Abstract
Cacao swollen shoot disease (CSSD) caused by complexes of cacao swollen shoot badnaviruses (family Caulimoviridae, genus Badnavirus) remains highly prevalent and devastating in West Africa. The disease continues to impact substantially on cacao yield loss, cacao tree mortality, and decline in foreign exchange income from cacao bean sales. Currently, the disease is estimated to have a prevalence rate of over 30% in Ghana, as assessed in the ongoing third country-wide surveillance program. Although achievements from past research interventions have greatly elucidated the etiology, biology, epidemiology, diagnostics, and management of the disease, there are some outstanding knowledge gaps. The role of these information gaps and their effect on CSSD epidemiology and prevalence remain unanswered. This paper summarizes existing scientific knowledge from past research achievements that have provided elucidation on CSSD epidemiology, management options, and guided future research. The discussion highlights the need for multidisciplinary research with modern tools and institutional collaborators to holistically bring clarity on knowledge gaps on pathogen biology, virus–host-–vector interactions, role of environmental and soil nutrient effects on CSSD severity, evolution pattern, role of alternative hosts on virus species diversity, vector population dynamics, and their overall impact on CSSD prevalence and integrated management in cacao plantations.
1. Introduction
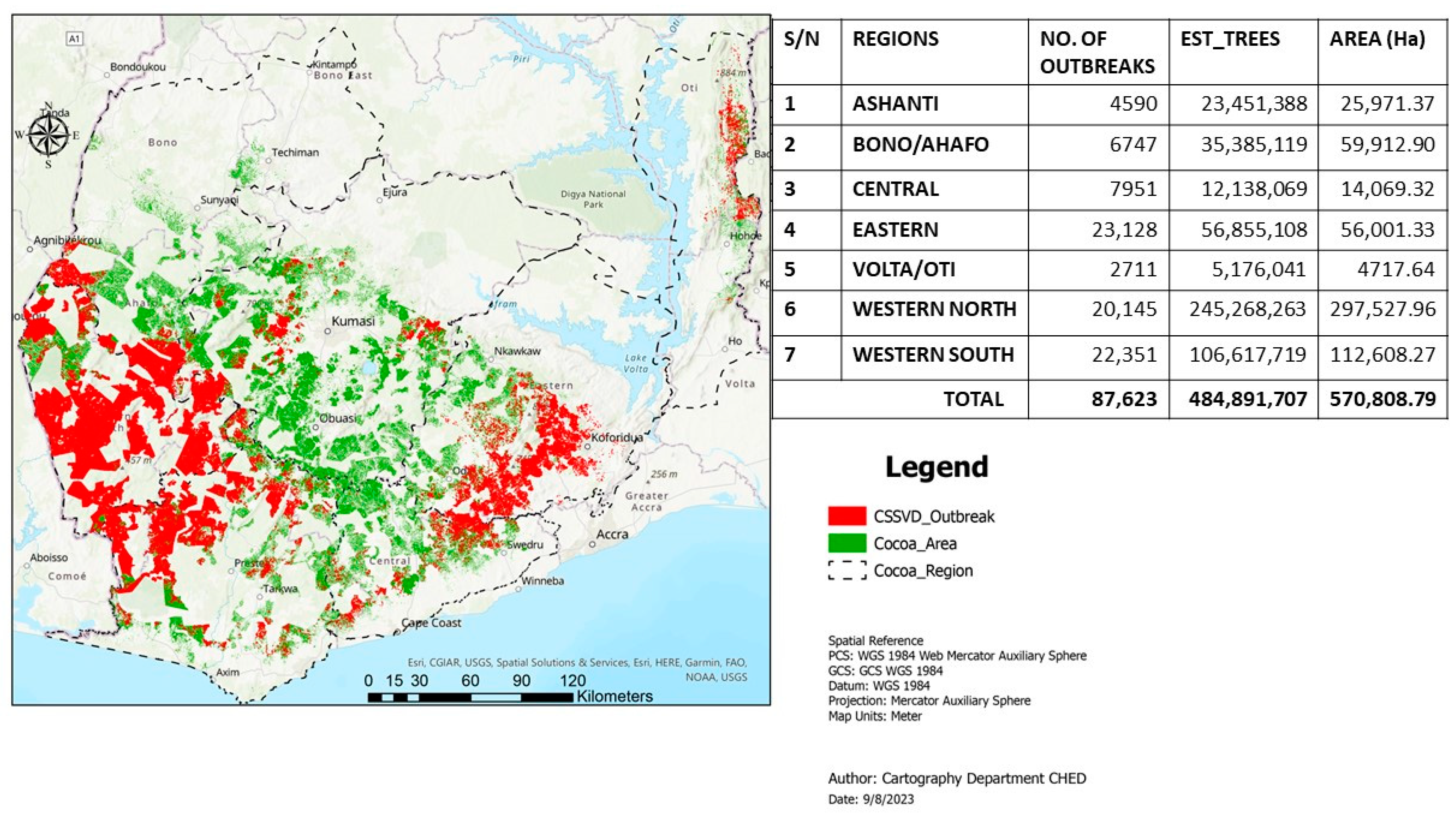
Cacao swollen shoot disease (CSSD) remains a major cacao (Theobroma cacao, L.) production constraint in Ghana and Côte d’Ivoire and the other West African cacao-producing nations, including Togo and Nigeria. The disease, which is transmitted by several species of mealybug insects, could potentially reduce yield by approximately 30% to 50% and even cause death of cacao trees within 2 to 3 years of infection [1]. Although the virus was first identified in the eastern region of Ghana in 1936 [2], it is now widespread in all the major cacao-growing nations in West Africa. The disease continues to contribute to severe cacao tree mortality, most especially the western north and south regions where the bulk of Ghana’s cacao beans are produced [3]. Over the years, the disease has been managed through the rogueing technique, i.e., “cutting-out approach”, with the aim of removing scatter infections in the field [4]. This strategy aims at minimizing inoculum sources and disease pressure to ultimately decrease its spread to newly replanted cacao farms [4,5]. This control strategy has, however, persistently encountered numerous challenges emanating from farmer opposition, limited funding, and frequent discontinuities in the implementation of the program [5]. Cumulatively, these challenges have resulted in high CSSD prevalence and continuous spread to newly established cacao farms in Ghana, especially in the western regions, as documented by the Cocoa Health and Extension Division (CHED) of Ghana Cocoa Board in the ongoing third country-wide survey (CSSD survey map, CHED, 2023) (Figure 1).
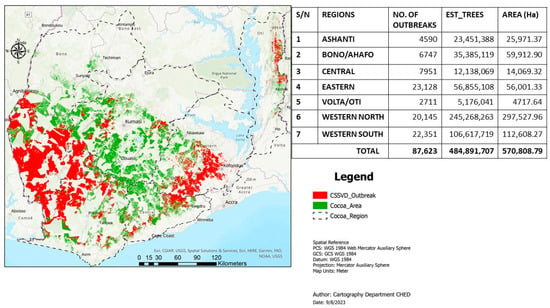
Figure 1.
Current CSSD survey map in Ghana as documented by the Cocoa Health and Extension Division (CHED) of Ghana Cocoa Board in the ongoing third country-wide surveillance program. Courtesy, Cartography Unit of CHED, Survey Report, 2023.
Findings from past research interventions have significantly elucidated the etiology, epidemiology, diagnostics, and directed scientific research and recommendations on strategies for integrated management of the disease in West Africa. However, there are outstanding knowledge gaps on the contemporary biological processes and identity of the viral causal pathogen, symptomatology, and influence of environmental conditions on symptom severity. Also, issues of host–pathogen–vector dynamics, virus species diversity, geographical distribution, and their interactive impact on cacao production have not been adequately clarified from past research. These existing information gaps hamper the deployment of effective CSSD management strategies aimed at minimizing the effect of the virus on cacao production in Ghana and the other producing nations in West Africa. This paper thus provides a review of existing research knowledge that has over the years clarified the status of CSSD etiology, epidemiology, serological, and molecular diagnostics of the disease. Prominent knowledge gaps and their effect on CSSD management are however highlighted and discussed. The paper emphasizes the need for further scientific investigations on the prevailing information gaps for better clarity on their impact on CSSD prevalence, symptom severity, and integrated management in the field.
2. CSSD Etiology and Physicochemical Features
The earliest scientific breakthrough on CSSD research at the Cocoa Research Institute of Ghana (CRIG) was the identification and confirmation of the then putative “swollen shoot” symptoms reported in farmers’ farms to be caused by a viral pathogen [2]. The causal pathogen was subsequently named the “cacao swollen shoot virus” (CSSV) in 1940 [6]. The physicochemical properties of the virus as determined from the earliest purified preparations indicated that the virus particles are organized in a non-enveloped bacilliform fashion, with sizes ranging between 121 × 28 nm and 128 × 28 nm. Other properties of the virus comprised a sedimentation coefficient of 218 S20w; thermal inactivation point (TIP) between 55–60 °C; and longevity in vitro (LIV) of 28–85 days (at 2 °C) [7,8]. The virus was reported to be inactive in causing infection at 50 °C after 10 min of continuous exposure to this temperature [8]. Infectivity of virus preparations was sustained after 24 h storage and was retained after 96 h at 0–4 °C, but was lost at 25 °C and at 1/100 dilution. Virus infective material (viral particles) was also precipitated by half saturation with ammonium sulphate at 25 °C and determined to be stable between pH 6.0 and 8.0 [7].
Further improvements in efficiency associated with CSSV purification protocols enabled the development of the first antisera for the virus for utilization in enzyme-linked immunosorbent assay (ELISA), immunosorbent electron microscopy (ISEM), and virobacterial agglutination tests (VBA) tests as protein-based diagnostic techniques for the virus [9,10]. Availability of purified viral particles led to the commencement of mechanical inoculation techniques for laboratory transmission of the virus for purposes of assaying for evidence of infections with suspected samples [8,9,10]. The efficiency of the mechanical transmission system was subsequently enhanced, and it was used as a tool to screen cacao germplasm materials in resistance breeding activities by the CRIG [11]. Based on these detection techniques, different types of the virus (mainly called strains/isolates) are recognized and characterized to exist in the different cacao growing regions of Ghana and the other growing nations in West Africa [9]. Accordingly, names of CSSV strains/isolates, as kept in the ‘CSSV museum’ of the Cocoa Research Institute of Ghana (CRIG), are labeled based on geographic location of first collection and symptom severity [8,10].
3. Genome Organization of CSSV
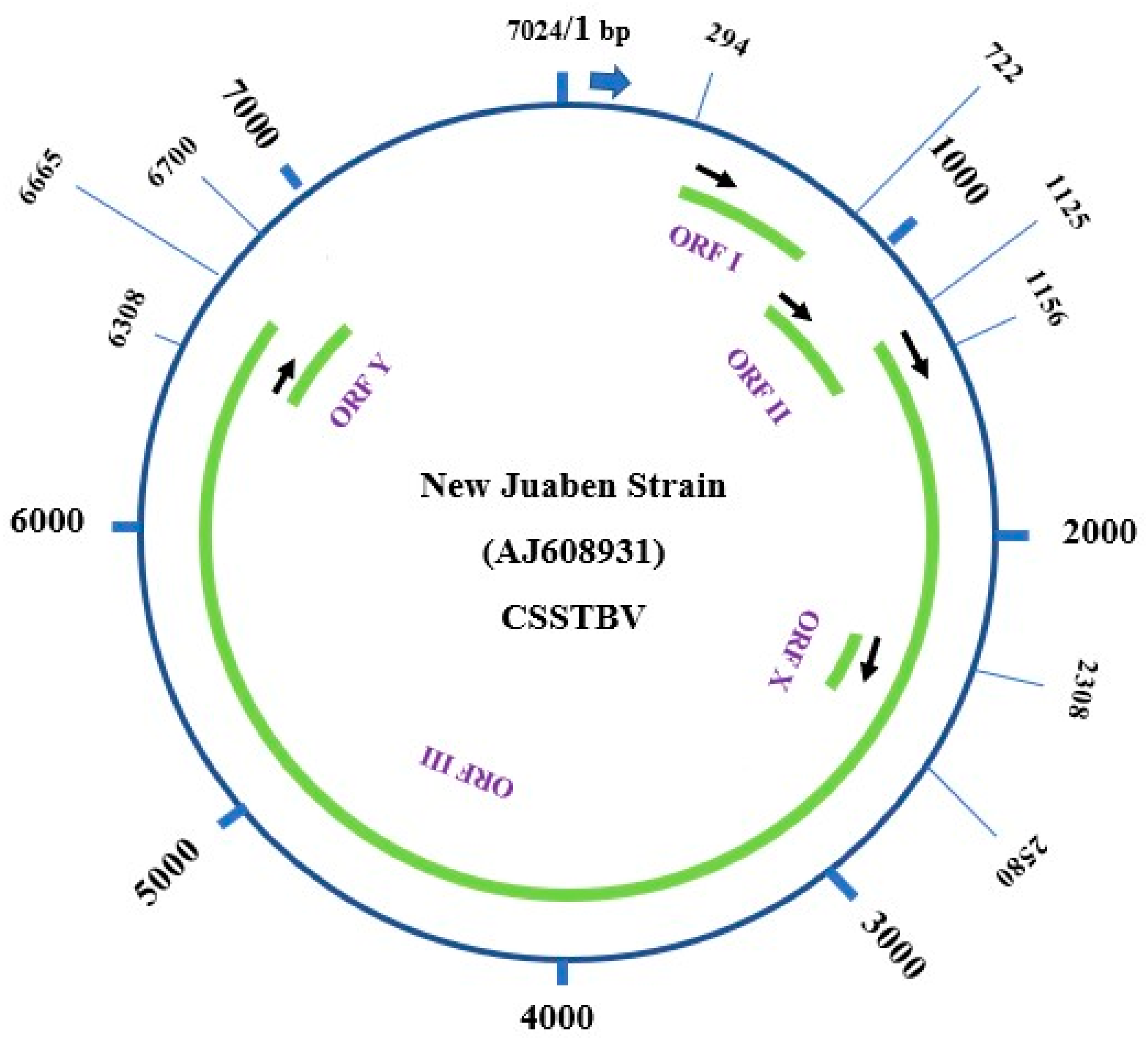
Earlier molecular investigations confirmed CSSV to be a double-stranded DNA virus, and it was taxonomically classified to be a member of plant virus family Caulimoviridae and genus Badnavirus, with double-stranded circular DNA (dsDNA) [12,13,14]. The complete genome sequence of the various strains and isolates of the virus, including the most characterized New Juaben Strain from Ghana, indicated that the genome has been organized into five main putative open reading frames (ORFs 1, 2, 3, X, and Y) on the plus strand of the genome [13,14,15] (Figure 2). Individually, the ORFs code for specific proteins with or without recognized functions [14,15]. For instance, ORF1 encodes a 16.7 kDa protein, whose function has not been clarified. ORF2 encodes a 14.4 kDa nucleic acid- binding protein [15]. ORF3 also codes for a 211 kDa polyprotein which contained several products such as an RNA binding domain, consensus sequences for cell-to-cell movement proteins, a reverse transcriptase (RTase), an aspartyl proteinase, and an RNase H [13,14,15]. The latter two ORFs, X (13 kDa) and Y (14 kDa), were, respectively, noted to overlay ORF3 and encode proteins of unfamiliar functions [12,13,14,16]. The genome size of the various isolates/strains of the virus from field infections generally varies from 7024 bp for isolate N1A to 7242 bp, with the ORF1 being the most conserved coding region compared to the other coding regions [13,14]. Notably, the first region of ORF3, which encodes for movement proteins, is highly conserved, comparable to other para-retroviruses, while the other regions of ORF3 had an intermediate level of variability [13]. Availability of information about the features of the different genomic regions of the virus and their variability provided the foundation for the development of specific and generic polymerase chain reaction (PCR) primers for the detection of the virus [13]. Nonetheless, further updates on genomic information about the more conserved regions of the ORFs of the virus are urgently required. This is based on the availability of more than 80 full-length genomic sequences currently deposited in the GenBank from over eight different species of the virus, in contrast to the few full-length sequences previously considered in the literature on this subject [13].

Figure 2.
A schematic representation of the full-length genomic organization of the New Juabeng strain of the virus depicting positions of the various ORFs, i.e., 1, 2, 3, X, and Y, and the estimated size of the whole genome. This was generated with the latest New Juabeng sequences as deposited in the GenBank with Accession no. AJ608931.
5. CSSD Symptomatology and Mixed Infections in the Field
Different kinds of visible symptoms are expressed in CSSD-affected cacao plants and this is mostly dependent on several factors, including virus species and strains involved in infection, host genotype, and environmental features [23,24]. Contingent on these factors, the types of visible symptoms expressed in affected cacao plants vary in severity and might include the characteristic stem swellings, root swellings, and pod deformation, with or without leaf symptoms [24]. The commonest leaf symptoms include red vein banding of the immature “flush” leaves, followed by chlorotic vein flecking or banding which may occur in angular flecks [24]. Stem swellings could occur at the nodes, internodes, or tips of shoots, and some severe strains sometimes cause infected pods to change shape and become rounder, smaller, and with a smoother surface [23,24]. Infections from less severe (mild) isolates of the virus, however, produce only transient leaf symptoms, and sometimes induce marginal stem swellings on affected cacao plants with little effect on yield [25,26]. The occurrence of mixed and co-infections with different species and strains sometimes makes it difficult to attribute visible symptoms to specific species or strains of the virus in field infections. Delineating the impact of individual viruses and/or synergistic effects on the severity of symptom expressions from mixed and co-infections on growth and yields of CSSD-affected cacao plants has not been well studied. Moreover, the possible beneficial role of mixed co-infections in conferring immunity and symptom suppression is poorly understood.
Visual examination of affected or suspected cacao plants and other hosts for evidence of visible leaf and stem symptoms has been the foremost method for field and graft-based recognition of suspected infections in field samples of CSSV disease [27]. This practice involves the constant lookout for identifiable symptoms of CSSV to confirm or reject infection status of suspected diseased plants. Visual inspection is considered ineffective, as latently infected hosts, i.e., infected but not expressing observable symptoms, are sometimes missed, and the suspected cacao plants are counted as non-infected. It is also known that successful detection of an infection with visual inspection might be subjective and may be reliant on the level of expertise of the inspector, as well as the physiological condition of the suspected host at the time of inspection [27]. Furthermore, results from visual inspection could be unreliable as they could be prejudiced by nutritional deficiencies and other physiological factors of the affected host plants which may cause somewhat similar leaf symptoms analogous to virus infection. Constant observation for appearance of visible symptoms is also time-consuming, as suspected disease plants sometimes to need to be grafted onto healthy counterparts (Amelonado cacao cultivars) and frequently monitored and assessed for a period of not less than 3 years before an infective status can be confirmed or rejected.
Consequently, there is the need for a quicker and more reliable detection system for the virus, and the lack of such efficient diagnostics techniques has been a major limitation with regards to CSSV epidemiology and management. Currently, there is a drift towards molecular diagnostic tools, as the efficiency, repeatability, sensitivity, and reliability associated with molecular detection methods are constantly being improved with modern techniques and protocols which tend to lower the detection time. Molecular detection procedures provide faster results which accordingly enables more field samples to be assessed within a shorter period for prompt decision making without necessarily depending on visible symptom expression. Presently, there is a lack of clarity on the interaction among biological processes of the key cacao badnavirus species, the occurrence of mixed infections, and the influence of environmental parameters on yield loss of affected cacao plants. Similarly, there is limited information on the impact of cacao virus mixed infection on virus titer loads and symptom expression. Ultimately, there is the need for advanced scientific investigations for better elucidation of the epidemiological importance of the highlighted gaps in knowledge on CSSD symptom severity and yield decline.
7. CSSD Alternative Host Plants Research
Evidence from earlier research activities suggested that CSSV might have originated from wild forest and indigenous trees within the cacao ecosystem [37]. It was considered that the virus might have crossed over into the cacao plant when it was first introduced into the various growing regions [37,38]. These assertions were based on earlier investigations on alternative host plants in the Western Region of Ghana which suggested that CSSV infections into cacao plants possibly originated from Cola chlamydantha (K. Schum) trees [37]. It was reported that C. chlamydantha trees were found infected with CSSV, both within cacao farms and miles away in forest reserves [37,38]. These wild (forest tree) hosts were thus assumed to have served as potential sources (reservoirs) of inoculum of the virus to the prevailing and wandering mealybug insect vectors, and were probably the original hosts of this virus before cacao was introduced into West Africa [37]. Subsequently, other tree species that were identified as plausible wild alternative hosts of the virus included Erythropsis barteri (Mast), Sterculia trangacantha (Lindle), Sterculia rhinopetala (K. Schum), Cola gigantea var. glabrescens, (Bronnan et Keay), Adansonia digitata (L.), Bombax buonopozense, and Ceiba pentandra (L.). The most common wild hosts in the cacao plantations in Ghana included C. pentandra (L.), A. digitata, and C. gigantea [38]. These were considered the most significant natural sources of infection, as these wild tree species occurred predominantly in all the cacao growing areas of Ghana at the time of these investigations [38]. It is, however, noteworthy to mention that not all the suspected wild hosts could be considered good sources of virus. Accessibility of the virus inoculum to the mealybug vectors declines to a low level in these alternative hosts and might not be readily available to mealybugs. Coppicing (cutting off the tops of the trees near the base of the crown) of the matured alternative host plants resulted in a temporary upsurge in virus concentration and availability to the mealybug vectors for transmission from the wild hosts to cacao plants and vice versa [38].
Based on the initial evidence of the potential involvement of the wild (alternative) host plants in CSSV spread, it was therefore recommended that the implicated wild host plants should thus be promptly removed as much as possible from cacao plantations [37,38]. This recommendation has not always been fully adopted by the farmers and stakeholders involved in CSSD management programs, and is thus considered as one of the main challenges that accounts for the continuous spread of the virus in the field [37,38]. Updated information on the status of the many forest trees species, shrubs, and weed species in the cacao ecosystem as plausible alternative hosts or otherwise remain critical for the continuous implementation of a sustainable integrated management strategy for the disease.
8. Germplasm Screening for Tolerant/Resistant Planting Materials
Availability of resistant/tolerant cacao varieties for establishment of new cacao plantations has long been suggested as the most feasible strategy and surest means to minimize CSSV spread in the field [39]. This strategy is anticipated to reduce the impact of the disease and the damage it causes to affected cacao plants as a long-term management for the virus [39]. Based on this assertion, series of germplasm screening tests have been carried out, involving local cacao cultivars and others introduced from a germplasm collection maintained in Trinidad. Thus far, none of the tested cacao varieties have been reported to be immune or resistant to the virus. It is, however, known that hybrids of Upper Amazon parentage were more difficult to infect with the virus than the local Amelonado variety [40]. Even when infected, the Upper Amazon hybrids developed only mild symptoms, thus indicating some degree of tolerance to infection [40,41].
The Upper Amazon varieties were therefore introduced in the early 1950s and multiplied for farmers to be used in new plantings to substitute for the Amelonado variety, which hitherto had been the foremost cultivars grown by the farmers [39]. These varieties were later found to have low tolerance to CSSV [41]. The F3 Upper Amazon and the later series II hybrids (Upper Amazon X x Amelonado and Upper Amazon X x Trinitario) were to a lesser extent susceptible than the Amelonado variety. These hybrids performed well against the hostile effects of the virus, even though none of the parents was specifically selected for resistance against CSSV. Further greenhouse and field trials were performed in the search for tolerant or resistant progenies in Ghana and Nigeria, respectively [42,43]. It was reported in these studies that the Parinari genotype and its hybrids showed a characteristic necrotic leaf symptom in Nigeria, which was considered not to be a true hypersensitivity, while Amelonado, Morona, and Trinitario clones were noted to be highly susceptible to the virus [44,45]. The Upper Amazon and Scavina clones were less susceptible than Nanay and Iquitos types, which were only slightly susceptible. It was found in further studies in Ghana that progeny of Trinidad pod T17 of Amazon parentage (Iquitos) might be resistant to the virus, as previously suggested in some earlier studies [46,47].
Thus, resistance breeding programs became the main research focus of the British Research Team (BRT) sent to Ghana between 1969 and 1978 to help reverse the CSSV-induced decline of the cacao industry with resistant and tolerant cacao varieties of the virus [43]. Through their sustained research efforts, some hybrids of Inter-Upper Amazon parents involving their male parents were selected in 1978 for resistance to CSSV, even though selection of female parents was constrained by the need to use existing seed gardens [47,48]. These Inter-Upper Amazon hybrids were generally more resistant to CSSV infection than the equivalent series II hybrids, and were therefore recommended for farmers to use in new cacao plantings [48]. Subsequent studies suggested that the level of resistance of these varieties to CSSV was not adequate for long term protection from infection of the virus [49]. The search for cacao varieties with improved resistance to the virus, through screening of existing and new cacao germplasm, mutation breeding, tissue culture techniques, and other modern breeding tools, has been a constant research priority at CRIG [49,50,51]. This is aimed at developing high performing varieties which can withstand the prevalence of high virus inoculum to complement efforts at integrated management of the virus in the field [50,51,52].
9. Mild Strain cross Protection Research
Observations from earlier research activities indicated that young cacao trees found around outbreaks of virulent virus isolates (New Juabeng strain) could be protected with mild forms of CSSV [53,54,55]. It was thus suggested that a mild strain protection method could be applicable in areas where the disease was highly prevalent and spreading uncontrollably [53]. Nonetheless, the main concern at that time was that mild virus might mutate after a period to cause serious devastation [54]. Mild strain protection approach was therefore considered not to be compatible with the intention to treat all outbreaks by cutting out sources of infection [56]. It was thus suggested that comprehensive research need to be carried out on the behavior and dissemination of the mild strains and their long-term protective effect on cacao [56].
Nonetheless, further research on mild strain research at CRIG has not progressed as expected, due to limited information and other concerns about the available mild strain-inoculated cacao trees being able to harbor severe strains at the same time [55], and also the probability of mild strains being mutated to change from less severe to severe strains in the field [57], as well as the fear of mild strain protection being temporary, and the challenges in differentiating between field symptoms induced by mild and severe strains to warrant prompt eradication of severe field infections [56]. These concerns and challenges were supposed to be investigated through long-term on-station trials at CRIG before a conclusive recommendation could be made for large-scale application by farmers [57]. Available information from the on-station investigations indicates that the apparent protection by the two main mild strains (i.e., N1 and SS365B) in conferring immunity to cacao plants breaks down between 15 years and 20 years post inoculation [58,59]. This finding suggests the need for repeated mild strain inoculations for the sustainable and long-term protection of cacao plants in the field [57,58,59]. Issues about the need for the development of inoculation approaches that would ensure efficient introduction of the mild virus into cacao seedlings prior to distribution and planting were highlighted in these reports. Collectively, this updated research information supports the need for better clarity of the status of the mild strain phenomenon and its associated challenges before its adoption as a management strategy for the virus in the field.
11. Environmental and Edaphic Factors on CSSD Epidemiology
Soil nutrition is considered one of the manageable factors of the environment that have been postulated to influence plant resistance or tolerance to pathogenic attack [68]. Nonetheless, the possible effects of soil nutrients on CSSD symptom expressions have not been adequately clarified, with several unanswered questions about the role of soil nutrition on CSSD epidemiology [69]. Similarly, there is inadequate evidence on the role of environmental parameters, such as level of shade, and their interactive effect with soil nutrient status on CSSD epidemiology. Although the apparent positive effect of adequate level of shade (i.e., 30 to 50% light penetration) in reducing severity of CSSD symptoms was previously reported [59,70], limited studies have subsequently been carried out on this subject. Thus far, the combined effect of soil nutrient status and shade level on CSSD remains poorly understood. It is therefore imperative that a controlled field experiment is initiated to understand the interactive effect of soil nutrient status and environmental factors on CSSD-affected cacao plants across different agroecological zones in Ghana and the other West African cacao-producing nations.
12. Discussion
This paper reviewed and outlined past and current research interventions that have elucidated the identity of cacao badnaviruses involved in field infections and their impact on CSSD prevalence, symptom severity, and cacao production in West Africa. Emphasis was placed on the prevailing knowledge gaps associated with the biological processes of the known virus species, their diagnosis, effect on disease progress, and prevalence. These information gaps emanate from limited scientific research in these thematic areas, due to inadequate funding and limited laboratory tools as submitted by [66]. The paper thus proposes sustained research interventions to investigate the existing knowledge gaps to guide decisions on effective field diagnostics and management of the disease [23]. Strategically, these investigations need to target key research areas for clarity on the interaction between virus identity, host genotype–virus interactions, and the effects of environmental parameters on disease progress. Similarly, the role of alternative hosts, vector population dynamics, and their impact on CSSD spread and symptom severity require further investigation. Optimization of molecular diagnostic techniques for efficient, reliable, and repeatable detection of the known cacao badnavirus species and their evolutionary pattern also require urgent research attention [22].
Thus far, the key CSSD badnavirus species identified to be predominating in field infections in Ghana and Côte d’Ivoire include Cacao swollen shoot Togo A virus (Cacao swollen shoot Togo A virus; CSSTAV), Cacao swollen shoot Togo B virus (Cacao swollen shoot Togo B virus; CSSTBV), Cacao swollen shoot CD virus (Cacao swollen CD virus; CSSCDV), Cacao swollen shoot CE virus (Cacao swollen shoot CE virus; CSSCEV), and Cacao swollen shoot Ghana M virus (Cacao swollen shoot Ghana M virus; CSSGMV) [18,19,20]. As highlighted in this paper, the evolutionary pattern and epidemiological dynamics associated with these dominant cacao badnavirus species relative to the occurrence of alternative hosts in the cacao landscape remains poorly understood. It is hypothesized that many of these species might have spread from historical exchange of infected planting material and vector-infested cacao pods on farms, albeit this has not been confirmed through systematic research [18]. This assertion therefore requires further scientific investigations and confirmation with modern molecular techniques and enhanced bioinformatic tools. Better clarity on the identity of CSSD badnavirus species/strains, mild strains, and their evolutionary pattern relative to geographical distribution, species diversity, and mixed co-infections would ultimately complement molecular diagnostics and field management of the disease [18].
Investigations on vector transmission efficiency have been complex and laborious to perform and time-consuming, hence the resultant wide gap in knowledge on the various dynamics of mealybug transmission of CSSD badnaviruses in the cacao ecosystem. The interactive role of mealybug population dynamics and host factors for efficient transmission in the field have not been well clarified and thus need to be further investigated. There is limited information on the role of endogenous viral elements (EVEs) [21] on CSSD epidemiology, and this requires further investigations through a combination of the various –omics techniques and field experimentation. Availability of research information will elucidate the infectious status and pathogenicity of the identified cacao EVEs on vertical transmission of CSSD, as documented in other field crops such as bananas [66,67,71]. Clarifying the pathogenic status of cacao EVEs is essential to guide future research and quarantine decisions on cacao planting material (seedling and pod) movement, breeding programs, and certification of cacao seedlings for field planting.
Understanding the role of soil nutrient status and their interactive effect with other environmental parameters such as level of shade on the epidemiology of the cacao swollen shoot virus disease (CSSD) remains a research priority across the different agroecological zones. Although past studies have reported the positive effect of adequate level of shade in reducing CSSD symptom severity in Ghana [70], there is limited information on the combined effect of soil nutrient status and shade level on CSSD epidemiology in the medium to long term. Similarly, the influence of farm management and cultural practices on CSSD progress has not been adequately clarified from past research. Consequently, further research interventions are thus advocated on the influence of soil nutrition and shade level on CSSD incidence and symptom progress in mixed cocoa hybrids for better clarity on this subject. This will invariably complement the integrated management strategies for the virus for sustainable cacao production across the different agroclimatic zones in West Africa. Furthermore, application of modern technologies such as artificial intelligence, robotics technology, big dataset algorithms, image acquisition technologies, and remote sensing tools is advocated for CSSD surveillance, early detection, and prompt management [72].
13. Conclusions
This paper advocates for better clarity on the role of molecular properties of the existing cacao virus species, their geographical distribution, their prevalence, the influence of environmental (abiotic) factors, and their overall impact on CSSD epidemiology and diagnostics in the field. To this end, comprehensive CSSD badnavirus characterization and diversity research with modern tools aimed at gaining better understanding of the identity of key virus species, vector population dynamics, alternative hosts, and cacao genotypes predominating in current field infections is necessary. Further investigations on effects of environmental and soil nutrient factors and their role on occurrence of differential outbreaks across the different geographic regions of Ghana and Côte d’Ivoire remain a research priority. Thus, a multidisciplinary research strategy on the virus and the vectors involving multinational collaborators is hereby proposed to address the prevailing knowledge gaps and challenges in CSSD diagnostics and field management for sustainable cacao production.
Author Contributions
G.A.A. (Conceptualization of first draft preparation, editing), O.D. (manuscript reviewing and editing), E.G. (manuscript reviewing and editing). All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by Cocoa Research Institute of Ghana (CRIG).
Institutional Review Board Statement
All co-authors have consented to submission. This paper is submitted and published with the permission of Executive of CRIG (CRIG/04/2023/042/014). No other consent is required.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon reasonable request from the corresponding author.
Acknowledgments
The contribution of numerous scientists at the CRIG in reviewing the initial drafts of the manuscript is duly acknowledged. The authors are also grateful to Rev. Edwin Afari (CHED), the staff of the Cartography Unit of CHED, and Farouk Nyame, Bunso Cocoa College (BCC), and formerly of CHED for permission to use the current CSSD map as documented in the ongoing third country-wide survey in Ghana.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- Muller, E. Cacao Swollen Shoot Virus (CSSV) History, Biology, and Genome. In Cacao Diseases—A History of Old Enemies and New Encounters; Bailey, B.A., Meinhard, L.W., Eds.; Springer International Publishing: New York, NY, USA, 2016; pp. 337–358. ISBN 978-3-319-247887-8. [Google Scholar] [CrossRef]
- Steven, W.H. A new disease of cacao in the Gold Coast. Gold Coast Farmer 1936, 5, 122–144. [Google Scholar]
- Domfeh, O.; Dzahini-Obiatey, H.K.; Ameyaw, G.A.; Abaka-Ewusie, K.; Opoku, G. Cocoa swollen shoot disease situation in Ghana: A review of current trends. Afr. J. Agric. Res. 2011, 6, 5033–5039. [Google Scholar]
- Ameyaw, G.A. Management of the cacao swollen shoot virus (CSSV) menace in Ghana: The past, present and the future. In Plant Diseases—Current Threats and Management Trends; Topolovec-Pintaric, S., Ed.; IntechOpen: London, UK, 2019; pp. 1–13. ISBN 978-1-78985-116-8. [Google Scholar] [CrossRef]
- Ameyaw, G.A.; Dzahini-Obiatey, H.K.; Domfeh, O. Perspectives on cocoa swollen shoot virus disease (CSSVD) management in Ghana. Crop Prot. 2014, 65, 64–70. [Google Scholar] [CrossRef]
- Posnette, A.F. Transmission of swollen shoot disease of cacao. Trop. Agric. 1940, 17, 98. [Google Scholar]
- Brunt, A.A.; Kenten, R.K.; Nixon, H.L. Some properties of Cacao swollen shoot virus. J. Gen. Microbiol. 1964, 36, 303–309. [Google Scholar] [CrossRef]
- Adomako, D.; Lesemann, D.-E.; Paul, H.L.; Owusu, G.K. Improved methods for the purification and detection of cacao swollen shoot virus. Ann. Appl. Biol. 1983, 103, 109–116. [Google Scholar] [CrossRef]
- Sagemann, W.; Lesemann, D.E.; Paul, H.L.; Adomako, D.; Owusu, G.K. Detection and comparison of some Ghanaian isolates of cocoa swollen shoot virus (CSSV) by enzyme-linked immunosorbent assay (ELISA) and immunoelectron microscopy (EM) using an antiserum to CSSV strain 1A. J. Phytopathol. 1985, 114, 78–89. [Google Scholar] [CrossRef]
- Kenten, R.H.; Legg, J.T. Serological relationship of some viruses from cocoa (Theobroma cacao L.) in Ghana. Ann. Appl. Biol. 1971, 67, 195. [Google Scholar] [CrossRef]
- Kenten, R.H.; Legg, J.T. Methods of assessing the tolerance and resistance of different types of cocoa to CSSV. Ann. Appl. Biol. 1970, 65, 419–424. [Google Scholar] [CrossRef]
- Lot, H.; Djiekpor, E.; Jacquemond, M. Characterization of the genome of cacao swollen shoot virus. J. Gen. Virol. 1991, 72, 1735–1739. [Google Scholar] [CrossRef]
- Muller, E.; Sackey, S. Molecular variability analysis of five new complete cacao swollen shoot virus genomic sequences. Arch. Virol. 2005, 150, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Hagen, L.S.; Jacquemond, M.; Lepingle, A.; Lot, H.; Tepfer, M. Nucleotide sequence and genomic organization of cacao swollen shoot virus. Virology 1993, 196, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Jacquot, E.; Hagen, L.S.; Jacquemond, M.; Lot, P. The open reading frame 2 product of cacao swollen shoot badnavirus is a nucleic acid binding protein. Virology 1996, 225, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Gyamera, E.A.; Domfeh, O.; Ameyaw, G.A. Cacao Swollen Shoot Viruses in Ghana. Plant Dis. 2023, 107, 1261–1278. [Google Scholar] [CrossRef]
- Muller, E.; Ravel, S.; Agret, C.; Abrokwah, F.; Dzahini-Obiatey, H.; Galyuon, I.; Kouakou, K.; Jeyaseelan, E.C.; Allainguillaume, J.; Wetten, A. Next generation sequencing elucidates cacao badnavirus diversity and reveals the existence of more than ten viral species. Virus Res. 2018, 244, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Chingandu, N.; Dongo, L.; Gutierrez, O.A.; Brown, J.K. Molecular characterization and detection of Cacao red vein-banding virus, a previously unidentified badnavirus species associated with cacao swollen shoot disease in Nigeria. Plant Dis. 2019, 103, 1302–1308. [Google Scholar] [CrossRef]
- Abrokwah, F.K.; Dabie, K.; Adulley, F. Genome variability, species diversity, phylogenetic relationships, origin, and geographical distribution of badnaviruses involved in the cacao swollen shoot disease: The case of West Africa. Trop. Plant Pathol. 2022, 47, 201–213. [Google Scholar] [CrossRef]
- Ramos-Sobrinho, R.; Kouakou, K.; Bi, A.B.; Keith, C.V.; Diby, L.; Kouame, C.; Aka, R.A.; Marelli, J.-P.; Brown, J.K. Molecular detection of cacao swollen shoot badnavirus species by amplification with four PCR primer pairs, and evidence that Cacao swollen shoot Togo B virus-like isolates are highly prevalent in Côte d’Ivoire. Eur. J. Plant Pathol. 2021. [Google Scholar] [CrossRef]
- Muller, E.; Ullah, I.; Dunwell, J.M.; Daymond, A.J.; Richardson, M.; Allainguillaume, J.; Wetten, A. Identification and distribution of novel badnaviral sequences integrated in the genome of cacao (Theobroma cacao). Sci. Rep. 2021, 11, 8270. [Google Scholar] [CrossRef]
- Ameyaw, G.A.; Domfeh, O.; Armooh, B.; Boakye, A.Y.; Arjarquah, A. Inconsistent PCR detection of Cacao swollen shoot virus (CSSV) is linked to the occurrence of different variants across the cocoa regions of Ghana. J. Virol. Methods 2021, 300, 114400. [Google Scholar] [CrossRef]
- Posnette, A.F. Control measures against swollen shoot virus disease of cacao. Trop. Agric. 1943, 20, 116–123. [Google Scholar]
- Thresh, J.M.; Owusu, G.K.; Boamah, A.; Lockwood, G. Ghanaian cocoa varieties and swollen shoot virus. Crop Prot. 1988, 7, 219–231. [Google Scholar] [CrossRef]
- Ameyaw, G.A.; Domfeh, O.; Dzahini-Obiatey, H.; Ollennu, L.A.A.; Owusu, G.K. Appraisal of Cocoa swollen shoot virus (CSSV) mild isolates for cross protection of cocoa against severe strains in Ghana. Plant Dis. 2016, 100, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Domfeh, O.; Ameyaw, G.A.; Boakye, A.Y.; Opoku-Agyeman, M.O.; Arthur, A.; Akpertey, A.; Padi, F.K.; Dzahini-Obiatey, H.K. Evaluation of mild strain cross protection in cacao—Further evidence of the protective potential of cacao swollen shoot virus strain N1 against the New Juabeng (1A) isolate under field conditions. Australas. Plant Pathol. 2021, 50, 329–340. [Google Scholar] [CrossRef]
- Ollennu, L.A.A. Synthesis: Case history of cocoa viruses. In Proceedings of the Conference on Plant Virology in Sub-Saharan Africa, Ibadan, Nigeria, 4–8 June 2001; pp. 33–39. [Google Scholar]
- Posnette, A.F.; Strickland, A.H. Virus diseases of cacao in West Africa. III. Technique of insect transmission. Ann. Appl. Biol. 1948, 35, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Roivainen, O. Transmission of cacao viruses by mealybugs (Homoptera: Pseudococcidae). J. Sci. Agric. Soc. Finl. 1976, 48, 203–204. [Google Scholar]
- Entwistle, P.F.; Longworth, J.F. The relationship between cocoa viruses and their vectors: The feeding behaviour of three mealybug species (Homoptera: Pseudocoidia). Ann. Appl. Biol. 1963, 52, 388–391. [Google Scholar] [CrossRef]
- Strickland, A.H. The dispersal of pseudococcidae (Hemiptera-Homoptera) by air currents in the Gold Coast. Proc. R. Entomol. Soc. Lond. 1950, 25, 1–9. [Google Scholar] [CrossRef]
- Hanna, A.D. Research in the use of systemic insecticides to control the mealybug vectors of swollen shoot disease of cocoa. Phytiat-Phytophram 1954, 425–432. [Google Scholar]
- Bowmann, J.S.; Casida, J.E. Systemic insecticides for Theobroma cacao L.; their translocation and persistence in foliage and residue in cocoa beans. J. Econ. Entomol. 1958, 51, 773–780. [Google Scholar] [CrossRef]
- Donald, R.G. Mealybug Studies. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 1955; p. 101. [Google Scholar]
- Decker, F.E. Mealybug Studies: Biological Control of Mealybugs. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 1955; p. 23. [Google Scholar]
- Ackonor, J.B. Preliminary studies on breeding and predation on Scymnus (Pullus) sp. and Hyperaspis egregia Mader on Planococcoides njalensis (Laing). In Proceedings of the First International Cocoa Pests and Diseases Seminar, Accra, Ghana, 6–10 November 1995; pp. 238–241. [Google Scholar]
- Posnette, A.F.; Robertson, N.F.; Todd, J.M. Virus diseases of cocoa in West Africa. V. Alternative host plants. Ann. Appl. Biol. 1950, 37, 229–240. [Google Scholar] [CrossRef]
- Posnette, A.F. The role of wild hosts. In Pests, Pathogens, and Vegetation; Thresh, J.M., Ed.; Pitman: London, UK, 1981; pp. 71–78. [Google Scholar]
- Posnette, A.F.; Todd, J.M. Virus diseases of cacao in West Africa VIII. The search for virus-resistant cacao. Ann. Appl. Biol. 1951, 38, 785–800. [Google Scholar] [CrossRef]
- Attafuah, A.; Glendinning, D.R. Studies on resistance and tolerance to cocoa viruses in Ghana: A survey of T17 progeny. Ann. Appl. Biol. 1965, 56, 219. [Google Scholar] [CrossRef]
- Dale, W.T.; Attafuah, A. The host range of cocoa viruses. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 1956; pp. 28–30. [Google Scholar]
- Blencowe, J.W.; Attafuah, A. Resistance and tolerance. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 1959; p. 18. [Google Scholar]
- Longworth, J.F.; Thresh, J.M. Field trial on the effect of a Nigerian swollen shoot virus on the growth of different cocoa types. Ann. Appl. Biol. 1963, 52, 217. [Google Scholar]
- Brunt, A.A.; Kenten, R.H. Viruses infecting cacao. Rev. Plant Pathol. 1971, 50, 591–602. [Google Scholar]
- Dale, W.T. Virus research: Resistance and tolerance. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 1957; pp. 35–36. [Google Scholar]
- Posnette, A.F. Viruses of cocoa in West Africa: 1. Cocoa viruses 1A, 1B, 1C, and 1D. Ann. Appl. Biol. 1947, 34, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Legg, J.T.; Lockwood, G. Resistance of cocoa to swollen shoot virus in Ghana. I. Field trials. Ann. Appl. Biol. 1981, 97, 75–89. [Google Scholar] [CrossRef]
- Adu-Ampomah, Y. The cocoa breeding program in Ghana: Achievements and prospects for the future. Cocoa Growers’ Bull. 1996, 50, 17–21. [Google Scholar]
- Padi, F.K.; Domfeh, O.; Takrama, J.; Opoku, S.Y. An evaluation of gains in breeding for resistance to the cacao swollen shoot virus disease in Ghana. Crop Prot. 2013, 51, 24–31. [Google Scholar] [CrossRef]
- Ofori, A.; Padi, F.K.; Ameyaw, G.A.; Dadzie, A.M.; Lowor, S. Genetic variation among cacao (Theobroma cacao L.) families for resistance to cacao swollen shoot virus disease in relation to total phenolic content. Plant Breed. 2015, 134, 477–484. [Google Scholar] [CrossRef]
- Adomako, B.; Opoku, S.Y.; Dadzie, A.M.; Adu-Ampomah, Y. Breeding for cacao varieties resistant/tolerant to CSSV. In Annual Report of West African Cocoa Research Institute; West African Cocoa Research Institute: Tafo, Ghana, 2008; pp. 97–100. [Google Scholar]
- Ofori, A.; Padi, F.K.; Ameyaw, G.A.; Dadzie, A.M.; Opoku-Agyeman, M.; Domfeh, O.; Owusu-Ansah, F. Field evaluation of the impact of cacao swollen shoot virus disease infection on yield traits of different cacao (Theobroma cacao L.) clones in Ghana. PLoS ONE 2022, 17, e0262461. [Google Scholar] [CrossRef] [PubMed]
- Posnette, A.F.; Todd, J.M. Virus diseases of cocoa in West Africa. IX. Strain variation and interference in virus 1A. Ann. Appl. Biol. 1955, 43, 433. [Google Scholar] [CrossRef]
- Crowdy, S.H.; Posnette, A.F. Virus diseases of cacao in West Africa. II. Cross-immunity experiments with viruses IA, IB and IC. Ann. Appl. Biol. 1947, 34, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Ollennu, L.A.A.; Owusu, G.K.; Dzahini-Obiatey, H. Recent studies of mild strain cross protection with cocoa swollen shoot virus. J. Ghana Sci. Assoc. 1999, 2, 5–11. [Google Scholar]
- Ollennu, L.A.A.; Owusu, G.K.; Thresh, J.M. The control of cocoa swollen shoot disease in Ghana. Cocoa Growers’ Bull. 1989, 42, 25–35. [Google Scholar]
- Domfeh, O.; Ameyaw, G.A.; Awudzi, G.K.; Arthur, A.; Ofori, A.; Anokye, E.; Gyamera, E.A.; Padi, F.K. Disease reaction of cacao progenies following inoculation with the cacao swollen shoot Togo B virus (CSSTBV) under field conditions. Trop. Plant Pathol. 2023, 48, 703–712. [Google Scholar] [CrossRef]
- Owusu, G.K.; Ollennu, L.A.A.; Dzahini-Obiatey, H. The prospects for mild strain cross-protection to control cocoa swollen shoot disease in Ghana. In Proceedings of the 12th International Cocoa Research Conference, Salvador de Bahia, Brazil, 17–23 November 1996; pp. 121–127. [Google Scholar]
- Domfeh, O.; Ameyaw, G.A.; Gyamera, E.A. Mild strain cross-protection in cacao: Evaluation of symptom suppression in three progenies exposed to natural spread of the New Juabeng strain of the Cacao swollen shoot Togo B virus (CSSTBV). Trop. Plant Pathol. 2022, 47, 562–568. [Google Scholar] [CrossRef]
- Hull, R. Matthews’ Plant Virology, 4th ed.; Academic Press: London, UK, 2002; p. 981. [Google Scholar]
- Hughes, J.D.; Adomako, D.; Ollennu, L.A.A. Evidence from the virobacterial agglutination test for the existence of eight serogroups of cocoa swollen shoot virus. Ann. Appl. Biol. 1995, 127, 297–307. [Google Scholar] [CrossRef]
- Sackey, S.T.; Dzahini-Obiatey, H.K.; Owusu, G.K.; Adomako, D. Variation in cocoa swollen shoot virus isolates. In Proceedings of the 12th International Cocoa Research Conference, Salvador, Brazil, 17–23 November 1996; pp. 107–112. [Google Scholar]
- Gambley, C.F.; Geering, A.D.W.; Thomas, J.E. Development of an immunomagnetic capture-reverse transcriptase-PCR assay for three pineapple ampeloviruses. J. Virol. Methods 2009, 155, 187–192. [Google Scholar] [CrossRef]
- Mumford, R.A.; Seal, S.E. Rapid Single-Tube Immunocapture RT-PCR for the Detection of Two Yam Potyviruses. J. Virol. Methods 1997, 69, 73–79. [Google Scholar] [CrossRef]
- Wang, M.; Li, F.; Zhou, G.; Lan, P.; Xu, D.; Li, R. Molecular Detection and Characterization of Chinese Yam Mild Mosaic Virus Isolates. Phytopathology 2015, 163, 1036–1040. [Google Scholar] [CrossRef]
- Umber, M.; Pichaut, J.-P.; Farinas, B.; Laboureau, N.; Janzac, B.; Plaisir-Pineau, K.; Pressat, G.; Baurens, F.-C.; Chabannes, M.; Duroy, P.-O. Marker-Assisted Breeding of Musa Balbisiana Genitors Devoid of Infectious Endogenous Banana Streak Virus Sequences. Mol. Breed. 2016, 36, 74. [Google Scholar] [CrossRef]
- Umber, M.; Pressat, G.; Fort, G.; Plaisir Pineau, K.; Guiougiou, C.; Lambert, F.; Farinas, B.; Pichaut, J.-P.; Janzac, B.; Delos, J.-M. Risk Assessment of Infectious Endogenous Banana Streak Viruses in Guadeloupe. Front. Plant Sci. 2022, 13, 951285. [Google Scholar] [CrossRef] [PubMed]
- Eraslan, F.; Akbas, B.; Inal, A.; Tarakcioglu, C. Effects of foliar sprayed calcium sources on Tomato mosaic virus (ToMV) infection in tomato plants grown in greenhouses. Phytoparasite 2007, 35, 150–158. [Google Scholar] [CrossRef]
- Ameyaw, G.A.; Arthur, A.; Domfeh, O.; Owusu-Ansah, F.; Boakye, A. Assessing the relationship of the three major soil nutrients (NPK) on cacao swollen shoot virus disease (CSSVD) symptom severity in Ghana. Solid State Technol. 2020, 63, 10684–10697. [Google Scholar]
- Andres, C.; Blaser, W.J.; Dzahini-Obiatey, H.K.; Ameyaw, G.A.; Domfeh, O.; Moses, A.A.; Gattinger, A.; Schneider, M.; Offei, S.K.; Six, J. Agroforestry systems can mitigate the severity of cocoa swollen shoot virus disease. Agric. Ecosyst. Environ. 2016, 252, 83–92. [Google Scholar] [CrossRef]
- Chabannes, M.; Baurens, F.-C.; Duroy, P.-O.; Bocs, S.; Vernerey, M.-S.; Rodier-Goud, M.; Barbe, V.; Gayral, P.; Iskra-Caruana, M.-L. Three Infectious Viral Species Lying in Wait in the Banana Genome. Virology 2013, 87, 8624–8637. [Google Scholar] [CrossRef]
- Agbeli, P.; Panford, J.K.; Anokye, K.; Hayfron-Acquah, J.B. Application of Digital Image Processing Technology to Detect Diseases in Cocoa Plants. Master’s Thesis, University of Applied Sciences Schmalkalden, Schmalkalden, Germany, 2022. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).