
Fucose Binding Cancels out Mechanical Differences between Distinct Human Noroviruses

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. NoroVLP Production and Purification
2.2. AFM Sample Preparation
2.3. Fucose Treatment
2.4. AFM Imaging and Nanoindentation
2.5. Data Processing
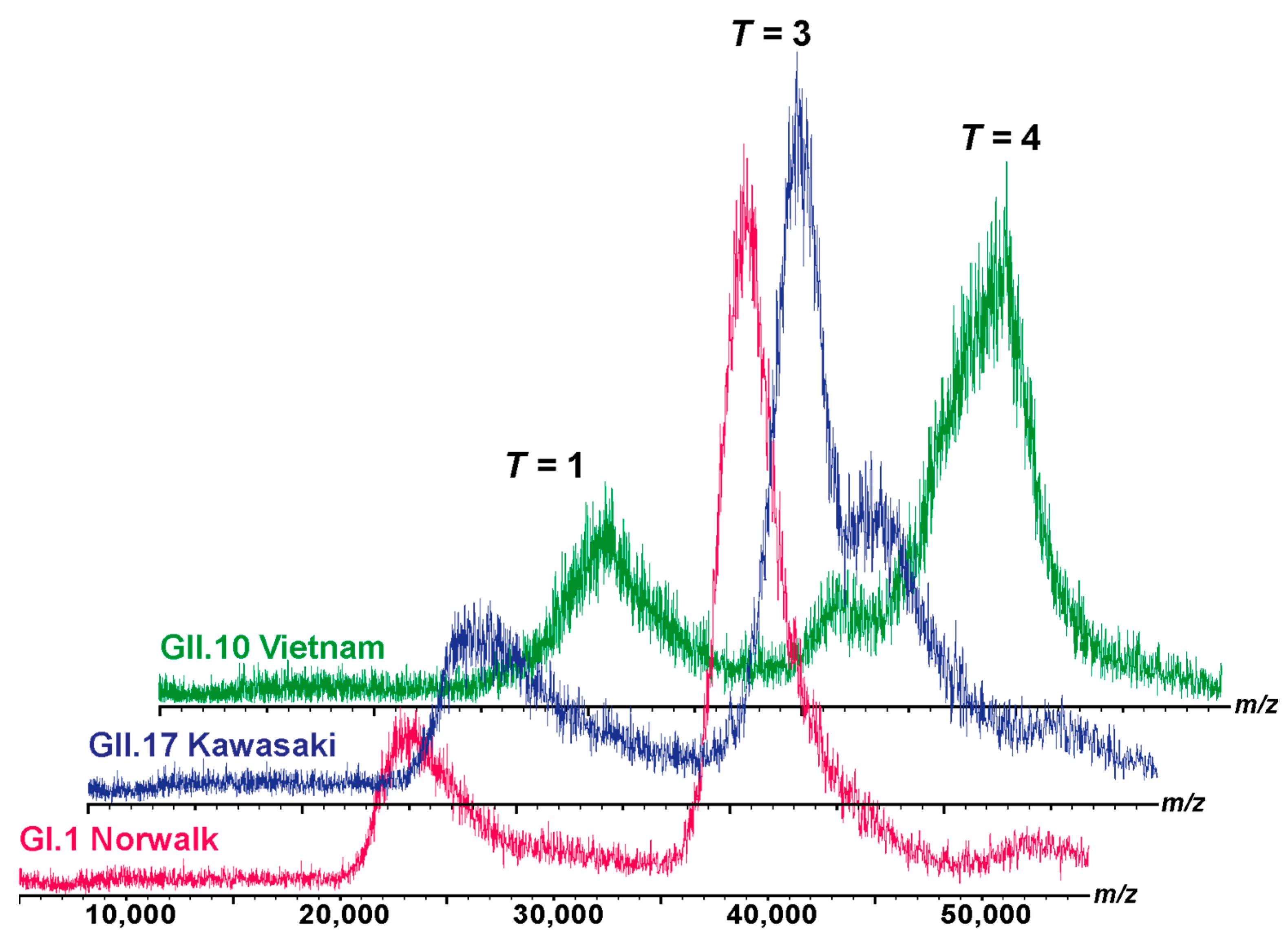
2.6. Native Mass Spectrometry
3. Results
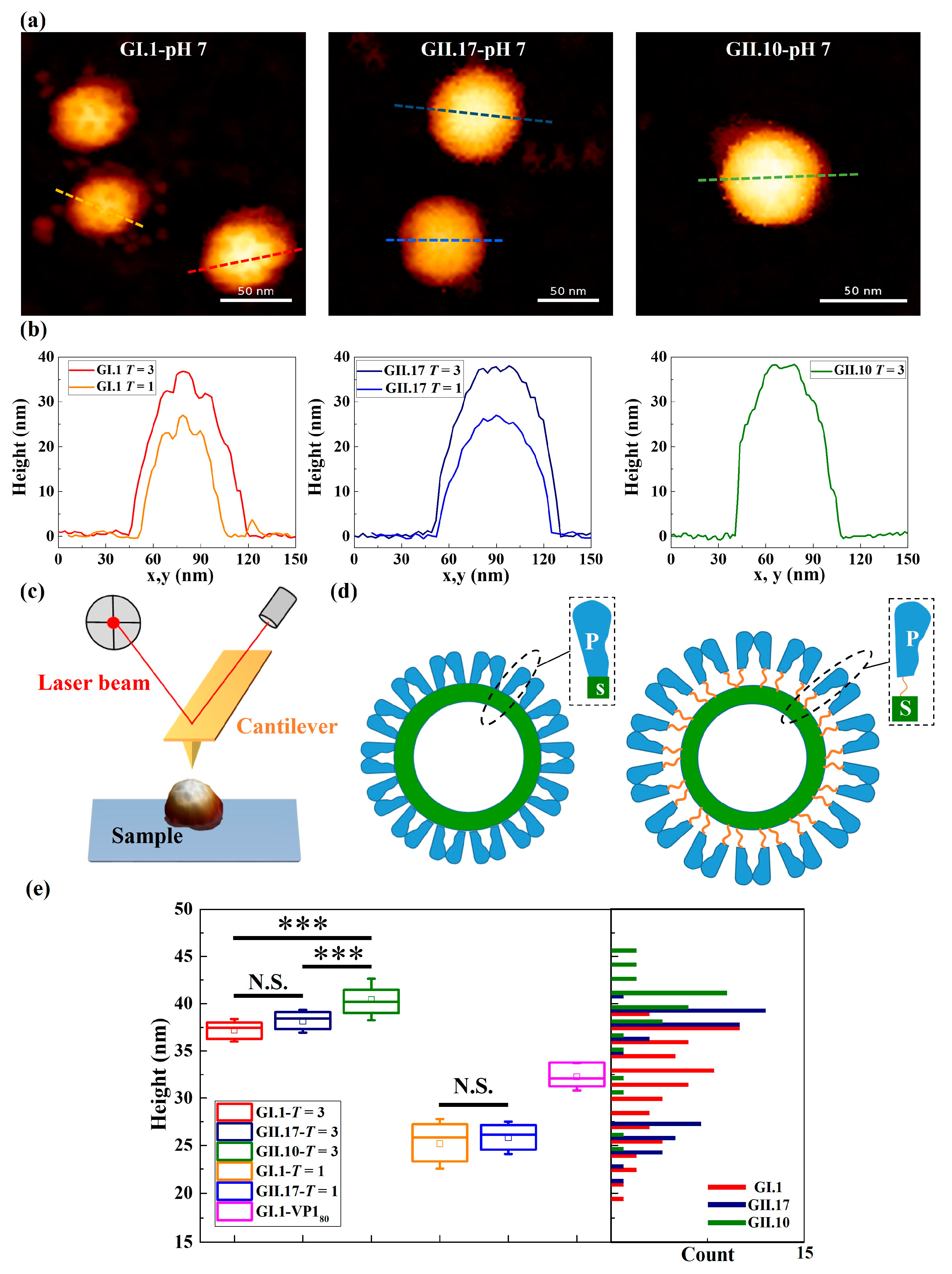
3.1. Different Types of Assemblies and Sizes among Three Variants under Neutral Condition
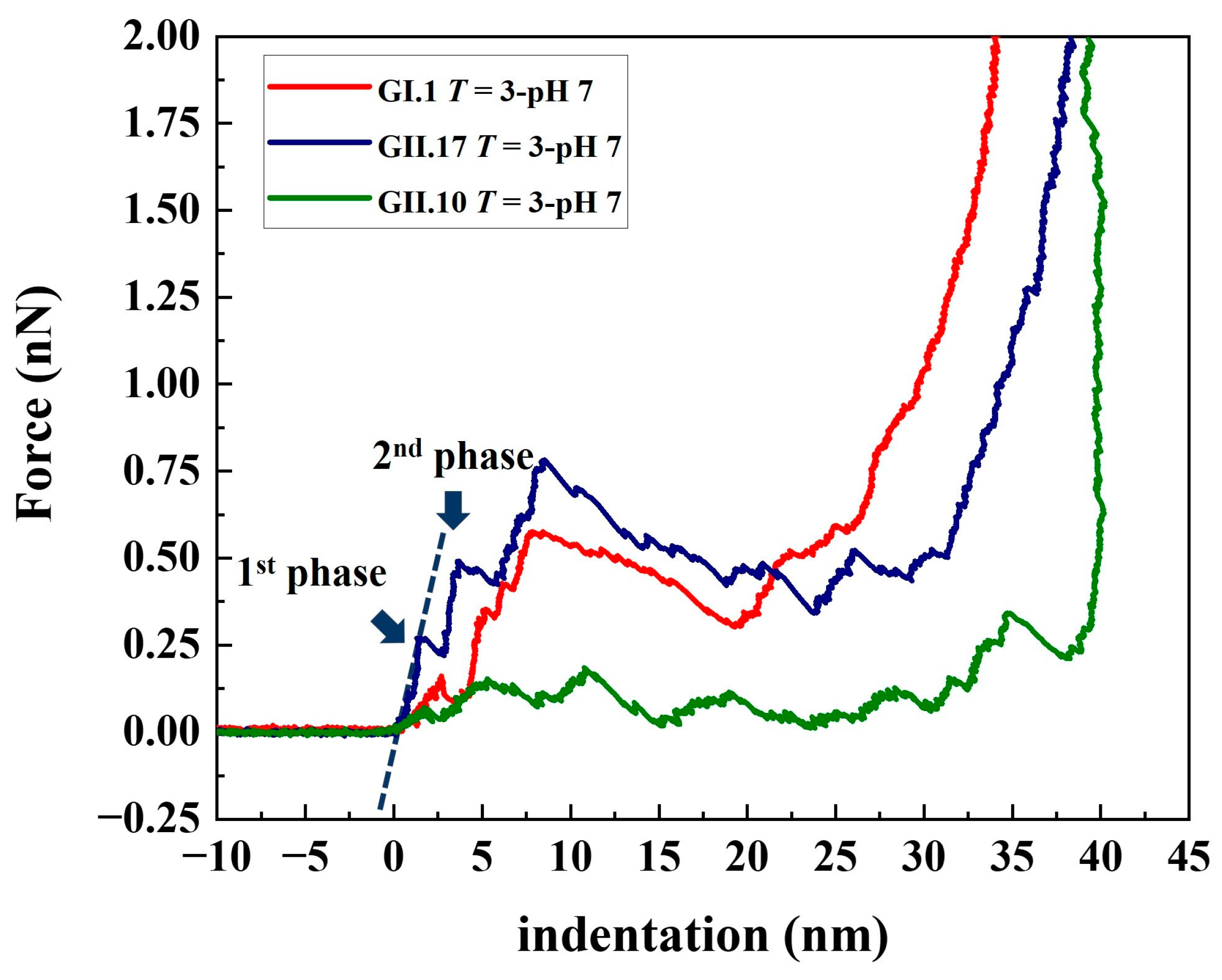
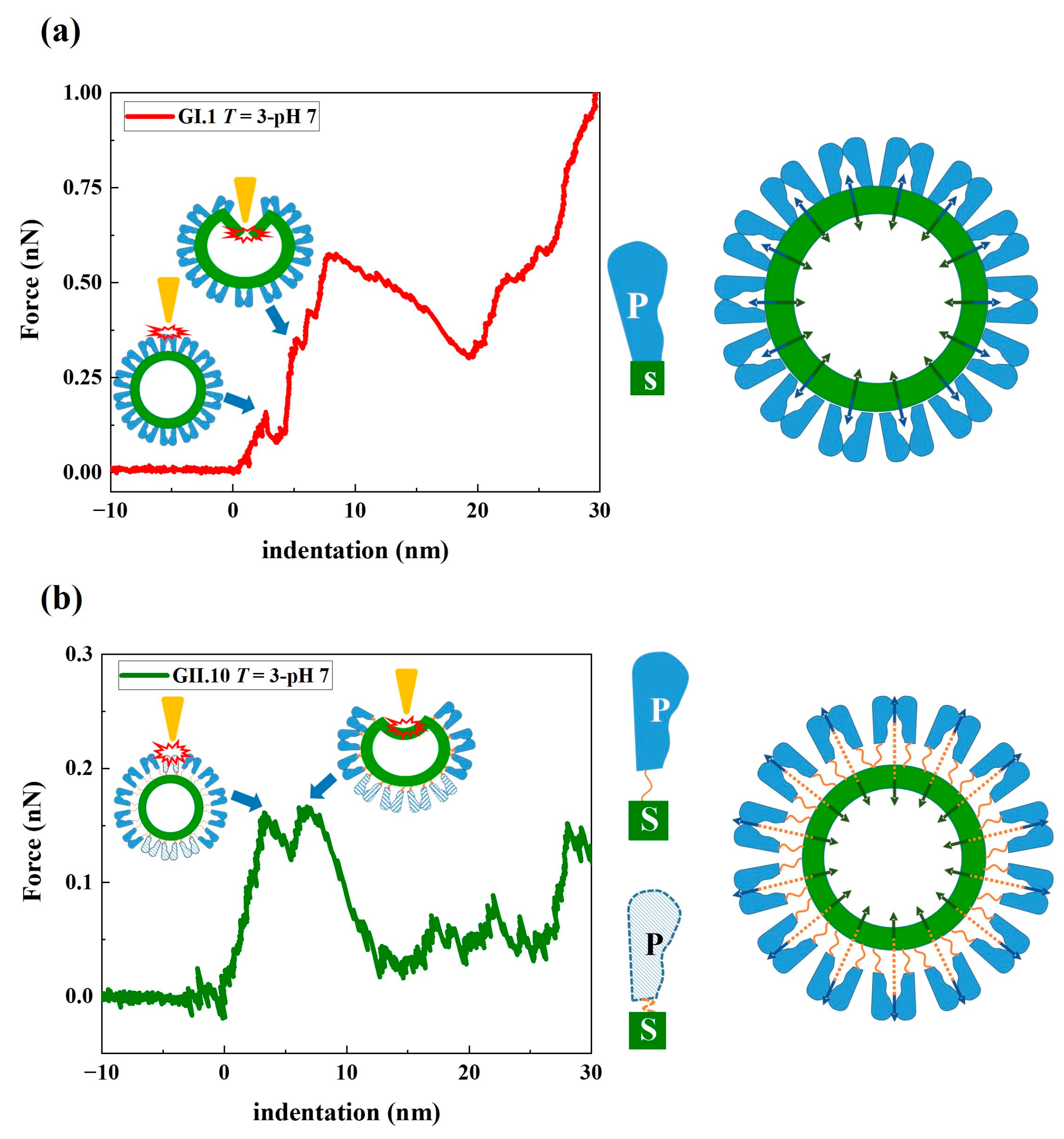
3.2. Stepwise Deformation of noroVLPs under Compression
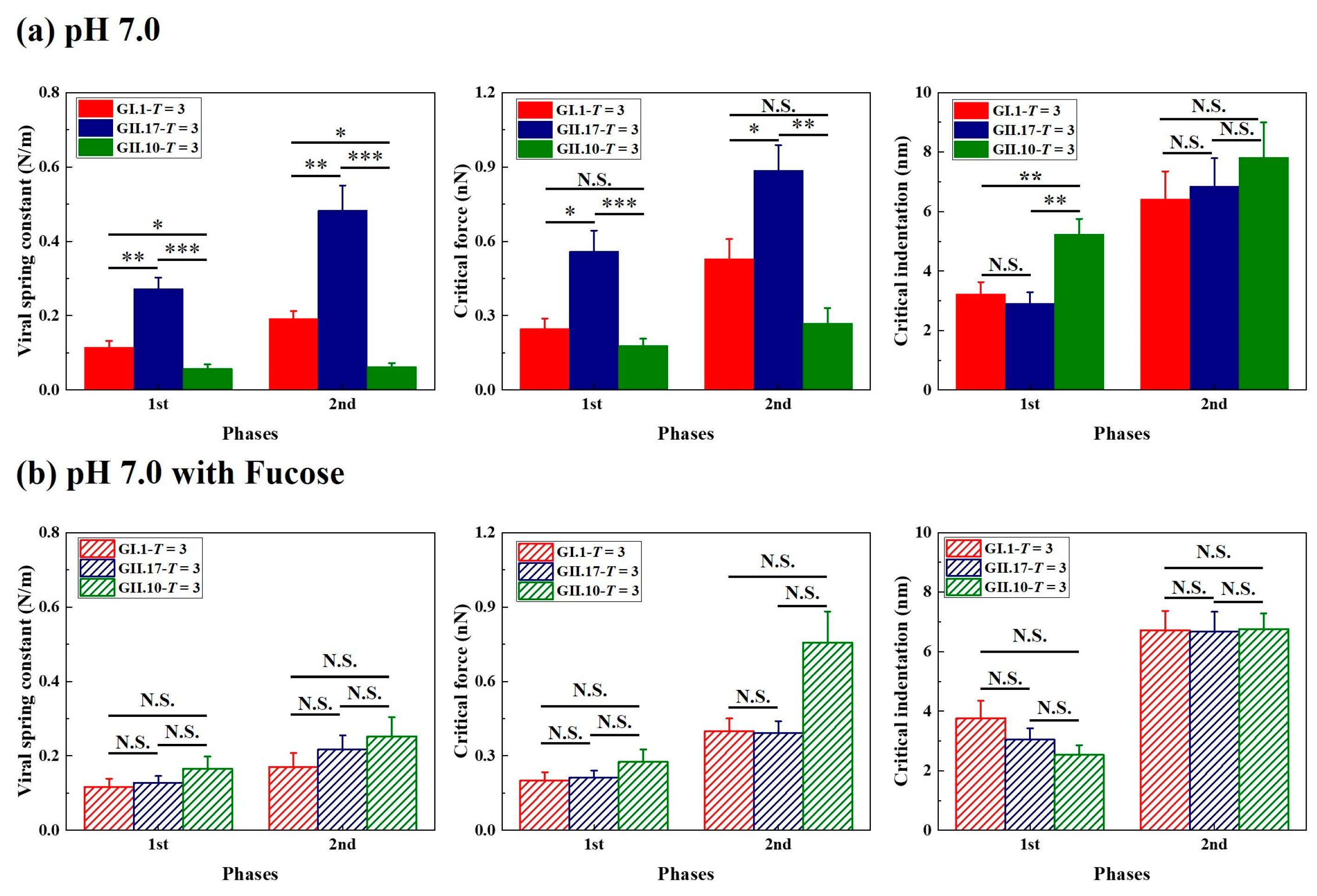
3.3. Different Mechanical Properties under Neutral Condition
3.4. Fucose Treatment Diminished Original Differences
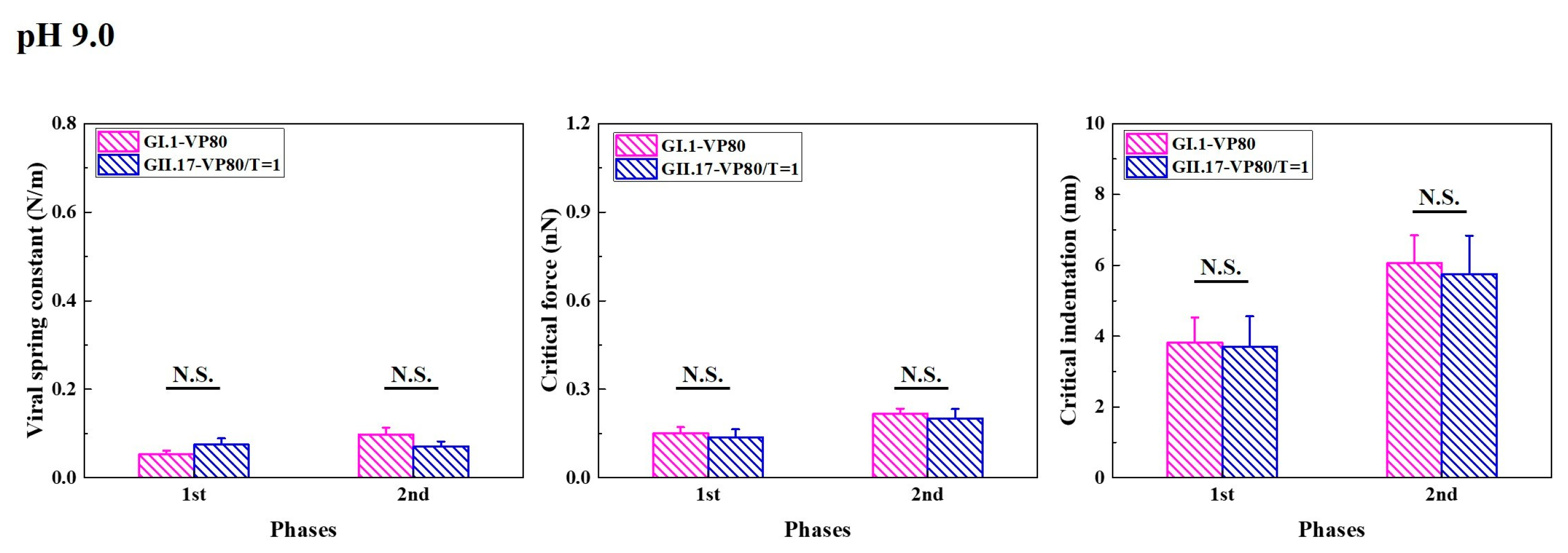
3.5. Alkaline Treatment Weakened Mechanics of noroVLPs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xi, J.A.; Graham, D.Y.; Wang, K.N.; Estes, M.K. Norwalk virus genome cloning and characterization. Science 1990, 250, 1580–1583. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.; Hall, A.J.; Robinson, A.E.; Verhoef, L.; Premkumar, P.; Parashar, U.D.; Koopmans, M.; Lopman, B.A. Global prevalence of norovirus in cases of gastroenteritis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 725–730. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, M.; van Beek, J.; Koopmans, M.P.G. Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 2016, 14, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Green, K.Y.; Kaufman, S.S.; Nagata, B.M.; Chaimongkol, N.; Kim, D.Y.; Levenson, E.A.; Tin, C.M.; Yardley, A.B.; Johnson, J.A.; Barletta, A.B.F.; et al. Human norovirus targets enteroendocrine epithelial cells in the small intestine. Nat. Commun. 2020, 11, 2759. [Google Scholar] [CrossRef]
- Tan, M.; Jiang, X. Norovirus and its histo-blood group antigen receptors: An answer to a historical puzzle. Trends Microbiol. 2005, 13, 285–293. [Google Scholar] [CrossRef]
- Koromyslova, A.D.; Leuthold, M.M.; Bowler, M.W.; Hansman, G.S. The sweet quartet: Binding of fucose to the norovirus capsid. Virology 2015, 483, 203–208. [Google Scholar] [CrossRef]
- Orchard, R.C.; Wilen, C.B.; Doench, J.G.; Baldridge, M.T.; McCune, B.T.; Lee, Y.-C.J.; Lee, S.; Pruett-Miller, S.M.; Nelson, C.A.; Fremont, D.H.; et al. Discovery of a proteinaceous cellular receptor for a norovirus. Science 2016, 353, 933–936. [Google Scholar] [CrossRef]
- Graziano, V.R.; Wei, J.; Wilen, C.B. Norovirus Attachment and Entry. Viruses 2019, 11, 495. [Google Scholar] [CrossRef]
- Chhabra, P.; de Graaf, M.; Parra, G.I.; Chan, M.C.-W.; Green, K.; Martella, V.; Wang, Q.; White, P.A.; Katayama, K.; Vennema, H.; et al. Updated classification of norovirus genogroups and genotypes. J. Gen. Virol. 2019, 100, 1393–1406. [Google Scholar] [CrossRef]
- de Graaf, M.; van Beek, J.; Vennema, H.; Podkolzin, A.T.; Hewitt, J.; Bucardo, F.; Templeton, K.; Mans, J.; Nordgren, J.; Reuter, G.; et al. Emergence of a novel GII.17 norovirus-End of the GII.4 era? Eurosurveillance 2015, 20, 8–15. [Google Scholar] [CrossRef]
- Pogan, R.; Schneider, C.; Reimer, R.; Hansman, G.; Uetrecht, C. Norovirus-like VP1 particles exhibit isolate dependent stability profiles. J. Phys.-Condens. Matter 2018, 30, 064006. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Tan, M.; Xia, M.; Wei, C.; Huang, P.W.; Wang, L.Y.; Zhong, W.M.; Duan, Z.J.; Jiang, X. Strain-specific interaction of a GII.10 Norovirus with HBGAs. Virology 2015, 476, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.A.; Min, W.; Graham, D.Y.; Estes, M.K. Expression, self-assembly, and antigenicity of the norwalk virus capsid protein. J. Virol. 1992, 66, 6527–6532. [Google Scholar] [CrossRef]
- Prasad, B.V.V.; Hardy, M.E.; Dokland, T.; Bella, J.; Rossmann, M.G.; Estes, M.K. X-ray crystallographic structure of the Norwalk virus capsid. Science 1999, 286, 287–290. [Google Scholar] [CrossRef]
- Hansman, G.S.; Taylor, D.W.; McLellan, J.S.; Smith, T.J.; Georgiev, I.; Tame, J.R.H.; Park, S.Y.; Yamazaki, M.; Gondaira, F.; Miki, M.; et al. Structural basis for broad detection of genogroup II noroviruses by a monoclonal antibody that binds to a site occluded in the viral particle. J. Virol. 2012, 86, 3635–3646. [Google Scholar] [CrossRef]
- Song, C.H.; Takai-Todaka, R.; Miki, M.; Haga, K.; Fujimoto, A.; Ishiyama, R.; Oikawa, K.; Yokoyama, M.; Miyazaki, N.; Iwasaki, K.; et al. Dynamic rotation of the protruding domain enhances the infectivity of norovirus. PLoS Pathog. 2020, 16, e1008619. [Google Scholar] [CrossRef]
- Smith, H.Q.; Smith, T.J. The dynamic capsid structures of the noroviruses. Viruses 2019, 11, 235. [Google Scholar] [CrossRef]
- Hu, L.; Salmen, W.; Chen, R.; Zhou, Y.; Neill, F.; Crowe, J.E.; Atmar, R.L.; Estes, M.K.; Prasad, B.V.V. Atomic structure of the predominant GII.4 human norovirus capsid reveals novel stability and plasticity. Nat. Commun. 2022, 13, 1241. [Google Scholar] [CrossRef]
- Storch, G.A. Diagnostic Virology. Clin. Infect. Dis. 2000, 31, 739–751. [Google Scholar] [CrossRef]
- Udugama, B.; Kadhiresan, P.; Kozlowski, H.N.; Malekjahani, A.; Osborne, M.; Li, V.Y.C.; Chen, H.; Mubareka, S.; Gubbay, J.B.; Chan, W.C.W. Diagnosing COVID-19: The Disease and Tools for Detection. ACS Nano 2020, 14, 3822–3835. [Google Scholar] [CrossRef]
- Roingeard, P.; Raynal, P.-I.; Eymieux, S.; Blanchard, E. Virus detection by transmission electron microscopy: Still useful for diagnosis and a plus for biosafety. Rev. Med. Virol. 2019, 29, e2019. [Google Scholar] [CrossRef] [PubMed]
- Roos, W.H.; Bruinsma, R.; Wuite, G.J.L. Physical virology. Nat. Phys. 2010, 6, 733–743. [Google Scholar] [CrossRef]
- Ortega-Esteban, Á.; Martín-González, N.; Moreno-Madrid, F.; Llauró, A.; Hernando-Pérez, M.; MartÚn, C.S.; de Pablo, P.J. Structural and Mechanical Characterization of Viruses with AFM. In Atomic Force Microscopy: Methods and Protocols; Santos, N.C., Carvalho, F.A., Eds.; Springer: New York, NY, USA, 2019; pp. 259–278. [Google Scholar]
- Buzón, P.; Maity, S.; Roos, W.H. Physical virology: From virus self-assembly to particle mechanics. WIREs Nanomed. Nanobiotechnol. 2020, 12, e1613. [Google Scholar] [CrossRef] [PubMed]
- Dülfer, J.; Kadek, A.; Kopicki, J.-D.; Krichel, B.; Uetrecht, C. Chapter Seven-Structural mass spectrometry goes viral. In Advances in Virus Research; Rey, F.A., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 105, pp. 189–238. [Google Scholar]
- Leney, A.C.; Heck, A.J.R. Native Mass Spectrometry: What is in the Name? J. Am. Soc. Mass Spectrom. 2017, 28, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Hansman, G.S.; Natori, K.; Oka, T.; Ogawa, S.; Tanaka, K.; Nagata, N.; Ushijima, H.; Takeda, N.; Katayama, K. Cross-reactivity among sapovirus recombinant capsid proteins. Arch. Virol. 2005, 150, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Hansman, G.S.; Saito, H.; Shibata, C.; Ishizuka, S.; Oseto, M.; Oka, T.; Takeda, N. Outbreak of gastroenteritis due to sapovirus. J. Clin. Microbiol. 2007, 45, 1347–1349. [Google Scholar] [CrossRef]
- Ivanovska, I.L.; de Pablo, P.J.; Ibarra, B.; Sgalari, G.; MacKintosh, F.C.; Carrascosa, J.L.; Schmidt, C.F.; Wuite, G.J.L. Bacteriophage capsids: Tough nanoshells with complex elastic properties. Proc. Natl. Acad. Sci. USA 2004, 101, 7600–7605. [Google Scholar] [CrossRef]
- Guo, Y.; Roos, W.H. AFM nanoindentation experiments on protein shells: A protocol. In Atomic Force Microscopy: Methods and Protocols; Santos, N.C., Carvalho, F.A., Eds.; Springer: New York, NY, USA, 2019; pp. 243–257. [Google Scholar]
- Han, L.; Kitov, P.I.; Kitova, E.N.; Tan, M.; Wang, L.; Xia, M.; Jiang, X.; Klassen, J.S. Affinities of recombinant norovirus P dimers for human blood group antigens. Glycobiology 2013, 23, 276–285. [Google Scholar] [CrossRef]
- Hansman Grant, S.; Shahzad-ul-Hussan, S.; McLellan Jason, S.; Chuang, G.-Y.; Georgiev, I.; Shimoike, T.; Katayama, K.; Bewley Carole, A.; Kwong Peter, D. Structural Basis for Norovirus Inhibition and Fucose Mimicry by Citrate. J. Virol. 2012, 86, 284–292. [Google Scholar] [CrossRef]
- Mallagaray, A.; Creutznacher, R.; Dülfer, J.; Mayer, P.H.O.; Grimm, L.L.; Orduña, J.M.; Trabjerg, E.; Stehle, T.; Rand, K.D.; Blaum, B.S.; et al. A post-translational modification of human Norovirus capsid protein attenuates glycan binding. Nat. Commun. 2019, 10, 1320. [Google Scholar] [CrossRef]
- Peters, T.; Creutznacher, R.; Maass, T.; Mallagaray, A.; Ogrissek, P.; Taube, S.; Thiede, L.; Uetrecht, C. Norovirus–glycan interactions—how strong are they really? Biochem. Soc. Trans. 2021, 50, 347–359. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, R.H.H.; van Duijn, E.; Mazon, H.; Synowsky, S.A.; Lorenzen, K.; Versluis, C.; Brouns, S.J.J.; Langridge, D.; van der Oost, J.; Hoyes, J.; et al. Improving the Performance of a Quadrupole Time-of-Flight Instrument for Macromolecular Mass Spectrometry. Anal. Chem. 2006, 78, 7473–7483. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, K.; Versluis, C.; van Duijn, E.; van den Heuvel, R.H.H.; Heck, A.J.R. Optimizing macromolecular tandem mass spectrometry of large non-covalent complexes using heavy collision gases. Int. J. Mass Spectrom. 2007, 268, 198–206. [Google Scholar] [CrossRef]
- Shoemaker, G.K.; van Duijn, E.; Crawford, S.E.; Uetrecht, C.; Baclayon, M.; Roos, W.H.; Wuite, G.J.L.; Estes, M.K.; Prasad, B.V.V.; Heck, A.J.R. Norwalk virus assembly and stability monitored by mass spectrometry. Mol. Cell Proteom. 2010, 9, 1742–1751. [Google Scholar] [CrossRef]
- Baclayon, M.; Shoemaker, G.K.; Uetrecht, C.; Crawford, S.E.; Estes, M.K.; Prasad, B.V.V.; Heck, A.J.R.; Wuite, G.J.L.; Roos, W.H. Prestress strengthens the shell of norwalk virus nanoparticles. Nano Lett. 2011, 11, 4865–4869. [Google Scholar] [CrossRef]
- Heck, A.J.R.; van den Heuvel, R.H.H. Investigation of intact protein complexes by mass spectrometry. Mass Spectrom. Rev. 2004, 23, 368–389. [Google Scholar] [CrossRef]
- Lorenzen, K.; Olia, A.S.; Uetrecht, C.; Cingolani, G.; Heck, A.J.R. Determination of Stoichiometry and Conformational Changes in the First Step of the P22 Tail Assembly. J. Mol. Biol. 2008, 379, 385–396. [Google Scholar] [CrossRef]
- Jung, J.; Grant, T.; Thomas, D.R.; Diehnelt, C.W.; Grigorieff, N.; Joshua-Tor, L. High-resolution cryo-EM structures of outbreak strain human norovirus shells reveal size variations. Proc. Natl. Acad. Sci. USA 2019, 116, 12828–12832. [Google Scholar] [CrossRef]
- Boyd, K.J.; Bansal, P.; Feng, J.; May, E.R. Stability of norwalk virus capsid protein interfaces evaluated by in silico nanoindentation. Front. Bioeng. Biotechnol. 2015, 3, 103. [Google Scholar] [CrossRef]
- Müller, B.; Nübling, C.M.; Kress, J.; Roth, W.K.; De Zolt, S.; Pichl, L. How safe is safe: New human immunodeficiency virus Type 1 variants missed by nucleic acid testing. Transfusion 2013, 53, 2422–2430. [Google Scholar] [CrossRef]
- Cuellar, J.L.; Meinhoevel, F.; Hoehne, M.; Donath, E. Size and mechanical stability of norovirus capsids depend on pH: A nanoindentation study. J. Gen. Virol. 2010, 91, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.M.; Hutson, A.M.; Estes, M.K.; Prasad, B.V.V. Atomic resolution structural characterization of recognition of histo-blood group antigens by norwalk virus. Proc. Natl. Acad. Sci. USA 2008, 105, 9175–9180. [Google Scholar] [CrossRef] [PubMed]
- Dulfer, J.; Yan, H.; Brodmerkel, M.N.; Creutznacher, R.; Mallagaray, A.; Peters, T.; Caleman, C.; Marklund, E.G.; Uetrecht, C. Glycan-Induced protein dynamics in human norovirus P dimers depend on virus strain and deamidation status. Molecules 2021, 26, 2125. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Y.; Pogan, R.; Thiede, L.; Müller-Guhl, J.; Uetrecht, C.; Roos, W.H. Fucose Binding Cancels out Mechanical Differences between Distinct Human Noroviruses. Viruses 2023, 15, 1482. https://doi.org/10.3390/v15071482
Feng Y, Pogan R, Thiede L, Müller-Guhl J, Uetrecht C, Roos WH. Fucose Binding Cancels out Mechanical Differences between Distinct Human Noroviruses. Viruses. 2023; 15(7):1482. https://doi.org/10.3390/v15071482
Chicago/Turabian StyleFeng, Yuzhen, Ronja Pogan, Lars Thiede, Jürgen Müller-Guhl, Charlotte Uetrecht, and Wouter H. Roos. 2023. "Fucose Binding Cancels out Mechanical Differences between Distinct Human Noroviruses" Viruses 15, no. 7: 1482. https://doi.org/10.3390/v15071482
APA StyleFeng, Y., Pogan, R., Thiede, L., Müller-Guhl, J., Uetrecht, C., & Roos, W. H. (2023). Fucose Binding Cancels out Mechanical Differences between Distinct Human Noroviruses. Viruses, 15(7), 1482. https://doi.org/10.3390/v15071482