Effect of High Hydrostatic Pressure Processing and Holder Pasteurization of Human Milk on Inactivation of Human Coronavirus 229E and Hepatitis E Virus

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Milk Samples, HHP and HoP Treatments
2.2. Cells and Plasmids
2.3. Virus Production
2.4. Infectious Titers and Quantification of Viral RNA in Infected Cells
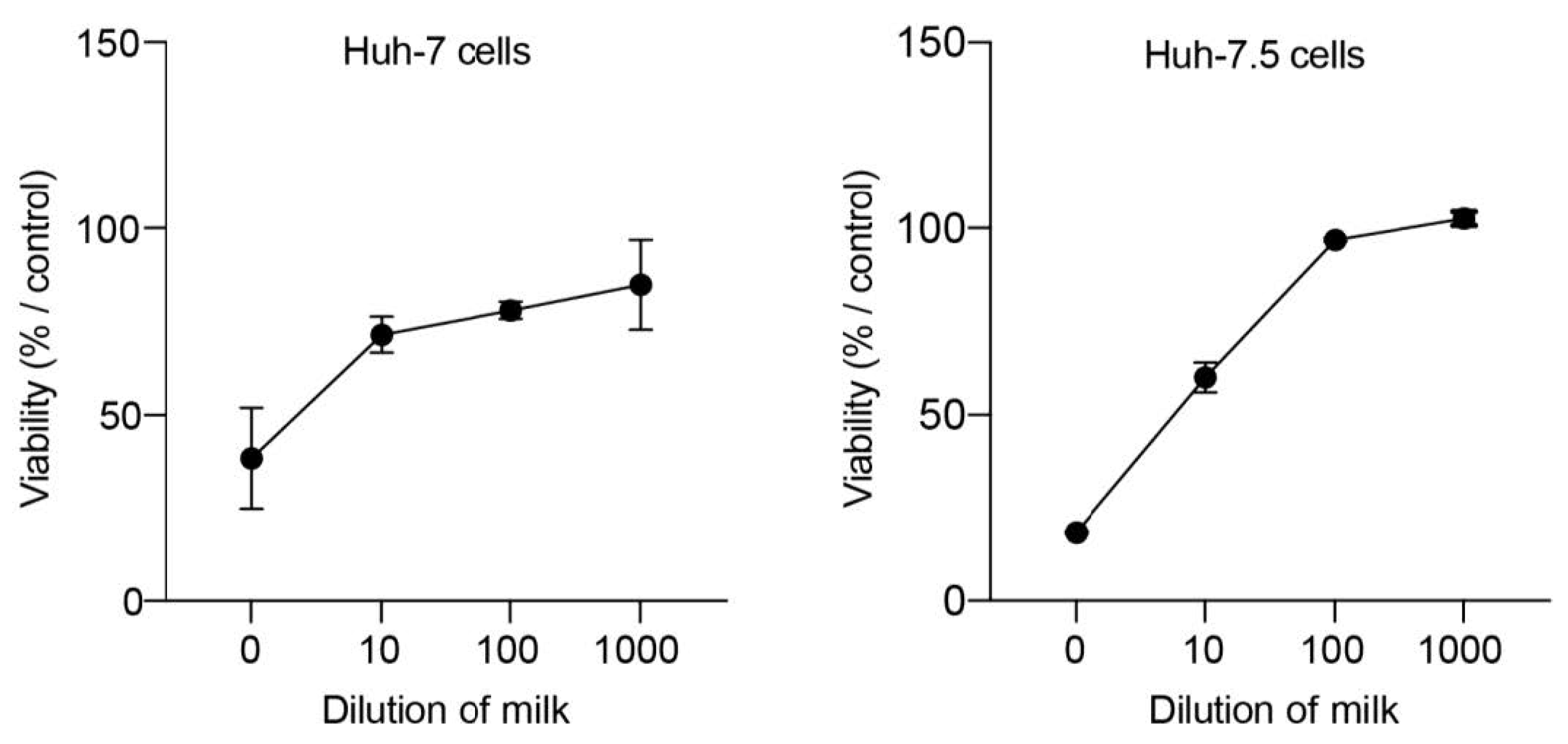
2.5. Cellular Toxicity Assay
2.6. Statistical Analysis
3. Results
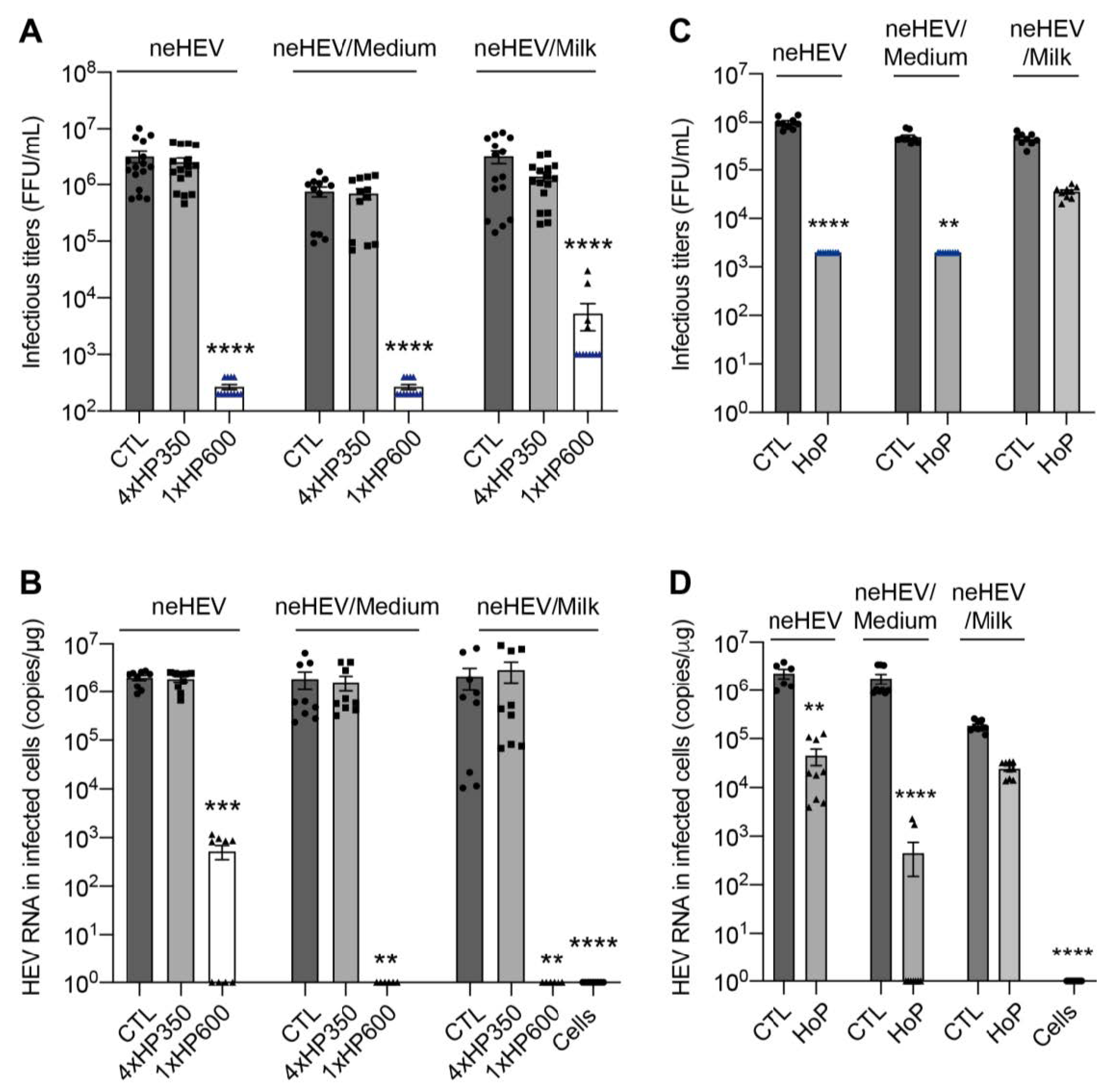
3.1. HHP and HoP Processing of Human Milk Infected with HCoV-229E, eHEV, or neHEV Viruses
3.2. HHP and HoP Processing of Human Milk Inoculated with HCoV-229E
3.3. HHP and HoP Processing of Human Milk Inoculated with eHEV
3.4. HHP and HoP Processing of Human Milk Inoculated with neHEV
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Picaud, J.-C.; Buffin, R. Human Milk—Treatment and Quality of Banked Human Milk. Clin. Perinatol. 2017, 44, 95–119. [Google Scholar] [CrossRef]
- Escuder-Vieco, D.; Espinosa-Martos, I.; Rodríguez, J.M.; Fernández, L.; Pallás-Alonso, C.R. Effect of HTST and Holder Pasteurization on the Concentration of Immunoglobulins, Growth Factors, and Hormones in Donor Human Milk. Front. Immunol. 2018, 9, 2222. [Google Scholar] [CrossRef] [Green Version]
- Marousez, L.; Tran, L.; Micours, E.; Lamballerie, M.D.; Gottrand, F.; Pierrat, V.; Eberlé, D.; Ley, D.; Lesage, J. Metabolic Hormones in Human Breast Milk Are Preserved by High Hydrostatic Pressure Processing but Reduced by Holder Pasteurization. Food Chem. 2022, 377, 131957. [Google Scholar] [CrossRef] [PubMed]
- Demazeau, G.; Rivalain, N. The Development of High Hydrostatic Pressure Processes as an Alternative to Other Pathogen Reduction Methods. J. Appl. Microbiol. 2011, 110, 1359–1369. [Google Scholar] [CrossRef] [PubMed]
- Billeaud, C. High Hydrostatic Pressure Treatment Ensures the Microbiological Safety of Human Milk Including Bacillus Cereus and Preservation of Bioactive Proteins Including Lipase and Immuno-Proteins: A Narrative Review. Foods 2021, 10, 1327. [Google Scholar] [CrossRef] [PubMed]
- Demazeau, G.; Plumecocq, A.; Lehours, P.; Martin, P.; Couëdelo, L.; Billeaud, C. A New High Hydrostatic Pressure Process to Assure the Microbial Safety of Human Milk While Preserving the Biological Activity of Its Main Components. Front. Public Health 2018, 6, 306. [Google Scholar] [CrossRef]
- Marousez, L.; Sprenger, N.; Lamballerie, M.D.; Jaramillo-Ortiz, S.; Tran, L.; Micours, E.; Gottrand, F.; Howsam, M.; Tessier, F.J.; Ley, D.; et al. High Hydrostatic Pressure Processing of Human Milk Preserves Milk Oligosaccharides and Avoids Formation of Maillard Reaction Products. Clin. Nutr. 2022, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wesolowska, A.; Sinkiewicz-Darol, E.; Barbarska, O.; Strom, K.; Rutkowska, M.; Karzel, K.; Rosiak, E.; Oledzka, G.; Orczyk-Pawiłowicz, M.; Rzoska, S.; et al. New Achievements in High-Pressure Processing to Preserve Human Milk Bioactivity. Front. Pediatr. 2018, 6, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, F.; Neetoo, H.; Chen, H.; Li, J. High Hydrostatic Pressure Processing: A Promising Nonthermal Technology to Inactivate Viruses in High-Risk Foods. Annu. Rev. Food Sci. Technol. 2015, 6, 389–409. [Google Scholar] [CrossRef]
- Han, S.; Hyun, S.; Son, J.W.; Song, M.S.; Lim, D.J.; Choi, C.; Park, S.H.; Ha, S. Innovative Nonthermal Technologies for Inactivation of Emerging Foodborne Viruses. Compr. Rev. Food Sci. Food Saf. 2023. [Google Scholar] [CrossRef]
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Alfenas-Zerbini, P.; Dempsey, D.M.; Dutilh, B.E.; García, M.L.; Hendrickson, R.C.; et al. Recent Changes to Virus Taxonomy Ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2022, 167, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- V’kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus Biology and Replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef]
- Dijkman, R.; Hoek, L. van der Human Coronaviruses 229E and NL63: Close Yet still So Far. J. Formos. Med. Assoc. 2009, 108, 270–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslan, A.T.; Balaban, H.Y. Hepatitis E Virus: Epidemiology, Diagnosis, Clinical Manifestations, and Treatment. World J. Gastroenterol. 2020, 26, 5543–5560. [Google Scholar] [CrossRef] [PubMed]
- Velavan, T.P.; Pallerla, S.R.; Johne, R.; Todt, D.; Steinmann, E.; Schemmerer, M.; Wenzel, J.J.; Hofmann, J.; Shih, J.W.K.; Wedemeyer, H.; et al. Hepatitis E: An Update on One Health and Clinical Medicine. Liver Int. 2021, 41, 1462–1473. [Google Scholar] [CrossRef]
- Khuroo, M.S. Hepatitis E and Pregnancy: An Unholy Alliance Unmasked from Kashmir, India. Viruses 2021, 13, 1329. [Google Scholar] [CrossRef]
- Nimgaonkar, I.; Ding, Q.; Schwartz, R.E.; Ploss, A. Hepatitis E Virus: Advances and Challenges. Nat. Reviews. Gastroenterol. Hepatol. 2018, 15, 96–110. [Google Scholar] [CrossRef]
- Feng, Z.; Hirai-Yuki, A.; McKnight, K.L.; Lemon, S.M. Naked Viruses That Aren’t Always Naked: Quasi-Enveloped Agents of Acute Hepatitis. Annu. Rev. Virol. 2014, 1, 539–560. [Google Scholar] [CrossRef]
- Blight, K.J.; Mckeating, J.A.; Rice, C.M. Highly Permissive Cell Lines for Subgenomic and Genomic Hepatitis C Virus RNA Replication. J. Virol. 2002, 76, 13001–13014. [Google Scholar] [CrossRef] [Green Version]
- Montpellier, C.; Wychowski, C.; Sayed, I.M.; Meunier, J.-C.; Saliou, J.-M.; Ankavay, M.; Bull, A.; Pillez, A.; Abravanel, F.; Helle, F.; et al. Hepatitis E Virus Lifecycle and Identification of 3 Forms of the ORF2 Capsid Protein. Gastroenterology 2018, 154, 211–223.e8. [Google Scholar] [CrossRef]
- Bentaleb, C.; Hervouet, K.; Montpellier, C.; Camuzet, C.; Ferrié, M.; Burlaud-Gaillard, J.; Bressanelli, S.; Metzger, K.; Werkmeister, E.; Ankavay, M.; et al. The Endocytic Recycling Compartment Serves as a Viral Factory for Hepatitis E Virus. Cell. Mol. Life Sci. 2022, 79, 615. [Google Scholar] [CrossRef]
- Yin, X.; Ambardekar, C.; Lu, Y.; Feng, Z. Distinct Entry Mechanisms for Non-Enveloped and Quasi-Enveloped Hepatitis E Virus. J. Virol. 2016, 90, JVI.02804-15. [Google Scholar] [CrossRef] [Green Version]
- Unger, S.; Christie-Holmes, N.; Guvenc, F.; Budylowski, P.; Mubareka, S.; Gray-Owen, S.D.; O’Connor, D.L. Holder Pasteurization of Donated Human Milk Is Effective in Inactivating SARS-CoV-2. Cmaj 2020, 192, cmaj.201309. [Google Scholar] [CrossRef]
- Walker, G.J.; Clifford, V.; Bansal, N.; Stella, A.O.; Turville, S.; Stelzer-Braid, S.; Klein, L.D.; Rawlinson, W. SARS-CoV-2 in Human Milk Is Inactivated by Holder Pasteurisation but Not Cold Storage. J. Paediatr. Child Health 2020, 56, 1872–1874. [Google Scholar] [CrossRef] [PubMed]
- Pitino, M.A.; O’Connor, D.L.; McGeer, A.J.; Unger, S. The Impact of Thermal Pasteurization on Viral Load and Detectable Live Viruses in Human Milk and Other Matrices: A Rapid Review. Appl. Physiol. Nutr. Metab. 2021, 46, 10–26. [Google Scholar] [CrossRef]
- Bhatt, H. Should COVID-19 Mother Breastfeed Her Newborn Child? A Literature Review on the Safety of Breastfeeding for Pregnant Women with COVID-19. Curr. Nutr. Rep. 2021, 10, 71–75. [Google Scholar] [CrossRef]
- Santos-Silva, S.; Gonçalves, H.M.R.; Rivero-Juarez, A.; Van der Poel, W.H.M.; Nascimento, M.S.J.; Mesquita, J.R. Detection of Hepatitis E Virus in Milk: Current Evidence for Viral Excretion in a Wide Range of Mammalian Hosts. Transbound. Emerg. Dis. 2022, 69, 3173–3180. [Google Scholar] [CrossRef]
- Rivero-Juarez, A.; Frias, M.; Rodriguez-Cano, D.; Cuenca-López, F.; Rivero, A. Isolation of Hepatitis E Virus from Breast Milk During Acute Infection. Clin. Infect. Dis. 2016, 62, 1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasheri, N.; Doctor, T.; Chen, A.; Harlow, J.; Gill, A. Evaluation of High-Pressure Processing in Inactivation of the Hepatitis E Virus. Front. Microbiol. 2020, 11, 461. [Google Scholar] [CrossRef] [Green Version]
- Johne, R.; Wolff, A.; Gadicherla, A.K.; Filter, M.; Schlüter, O. Stability of Hepatitis E Virus at High Hydrostatic Pressure Processing. Int. J. Food Microbiol. 2021, 339, 109013. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Li, Y.; Yu, W.; Jing, S.; Wang, J.; Long, F.; He, Z.; Yang, C.; Bi, Y.; Cao, W.; et al. Excretion of Infectious Hepatitis E Virus into Milk in Cows Imposes High Risks of Zoonosis. Hepatology 2016, 64, 350–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouquet, P.; Alexandre, V.; De Lamballerie, M.; Ley, D.; Lesage, J.; Goffard, A.; Cocquerel, L. Effect of High Hydrostatic Pressure Processing and Holder Pasteurization of Human Milk on Inactivation of Human Coronavirus 229E and Hepatitis E Virus. Viruses 2023, 15, 1571. https://doi.org/10.3390/v15071571
Bouquet P, Alexandre V, De Lamballerie M, Ley D, Lesage J, Goffard A, Cocquerel L. Effect of High Hydrostatic Pressure Processing and Holder Pasteurization of Human Milk on Inactivation of Human Coronavirus 229E and Hepatitis E Virus. Viruses. 2023; 15(7):1571. https://doi.org/10.3390/v15071571
Chicago/Turabian StyleBouquet, Peggy, Virginie Alexandre, Marie De Lamballerie, Delphine Ley, Jean Lesage, Anne Goffard, and Laurence Cocquerel. 2023. "Effect of High Hydrostatic Pressure Processing and Holder Pasteurization of Human Milk on Inactivation of Human Coronavirus 229E and Hepatitis E Virus" Viruses 15, no. 7: 1571. https://doi.org/10.3390/v15071571