez-Ruiz, R.S.; Aguilar-Ruíz, S.R.; Torres-Aguilar, H.; Chávez-Olmos, P.; Garrido, E.; Baltiérrez-Hoyos, R.; Romero-Tlalolini, M.d.l.A. Quantitative Proteomics for the Identification of Differentially Expressed Proteins in the Extracellular Vesicles of Cervical Cancer Cells. Viruses 2023, 15, 702. https://doi.org/10.3390/v15030702
Acevedo-Sánchez V, Martínez-Ruiz RS, Aguilar-Ruíz SR, Torres-Aguilar H, Chávez-Olmos P, Garrido E, Baltiérrez-Hoyos R, Romero-Tlalolini MdlA. Quantitative Proteomics for the Identification of Differentially Expressed Proteins in the Extracellular Vesicles of Cervical Cancer Cells. Viruses. 2023; 15(3):702. https://doi.org/10.3390/v15030702
Chicago/Turabian StyleAcevedo-Sánchez, Víctor, Roy S. Martínez-Ruiz, Sergio R. Aguilar-Ruíz, Honorio Torres-Aguilar, Pedro Chávez-Olmos, Efraín Garrido, Rafael Baltiérrez-Hoyos, and María de los A. Romero-Tlalolini. 2023. "Quantitative Proteomics for the Identification of Differentially Expressed Proteins in the Extracellular Vesicles of Cervical Cancer Cells" Viruses 15, no. 3: 702. https://doi.org/10.3390/v15030702
APA StyleAcevedo-Sánchez, V., Martínez-Ruiz, R. S., Aguilar-Ruíz, S. R., Torres-Aguilar, H., Chávez-Olmos, P., Garrido, E., Baltiérrez-Hoyos, R., & Romero-Tlalolini, M. d. l. A. (2023). Quantitative Proteomics for the Identification of Differentially Expressed Proteins in the Extracellular Vesicles of Cervical Cancer Cells. Viruses, 15(3), 702. https://doi.org/10.3390/v15030702