
SARS-CoV-2 in Domestic UK Cats from Alpha to Omicron: Swab Surveillance and Case Reports

 , ,
, ,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
3. Results
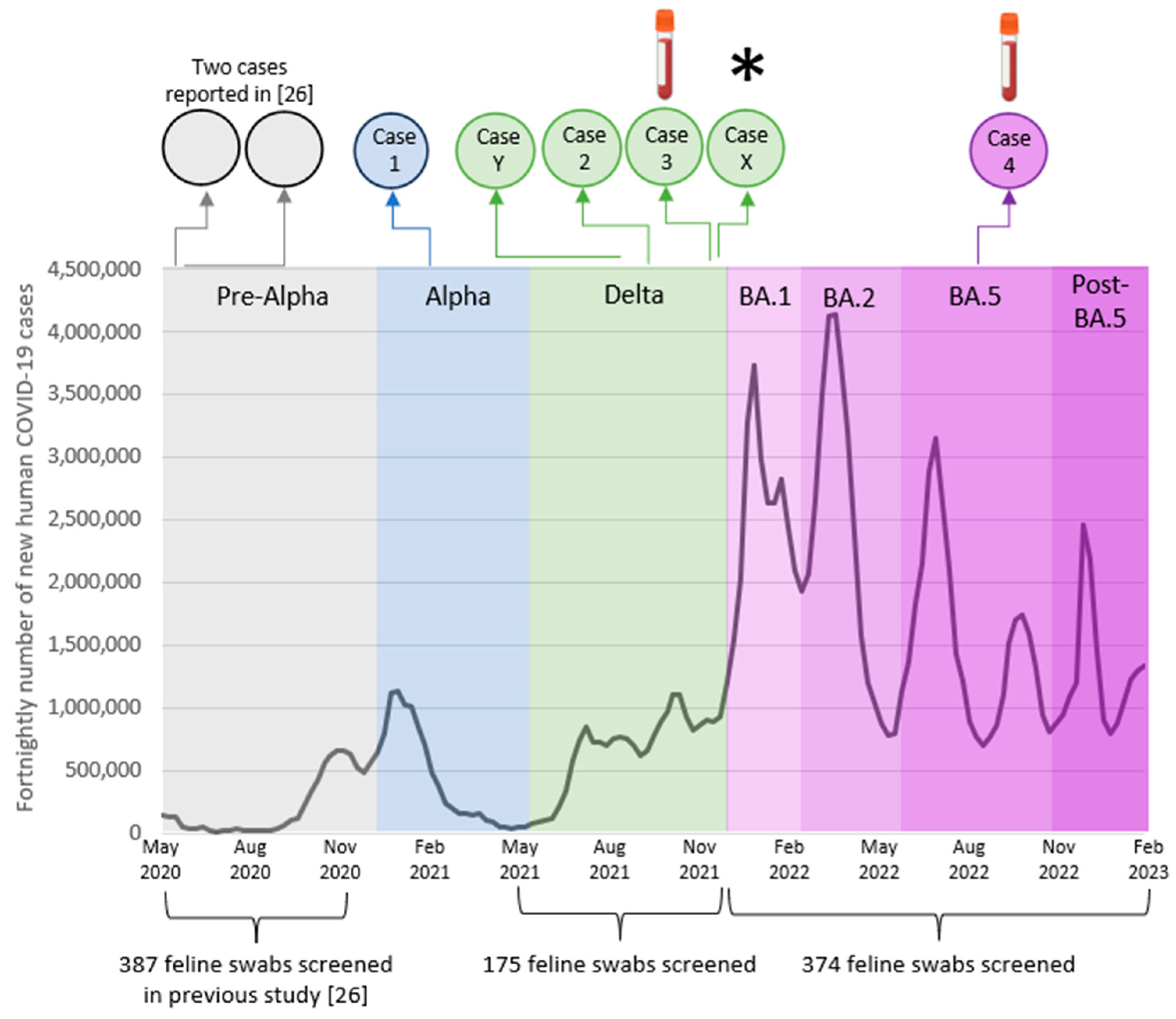
3.1. Active Surveillance
3.2. Passive Surveillance
3.3. Seropositive Cases Identified Previously
3.4. Case Series
3.4.1. Case One: Acute Pyrexia
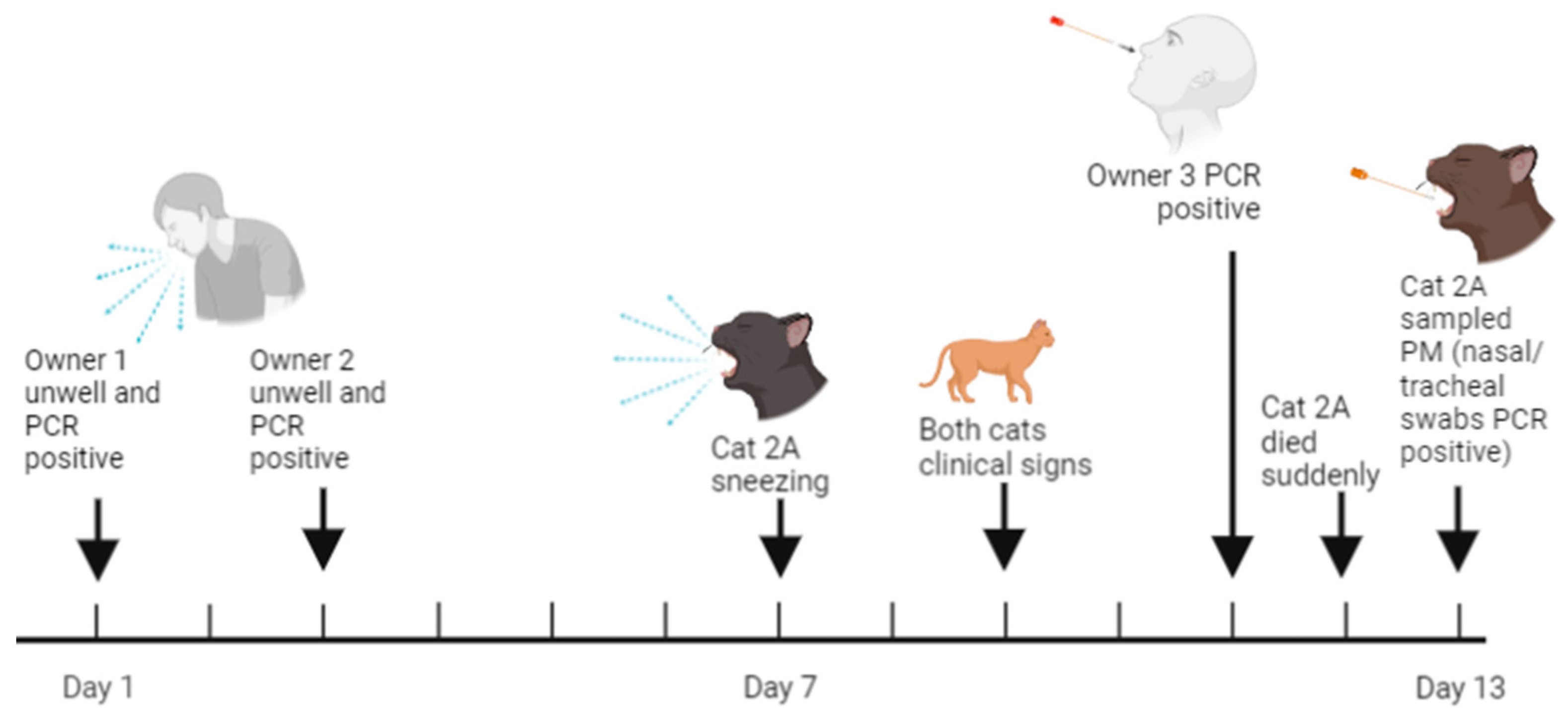
3.4.2. Case Two: Sudden Death
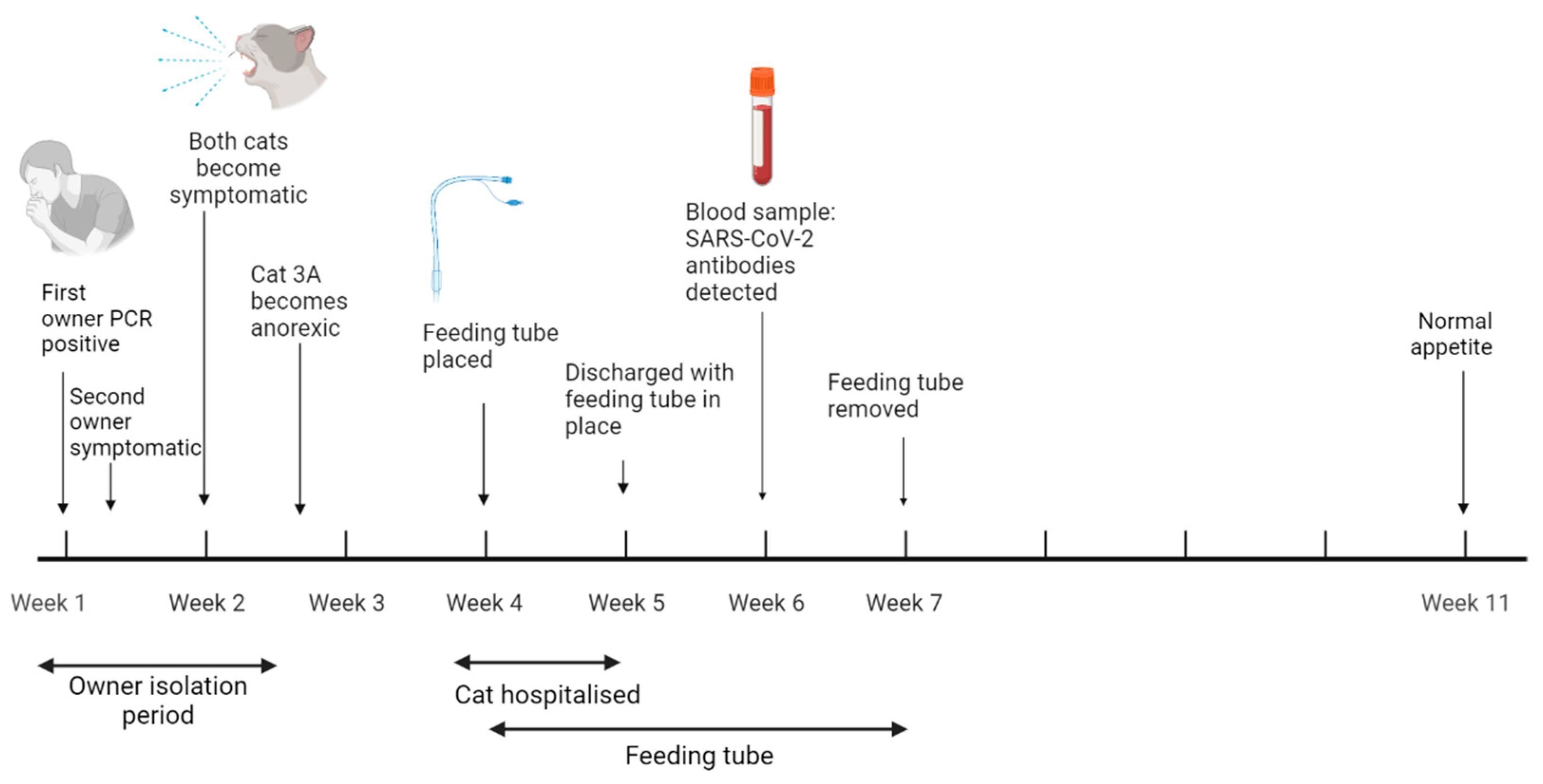
3.4.3. Case Three: Anorexia
3.4.4. Case Four: Respiratory Distress and Renal Disease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Supplementary Methods
Appendix A.1. SARS-CoV-2 RT-qPCR
Appendix A.1.1. Reagents
Appendix A.1.2. Primer/Probe Sequences

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temp (°C) | Time | Cycles |
|---|---|---|
| 55 | 10 min | 1 |
| 95 | 1 min | 1 |
| 95 | 10 s | 45 |
| 58 | 1 min |
Appendix A.2. SARS-CoV-2 Viral Full Genome Sequencing
Appendix A.2.1. Cats 1A, 2A and Y
Appendix A.2.2. Cat X
Appendix A.3. Phylogenetic and Mutation Analysis of Cat Sequences
Appendix B. Supplementary Information
| Swab | N1 Assay Ct | N2 Assay Ct |
|---|---|---|
| Nasal | 18 | 17.1 |
| Oral | Undet. | Undet. |
| Tracheal | 18.9 | 18.1 |
| Rectal | Undet. | 37.5 |



| Cat X (Delta AY.4.2.1) | Cat 2A (Delta AY.4) | Cat 1A (Alpha B.1.1.7) |
|---|---|---|
| T19R | T19R | |
| V36F | ||
| T95I | T95I | |
| I68I | ||
| H69del | ||
| V70del | ||
| G142D | G142D | |
| Y144del | ||
| Y145H | ||
| E156del | E156del | |
| F157del | F157del | |
| R158G | R158G | |
| A222V | ||
| L452R | L452R | |
| T478K | T478K | |
| N501Y | ||
| A570D | ||
| D614G | D614G | D614G |
| P681R | P681R | |
| P681H | ||
| T716I | ||
| L821L | ||
| D950N | D950N | |
| S982A | ||
| D1118H | ||
| L1141F |
References
- Nerpel, A.; Yang, L.; Sorger, J.; Käsbohrer, A.; Walzer, C.; Desvars-Larrive, A. SARS-ANI: A global open access dataset of reported SARS-CoV-2 events in animals. Sci. Data 2022, 9, 438. [Google Scholar] [CrossRef] [PubMed]
- Braun, K.M.; Moreno, G.K.; Halfmann, P.J.; Hodcroft, E.B.; Baker, D.A.; Boehm, E.C.; Weiler, A.M.; Haj, A.K.; Hatta, M.; Chiba, S. Transmission of SARS-CoV-2 in domestic cats imposes a narrow bottleneck. PLoS Pathog. 2021, 17, e1009373. [Google Scholar] [CrossRef] [PubMed]
- Halfmann, P.J.; Hatta, M.; Chiba, S.; Maemura, T.; Fan, S.; Takeda, M.; Kinoshita, N.; Hattori, S.-i.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K. Transmission of SARS-CoV-2 in domestic cats. N. Engl. J. Med. 2020, 383, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Lauth, A.M.; Hartwig, A.E.; Porter, S.M.; Gordy, P.W.; Nehring, M.; Byas, A.D.; VandeWoude, S.; Ragan, I.K.; Maison, R.M.; Bowen, R.A. Experimental infection of domestic dogs and cats with SARS-CoV-2: Pathogenesis, transmission, and response to reexposure in cats. Proc. Natl. Acad. Sci. USA 2020, 117, 26382–26388. [Google Scholar] [CrossRef]
- Sila, T.; Sunghan, J.; Laochareonsuk, W.; Surasombatpattana, S.; Kongkamol, C.; Ingviya, T.; Siripaitoon, P.; Kositpantawong, N.; Kanchanasuwan, S.; Hortiwakul, T. Suspected Cat-to-Human Transmission of SARS-CoV-2, Thailand, July–September 2021. Emerg. Infect. Dis. 2022, 28, 1485. [Google Scholar] [CrossRef]
- Siegrist, A.A.; Richardson, K.L.; Ghai, R.R.; Pope, B.; Yeadon, J.; Culp, B.; Behravesh, C.B.; Liu, L.; Brown, J.A.; Boyer, L.V. Probable Transmission of SARS-CoV-2 from African Lion to Zoo Employees, Indiana, USA, 2021. Emerg. Infect. Dis. 2023, 29, 1102. [Google Scholar] [CrossRef]
- OIE. Statement from the Advisory Group on SARS-CoV-2 Evolution in Animals. 2022. Available online: https://www.oie.int/app/uploads/2022/01/statement-agve-omicron.pdf (accessed on 11 January 2022).
- Kannekens-Jager, M.M.; de Rooij, M.M.; de Groot, Y.; Biesbroeck, E.; de Jong, M.K.; Pijnacker, T.; Smit, L.A.; Schuurman, N.; Broekhuizen-Stins, M.J.; Zhao, S. SARS-CoV-2 infection in dogs and cats is associated with contact to COVID-19-positive household members. Transbound. Emerg. Dis. 2022, 69, 4034–4040. [Google Scholar] [CrossRef]
- Goryoka, G.W.; Cossaboom, C.M.; Gharpure, R.; Dawson, P.; Tansey, C.; Rossow, J.; Mrotz, V.; Rooney, J.; Torchetti, M.; Loiacono, C.M. One Health investigation of SARS-CoV-2 infection and seropositivity among pets in households with confirmed human COVID-19 cases—Utah and Wisconsin, 2020. Viruses 2021, 13, 1813. [Google Scholar] [CrossRef]
- Barrs, V.R.; Peiris, M.; Tam, K.W.; Law, P.Y.; Brackman, C.J.; To, E.M.; Yu, V.Y.; Chu, D.K.; Perera, R.A.; Sit, T.H. SARS-CoV-2 in quarantined domestic cats from COVID-19 households or close contacts, Hong Kong, China. Emerg. Infect. Dis. 2020, 26, 3071. [Google Scholar] [CrossRef]
- Bienzle, D.; Rousseau, J.; Marom, D.; MacNicol, J.; Jacobson, L.; Sparling, S.; Prystajecky, N.; Fraser, E.; Weese, J.S. Risk factors for SARS-CoV-2 infection and illness in cats and dogs. Emerg. Infect. Dis. 2022, 28, 1154. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmeier, E.; Chan, T.; Meli, M.L.; Willi, B.; Wolfensberger, A.; Reitt, K.; Hüttl, J.; Jones, S.; Tyson, G.; Hosie, M.J. A Risk Factor Analysis of SARS-CoV-2 Infection in Animals in COVID-19-Affected Households. Viruses 2023, 15, 731. [Google Scholar] [CrossRef] [PubMed]
- SPI-M-O: Consensus Statement on COVID-19. 2021. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/993321/S1267_SPI-M-O_Consensus_Statement.pdf (accessed on 31 January 2022).
- Adler, J.M.; Weber, C.; Wernike, K.; Michelitsch, A.; Friedrich, K.; Trimpert, J.; Beer, M.; Kohn, B.; Osterrieder, K.; Müller, E. Prevalence of anti-severe acute respiratory syndrome coronavirus 2 antibodies in cats in Germany and other European countries in the early phase of the coronavirus disease-19 pandemic. Zoonoses Public Health 2022, 69, 439–450. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira-Filho, E.F.; de Carvalho, O.V.; Carneiro, I.O.; Fernandes, F.; Vaz, S.N.; Pedroso, C.; Gonzalez-Auza, L.; Urbieta, V.C.; Kühne, A.; Mayoral, R. Frequent infection of cats with SARS-CoV-2 irrespective of pre-existing enzootic coronavirus immunity, Brazil 2020. Front. Immunol. 2022, 13, 1000. [Google Scholar] [CrossRef]
- Bessière, P.; Vergne, T.; Battini, M.; Brun, J.; Averso, J.; Joly, E.; Guérin, J.-L.; Cadiergues, M.-C. SARS-CoV-2 Infection in Companion Animals: Prospective Serological Survey and Risk Factor Analysis in France. Viruses 2022, 14, 1178. [Google Scholar] [CrossRef]
- Smith, S.L.; Anderson, E.R.; Cansado-Utrilla, C.; Prince, T.; Farrell, S.; Brant, B.; Smyth, S.; Noble, P.M.; Pinchbeck, G.L.; Marshall, N.; et al. SARS-CoV-2 neutralising antibodies in dogs and cats in the United Kingdom. Curr. Res. Virol. Sci. 2021, 2, 100011. [Google Scholar] [CrossRef]
- Tyson, G.; Jones, S.; Logan, N.; McDonald, M.; Marshall, L.; Murcia, P.; Willett, B.; Weir, W.; Hosie, M. SARS-CoV-2 Seroprevalence and Cross-Variant Antibody Neutralization in Cats, United Kingdom. Emerg. Infect. Dis. 2023, 29, 1223. [Google Scholar] [CrossRef]
- Gaudreault, N.N.; Trujillo, J.D.; Carossino, M.; Meekins, D.A.; Morozov, I.; Madden, D.W.; Indran, S.V.; Bold, D.; Balaraman, V.; Kwon, T. SARS-CoV-2 infection, disease and transmission in domestic cats. Emerg. Microbes Infect. 2020, 9, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Barroso-Arévalo, S.; Sánchez-Morales, L.; Pérez-Sancho, M.; Domínguez, L.; Sánchez-Vizcaíno, J.M. First detection of SARS-CoV-2 B.1.617.2 (Delta) variant of concern in a symptomatic cat in Spain. Front. Vet. Sci. 2022, 9, 841430. [Google Scholar] [CrossRef]
- Neira, V.; Brito, B.; Agüero, B.; Berrios, F.; Valdés, V.; Gutierrez, A.; Ariyama, N.; Espinoza, P.; Retamal, P.; Holmes, E.C. A household case evidences shorter shedding of SARS-CoV-2 in naturally infected cats compared to their human owners. Emerg. Microbes Infect. 2021, 10, 376–383. [Google Scholar] [CrossRef]
- Liew, A.Y.; Carpenter, A.; Moore, T.A.; Wallace, R.M.; Hamer, S.A.; Hamer, G.L.; Fischer, R.S.; Zecca, I.B.; Davila, E.; Auckland, L.D. Clinical and epidemiologic features of SARS-CoV-2 in dogs and cats compiled through national surveillance in the United States. J. Am. Vet. Med. Assoc. 2023, 261, 480–498. [Google Scholar] [CrossRef] [PubMed]
- Kuhlmeier, E.; Chan, T.; Agüí, C.V.; Willi, B.; Wolfensberger, A.; Beisel, C.; Topolsky, I.; Beerenwinkel, N.; Stadler, T.; Jones, S. Detection and Molecular Characterization of the SARS-CoV-2 Delta Variant and the Specific Immune Response in Companion Animals in Switzerland. Viruses 2023, 15, 245. [Google Scholar] [CrossRef] [PubMed]
- Garigliany, M.; Van Laere, A.S.; Clercx, C.; Giet, D.; Escriou, N.; Huon, C.; van der Werf, S.; Eloit, M.; Desmecht, D. SARS-CoV-2 Natural Transmission from Human to Cat, Belgium, March 2020. Emerg. Infect. Dis. 2020, 26, 3069–3071. [Google Scholar] [CrossRef]
- Hosie, M.J.; Epifano, I.; Herder, V.; Orton, R.J.; Stevenson, A.; Johnson, N.; MacDonald, E.; Dunbar, D.; McDonald, M.; Howie, F.; et al. Detection of SARS-CoV-2 in respiratory samples from cats in the UK associated with human-to-cat transmission. Vet. Rec. 2021, 188, e247. [Google Scholar] [CrossRef] [PubMed]
- Klaus, J.; Meli, M.L.; Willi, B.; Nadeau, S.; Beisel, C.; Stadler, T.; Team, E.S.-C.-S.; Egberink, H.; Zhao, S.; Lutz, H. Detection and genome sequencing of SARS-CoV-2 in a domestic cat with respiratory signs in Switzerland. Viruses 2021, 13, 496. [Google Scholar] [CrossRef]
- Keller, M.; Hagag, I.T.; Balzer, J.; Beyer, K.; Kersebohm, J.C.; Sadeghi, B.; Wernike, K.; Höper, D.; Wylezich, C.; Beer, M. Detection of SARS-CoV-2 variant B.1.1.7 in a cat in Germany. Res. Vet. Sci. 2021, 140, 229–232. [Google Scholar] [CrossRef]
- Zoccola, R.; Beltramo, C.; Magris, G.; Peletto, S.; Acutis, P.; Bozzetta, E.; Radovic, S.; Zappulla, F.; Porzio, A.M.; Gennero, M.S. First detection of an Italian human-to-cat outbreak of SARS-CoV-2 Alpha variant–lineage B.1.1.7. One Health 2021, 13, 100295. [Google Scholar] [CrossRef]
- Ferasin, L.; Fritz, M.; Ferasin, H.; Becquart, P.; Corbet, S.; Ar Gouilh, M.; Legros, V.; Leroy, E.M. Infection with SARS-CoV-2 variant B.1.1.7 detected in a group of dogs and cats with suspected myocarditis. Vet. Rec. 2021, 189, e944. [Google Scholar] [CrossRef]
- Carvallo, F.R.; Martins, M.; Joshi, L.R.; Caserta, L.C.; Mitchell, P.K.; Cecere, T.; Hancock, S.; Goodrich, E.L.; Murphy, J.; Diel, D.G. Severe SARS-CoV-2 Infection in a Cat with Hypertrophic Cardiomyopathy. Viruses 2021, 13, 1510. [Google Scholar] [CrossRef]
- Chetboul, V.; Foulex, P.; Kartout, K.; Klein, A.M.; Sailleau, C.; Dumarest, M.; Delaplace, M.; Gouilh, M.A.; Mortier, J.; Le Poder, S. Myocarditis and Subclinical-Like Infection Associated With SARS-CoV-2 in Two Cats Living in the Same Household in France: A Case Report With Literature Review. Front. Vet. Sci. 2021, 8, 748869. [Google Scholar] [CrossRef]
- McAloose, D.; Laverack, M.; Wang, L.; Killian, M.L.; Caserta, L.C.; Yuan, F.; Mitchell, P.K.; Queen, K.; Mauldin, M.R.; Cronk, B.D. From people to Panthera: Natural SARS-CoV-2 infection in tigers and lions at the Bronx Zoo. MBio 2020, 11, e02220. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, S.L.; Diel, D.G.; Wang, L.; Zec, S.; Laverack, M.; Martins, M.; Caserta, L.C.; Killian, M.L.; Terio, K.; Olmstead, C. SARS-CoV-2 infection and longitudinal fecal screening in Malayan tigers (Panthera tigris jacksoni), Amur tigers (Panthera tigris altaica), and African lions (Panthera leo krugeri) at the Bronx Zoo, New York, USA. J. Zoo Wildl. Med. 2021, 51, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Kumar, N.; Bhatia, S.; Aasdev, A.; Kanniappan, S.; Sekhar, A.T.; Gopinadhan, A.; Silambarasan, R.; Sreekumar, C.; Dubey, C.K. SARS-CoV-2 Delta variant among asiatic lions, India. Emerg. Infect. Dis. 2021, 27, 2723. [Google Scholar] [CrossRef] [PubMed]
- Giraldo-Ramirez, S.; Rendon-Marin, S.; Jaimes, J.A.; Martinez-Gutierrez, M.; Ruiz-Saenz, J. SARS-CoV-2 clinical outcome in domestic and wild cats: A systematic review. Animals 2021, 11, 2056. [Google Scholar] [CrossRef] [PubMed]
- Klaus, J.; Zini, E.; Hartmann, K.; Egberink, H.; Kipar, A.; Bergmann, M.; Palizzotto, C.; Zhao, S.; Rossi, F.; Franco, V. SARS-CoV-2 infection in dogs and cats from southern Germany and northern Italy during the first wave of the COVID-19 pandemic. Viruses 2021, 13, 1453. [Google Scholar] [CrossRef] [PubMed]
- Murcia, P.; Streiker, D.; Philipe, A.D.S.; Robertson, D.; Jarrett, R.; Willett, B.; Hosie, M.; Biek, R.; Allan, K.; Weir, W. Send cat and dog samples to test for SARS-CoV-2. Vet. Rec. 2020, 186, 571. [Google Scholar] [CrossRef]
- Boom, R.; Sol, C.; Salimans, M.; Jansen, C.; Wertheim-van Dillen, P.; Van der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Logan, N.; Tyson, G.; Orton, R.; Harvey, W.T.; Perkins, J.S.; Mollett, G.; Blacow, R.M.; Consortium, C.-G.U.; Peacock, T.P. Reduced neutralisation of the Delta (B.1.617.2) SARS-CoV-2 variant of concern following vaccination. PLoS Pathog. 2021, 17, e1010022. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; De Lorenzo, G.; Furnon, W.; Cantoni, D.; Scott, S.; Logan, N.; Ashraf, S. SARS-CoV-2 Omicron is an immune escape variant with an altered cell entry pathway. Nat. Microbiol. 2022, 7, 1161–1179. [Google Scholar] [CrossRef]
- APHA. SARS-CoV-2 in Animals—Case Definition, Testing and International Reporting Obligations. 2021. Available online: http://apha.defra.gov.uk/documents/ov/Briefing-Note-0921.pdf (accessed on 1 May 2021).
- Office for National Statistics (ONS). Coronavirus (COVID19) Infection Survey technical Article: Cumulative Incidence of the Number of People Who have been Infected with COVID-19 by Variant and Age, England: 9 February 2023. 2023. Available online: https://www.ons.gov.uk/peoplepopulationandcommunity/healthandsocialcare/conditionsanddiseases/articles/coronaviruscovid19infectionsurveytechnicalarticlecumulativeincidenceofthenumberofpeoplewhohavebeeninfectedwithcovid19byvariantandageengland/9february2023 (accessed on 1 March 2023).
- O’Toole, Á.; Scher, E.; Underwood, A.; Jackson, B.; Hill, V.; McCrone, J.T.; Colquhoun, R.; Ruis, C.; Abu-Dahab, K.; Taylor, B.; et al. Assignment of epidemiological lineages in an emerging pandemic using the pangolin tool. Virus Evol. 2021, 7, veab064. [Google Scholar] [CrossRef]
- Khare, S.; Gurry, C.; Freitas, L.; Schultz, M.B.; Bach, G.; Diallo, A.; Akite, N.; Ho, J.; Lee, R.T.; Yeo, W. GISAID’s role in pandemic response. China CDC Wkly. 2021, 3, 1049. [Google Scholar] [CrossRef] [PubMed]
- International Renal Interest Society. IRIS Staging of CKD (Modified in 2023). 2023. Available online: http://www.iris-kidney.com/guidelines/staging.html (accessed on 14 June 2023).
- Martins, M.; do Nascimento, G.M.; Nooruzzaman, M.; Yuan, F.; Chen, C.; Caserta, L.C.; Miller, A.D.; Whittaker, G.R.; Fang, Y.; Diel, D.G. The Omicron variant BA.1.1 presents a lower pathogenicity than B.1 D614G and Delta variants in a feline model of SARS-CoV-2 infection. J. Virol. 2022, 96, e00961-22. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Morales, L.; Sánchez-Vizcaíno, J.M.; Pérez-Sancho, M.; Domínguez, L.; Barroso-Arévalo, S. The Omicron (B.1.1.529) SARS-CoV-2 variant of concern also affects companion animals. Front. Vet. Sci. 2022, 9, 940710. [Google Scholar] [CrossRef]
- Pickering, B.; Lung, O.; Maguire, F.; Kruczkiewicz, P.; Kotwa, J.D.; Buchanan, T.; Gagnier, M.; Guthrie, J.L.; Jardine, C.M.; Marchand-Austin, A. Divergent SARS-CoV-2 variant emerges in white-tailed deer with deer-to-human transmission. Nat. Microbiol. 2022, 7, 2011–2024. [Google Scholar] [CrossRef]
- Munnink, B.B.O.; Sikkema, R.S.; Nieuwenhuijse, D.F.; Molenaar, R.J.; Munger, E.; Molenkamp, R.; Van Der Spek, A.; Tolsma, P.; Rietveld, A.; Brouwer, M. Transmission of SARS-CoV-2 on mink farms between humans and mink and back to humans. Science 2021, 371, 172–177. [Google Scholar] [CrossRef]
- Michel, K. Management of anorexia in the cat. J. Feline Med. Surg. 2001, 3, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Burges Watson, D.L.; Campbell, M.; Hopkins, C.; Smith, B.; Kelly, C.; Deary, V. Altered smell and taste: Anosmia, parosmia and the impact of long COVID-19. PLoS ONE 2021, 16, e0256998. [Google Scholar] [CrossRef]
- Hecht, G.; Sarbo, N.; Svoboda, W.; Mead, H.L.; Ruberto, I.; Altin, J.A.; Engelthaler, D.M.; Venkat, H.; Yaglom, H.D. “Sniffing” out SARS-CoV-2 in Arizona working dogs: An exploratory serosurvey. Front. Vet. Sci. 2023, 10, 1166101. [Google Scholar] [CrossRef]
- Segalés, J.; Puig, M.; Rodon, J.; Avila-Nieto, C.; Carrillo, J.; Cantero, G.; Terrón, M.T.; Cruz, S.; Parera, M.; Noguera-Julián, M. Detection of SARS-CoV-2 in a cat owned by a COVID-19−affected patient in Spain. Proc. Natl. Acad. Sci. USA 2020, 117, 24790–24793. [Google Scholar] [CrossRef]
- Shirazi, S.; Mami, S.; Mohtadi, N.; Ghaysouri, A.; Tavan, H.; Nazari, A.; Kokhazadeh, T.; Mollazadeh, R. Sudden cardiac death in COVID-19 patients, a report of three cases. Future Cardiol. 2020, 17, 113–118. [Google Scholar] [CrossRef]
- Al-Aly, Z.; Xie, Y.; Bowe, B. High-dimensional characterization of post-acute sequelae of COVID-19. Nature 2021, 594, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Grubaugh, N.D.; Gangavarapu, K.; Quick, J.; Matteson, N.L.; De Jesus, J.G.; Main, B.J.; Tan, A.L.; Paul, L.M.; Brackney, D.E.; Grewal, S. An amplicon-based sequencing framework for accurately measuring intrahost virus diversity using PrimalSeq and iVar. Genome Biol. 2019, 20, 8. [Google Scholar] [CrossRef] [PubMed]
- Turakhia, Y.; Thornlow, B.; Hinrichs, A.S.; De Maio, N.; Gozashti, L.; Lanfear, R.; Haussler, D.; Corbett-Detig, R. Ultrafast Sample placement on Existing tRees (UShER) enables real-time phylogenetics for the SARS-CoV-2 pandemic. Nat. Genet. 2021, 53, 809–816. [Google Scholar] [CrossRef]



| Animal ID | Household Case | Sampling Date | Clinical Signs | Surveillance Method | RT-qPCR | Serology (PVNT) | Viral Sequence | Highest titre | GISAID Accession Number |
|---|---|---|---|---|---|---|---|---|---|
| Cat 1A | 1 | Feb-21 | Pyrexia | Passive | + | + | Alpha | Alpha | EPI_ISL_17971924 |
| Cat 1B | 1 | May-21 | None, same household as Cat 1A | Passive | Not done | + | Alpha | ||
| Cat Y | Sep-21 | Mild respiratory | Passive | + | Not done | Incomplete | |||
| Cat 2A | 2 | Oct-21 | Mild respiratory then sudden death | Passive | + | Not done | Delta AY.4 | EPI_ISL_17971926 | |
| Cat 2B | 2 | Nov-21 | Mild respiratory, lip licking, same household as Cat 2A | Passive | Not done | + | Delta | ||
| Cat X | Dec-21 | Mild respiratory | Active | + | Not done | Delta AY.4.2.1 | EPI_ISL_17971927 | ||
| Cat 3A | 3 | Dec-21 | Mild respiratory, prolonged hyporexia | Passive | Not done | + | Delta | ||
| Cat 4A | 4 | Sep-22 | Severe respiratory | Passive | - | + | Omicron |
| Drug | Trade Name | Route of Administration | Dose | Dosing Interval | Days of Hospitalisation |
|---|---|---|---|---|---|
| Maropitant | Vetemex (Virbac, Bury St Edmunds, UK) | Intravenous/subcutaneous | 1 mg/kg | 24h | 1 to 5 |
| Metoclopramide | Vomend (Dechra, Northwich, UK) | Subcutaneous | 0.5 mg/kg | 12h | 5 to 7 and post-discharge |
| Omeprazole | Intravenous | 1 mg/kg | 12h | 1 and 2 | |
| Mirtazapine | Oral | 1.9–3.8 mg | 48h | Throughout | |
| Vitamin B | Subcutaneous | 0.02 mg/kg | Once | 2 | |
| Buprenorphine | Intravenous/subcutaneous/oral | 0.01–0.02 mg/kg | 6–8 h | 2, 3, 5, 6 and post-discharge | |
| Meloxicam | Loxicam (Norbrook, Newry, UK) | Subcutaneous | 0.1 mg/kg | Once | 5 |
| Amoxycillin-clavulanate | Synulox (Zoetis, Leatherhead, UK) | Oral | 100 mg | 12h | Post-discharge (for 5 days) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, S.; Tyson, G.B.; Orton, R.J.; Smollett, K.; Manna, F.; Kwok, K.; Suárez, N.M.; Logan, N.; McDonald, M.; Bowie, A.; et al. SARS-CoV-2 in Domestic UK Cats from Alpha to Omicron: Swab Surveillance and Case Reports. Viruses 2023, 15, 1769. https://doi.org/10.3390/v15081769
Jones S, Tyson GB, Orton RJ, Smollett K, Manna F, Kwok K, Suárez NM, Logan N, McDonald M, Bowie A, et al. SARS-CoV-2 in Domestic UK Cats from Alpha to Omicron: Swab Surveillance and Case Reports. Viruses. 2023; 15(8):1769. https://doi.org/10.3390/v15081769
Chicago/Turabian StyleJones, Sarah, Grace B. Tyson, Richard J. Orton, Katherine Smollett, Federica Manna, Kirsty Kwok, Nicolás M. Suárez, Nicola Logan, Michael McDonald, Andrea Bowie, and et al. 2023. "SARS-CoV-2 in Domestic UK Cats from Alpha to Omicron: Swab Surveillance and Case Reports" Viruses 15, no. 8: 1769. https://doi.org/10.3390/v15081769
APA StyleJones, S., Tyson, G. B., Orton, R. J., Smollett, K., Manna, F., Kwok, K., Suárez, N. M., Logan, N., McDonald, M., Bowie, A., Filipe, A. D. S., Willett, B. J., Weir, W., & Hosie, M. J. (2023). SARS-CoV-2 in Domestic UK Cats from Alpha to Omicron: Swab Surveillance and Case Reports. Viruses, 15(8), 1769. https://doi.org/10.3390/v15081769