
Isolation and Characterization of a Bacteriophage with Potential for the Control of Multidrug-Resistant Salmonella Strains Encoding Virulence Factors Associated with the Promotion of Precancerous Lesions

 ,
, 
 , , and
, , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Isolation of Salmonella Strains and Identification of Genes with Oncogenic Potential
2.2. Antibiotic Susceptibility Tests
2.3. Bacteriophage Isolation and Purification
2.4. Transmission Electron Microscopy
2.5. One-Step Replication Curve
2.6. Bacteriophage Host Range
2.7. Stability of the Bacteriophage
2.8. Bacteriolytic Activity of the Bacteriophage
2.9. Genomic Sequencing and Bioinformatic Analysis
2.10. Statistical Analysis
3. Results
3.1. Isolation of Salmonella and Identification of Genes Associated with Cancer Induction
3.2. Antibiotic Sensitivity Tests
3.3. Isolation of the Bacteriophage
3.4. Bacteriophage Morphology
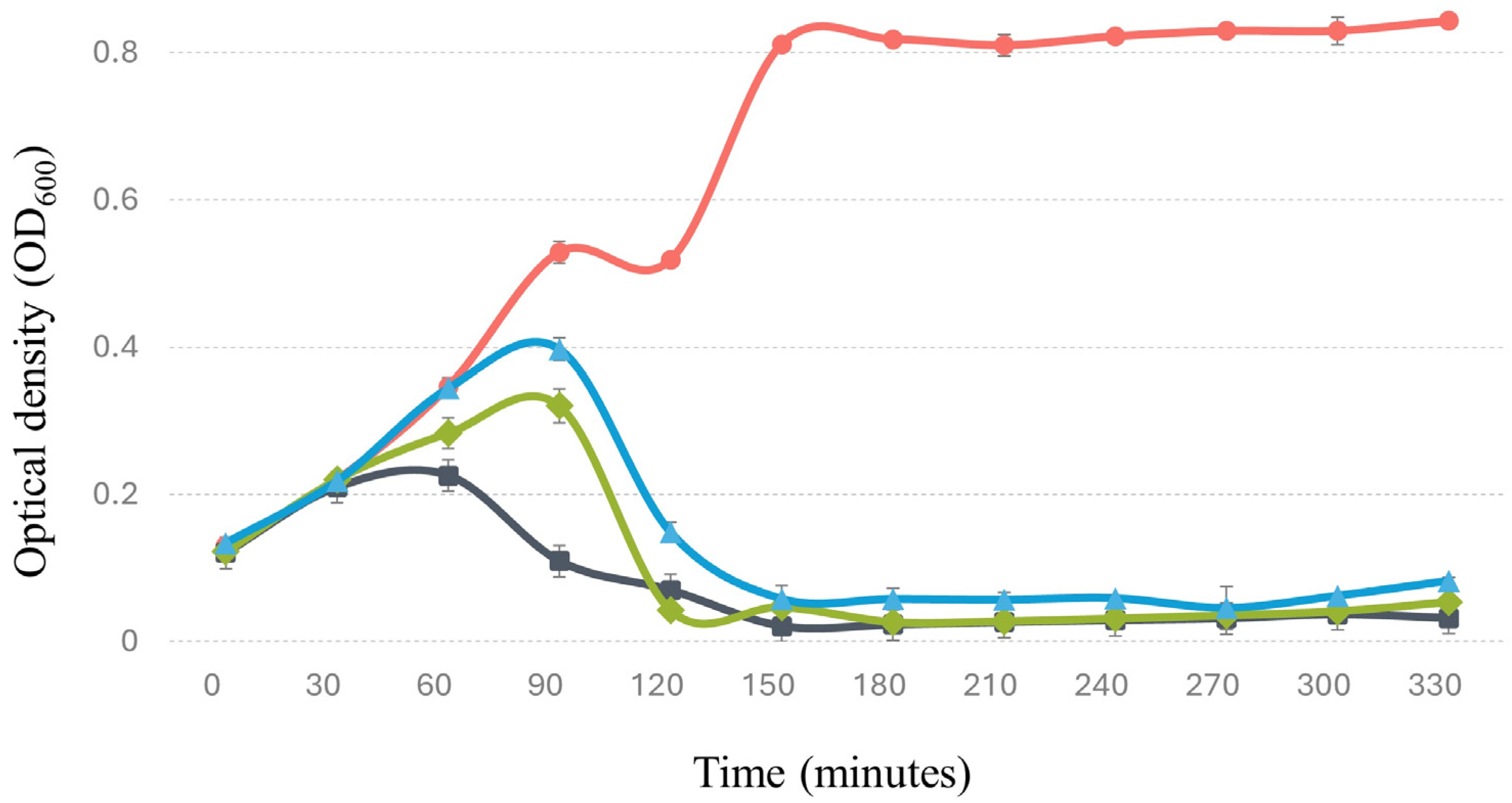
3.5. Bacteriophage Replication Curve
3.6. Host Range
3.7. Stability of the Bacteriophage
3.7.1. Storage Stability
3.7.2. Stability in Simulated Gastrointestinal System
3.8. Capacity of the Bacteriophage to Control Salmonella
3.8.1. In Culture Medium
3.8.2. In Simulated Gastrointestinal System
3.9. Bacteriophage Genomics Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.E.; Han, C.; Hayoon, A.G.; et al. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef] [PubMed]
- Loftus, M.J.; Everts, R.J.; Cheng, A.C.; Eti, P.; Fakasiieiki, T.; Isaia, L.; Isopo, E.; Jenney, A.W.; Lameko, V.; Leaupepe, H.; et al. Antimicrobial susceptibility of bacterial isolates from clinical specimens in four Pacific Island countries, 2017–2021. Lancet Reg. Health—West. Pac. 2022, 32, 100677. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Karvanen, M.; Cars, O. The language of antimicrobial and antibiotic resistance is blocking global collective action. Infect. Dis. 2024, 56, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Heinzel, M.; Koenig-Archibugi, M. National action on antimicrobial resistance and the political economy of health care. J. Eur. Public Policy 2024, 31, 3981–4007. [Google Scholar] [CrossRef]
- WHO. WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance to Guide Research, Development and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2024; pp. 12–13. [Google Scholar]
- Soubeiga, A.P.; Kpoda, D.S.; Compaoré, M.K.; Somda-Belemlougri, A.; Kaseko, N.; Rouamba, S.S.; Ouedraogo, S.; Traoré, R.; Karfo, P.; Nezien, D. Molecular characterization and the antimicrobial resistance profile of Salmonella spp. isolated from ready-to-eat foods in Ouagadougou, Burkina Faso. Int. J. Microbiol. 2022, 2022, 9640828. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, H.; Liu, Y.; Huang, C.; Chen, J.; Siddique, A.; Yin, R.; Jia, C.; Li, Y.; Zhao, G.; et al. Nationwide trends and features of human salmonellosis outbreaks in China. Emerg. Microbes Infect. 2024, 13, 2372364. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, B.; Mawad, A.M.; Saleh, M.; Kelley, W.G.; Harrington, P.J.; Lovestad, C.W.; Amezcua, J.; Sarhan, M.M.; El Zowalaty, M.E.; Ramadan, H. Salmonellosis: An overview of epidemiology, pathogenesis, and innovative approaches to mitigate the antimicrobial resistant infections. Antibiotics 2024, 13, 76. [Google Scholar] [CrossRef]
- Scanu, T.; Spaapen, R.M.; Bakker, J.M.; Pratap, C.B.; Wu, L.E.; Hofland, I.; Broeks, A.; Shukla, V.K.; Kumar, M.; Janssen, H. Salmonella manipulation of host signaling pathways provokes cellular transformation associated with gallbladder carcinoma. Cell Host Microbe 2015, 17, 763–774. [Google Scholar] [CrossRef]
- Lu, R.; Wu, S.; Zhang, Y.G.; Xia, Y.; Zhou, Z.; Kato, I.; Dong, H.; Bissonnette, M.; Sun, J. Salmonella protein AvrA activates the STAT3 signaling pathway in colon cancer. Neoplasia 2016, 18, 307–316. [Google Scholar] [CrossRef]
- de Savornin Lohman, E.; Duijster, J.; Groot Koerkamp, B.; van der Post, R.; Franz, E.; Mughini Gras, L.; de Reuver, P. Severe Salmonella spp. or Campylobacter spp. infection and the risk of biliary tract cancer: A population-based study. Cancers 2020, 12, 3348. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.N.; Ni, J.J.; Li, J.H.; Gao, Y.Q.; Ni, F.J.; Zhang, Z.Z.; Fang, J.Y.; Lu, J.; Yao, Y.F. Bacterial infection promotes tumorigenesis of colorectal cancer via regulating CDC42 acetylation. PLoS Pathog. 2023, 19, e1011189. [Google Scholar] [CrossRef] [PubMed]
- Lopez Chiloeches, M.; Bergonzini, A.; Martin, O.C.; Bergstein, N.; Erttmann, S.F.; Aung, K.M.; Gekara, N.O.; Avila Cariño, J.F.; Pateras, I.S.; Frisan, T. Genotoxin-producing Salmonella enterica induces tissue-specific types of DNA damage and DNA damage response outcomes. Front. Immunol. 2023, 14, 1270449. [Google Scholar] [CrossRef]
- Lu, R.; Bosland, M.; Xia, Y.; Zhang, Y.G.; Kato, I.; Sun, J. Presence of Salmonella AvrA in colorectal tumor and its precursor lesions in mouse intestine and human specimens. Oncotarget 2017, 8, 55104. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Xu, Q.; Cui, Y.; Yao, S.; Jin, S.; Zhang, Q.; Wen, Z.; Ruan, H.; Liang, X.; Chao, Y.; et al. Salmonella effector SopB reorganizes cytoskeletal vimentin to maintain replication vacuoles for efficient infection. Nat. Commun. 2023, 14, 478. [Google Scholar] [CrossRef]
- ElGhazaly, M.; Collins, M.O.; Ibler, A.E.M.; Humphreys, D. Typhoid toxin hijacks Wnt5a to establish host senescence and Salmonella infection. Cell Rep. 2023, 42, 113181. [Google Scholar] [CrossRef]
- Shukla, R.; Shukla, P.; Behari, A.; Khetan, D.; Chaudhary, R.K.; Tsuchiya, Y.; Ikoma, T.; Asai, T.; Nakamura, K.; Kapoor, V.K. Roles of Salmonella typhi and Salmonella paratyphi in gallbladder cancer development. Asian Pac. J. Cancer Prev. 2021, 22, 509–516. [Google Scholar] [CrossRef]
- Mughini-Gras, L.; Schaapveld, M.; Kramers, J.; Mooij, S.; Neefjes-Borst, E.A.; Pelt, W.V.; Neefjes, J. Increased colon cancer risk after severe Salmonella infection. PLoS ONE 2018, 13, e0189721. [Google Scholar] [CrossRef]
- Onallah, H.; Hazan, R.; Nir-Paz, R. Compassionate use of bacteriophages for failed persistent infections during the first 5 years of the Israeli Phage Therapy Center. Open Forum Infect. Dis. 2023, 10, ofad221. [Google Scholar] [CrossRef]
- Zaki, B.M.; Fahmy, N.A.; Aziz, R.K.; Samir, R.; El-Shibiny, A. Characterization and comprehensive genome analysis of novel bacteriophage, vB_Kpn_ZCKp20p, with lytic and anti-biofilm potential against clinical multidrug-resistant Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2023, 13, 1077995. [Google Scholar] [CrossRef]
- Ferreira, R.; Sousa, C.; Gonçalves, R.F.; Pinheiro, A.C.; Oleastro, M.; Wagemans, J.; Lavigne, R.; Figueiredo, C.; Azeredo, J.; Melo, L.D. Characterization and genomic analysis of a new phage infecting Helicobacter pylori. Int. J. Mol. Sci. 2022, 23, 7885. [Google Scholar] [CrossRef]
- Principi, N.; Silvestri, E.; Esposito, S. Advantages and limitations of bacteriophages for the treatment of bacterial infections. Front. Pharmacology 2019, 10, 457104. [Google Scholar]
- Gindin, M.; Febvre, H.P.; Rao, S.; Wallace, T.C.; Weir, T.L. bacteriophage for gastrointestinal health (PHAGE) study: Evaluating the safety and tolerability of supplemental bacteriophage consumption. J. Am. Coll. Nutr. 2019, 38, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Krysiak-Baltyn, K.; Martin, G.J.O.; Gras, S.L. Computational modelling of large scale phage production using a two-stage batch process. Pharmaceuticals 2018, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Castillo, D.; Kauffman, K.; Hussain, F.; Kalatzis, P.; Rørbo, N.; Polz, M.F.; Middelboe, M. Widespread distribution of prophage-encoded virulence factors in marine Vibrio communities. Sci. Rep. 2018, 8, 9973. [Google Scholar] [CrossRef] [PubMed]
- Flint, R.; Laucirica, D.R.; Chan, H.K.; Chang, B.J.; Stick, S.M.; Kicic, A. Stability considerations for bacteriophages in liquid formulations designed for nebulization. Cells 2023, 12, 2057. [Google Scholar] [CrossRef]
- Berkson, J.D.; Wate, C.E.; Allen, G.B.; Schubert, A.M.; Dunbar, K.E.; Coryell, M.P.; Sava, R.L.; Gao, Y.; Hastie, J.L.; Smith, E.M.; et al. Phage-specific immunity impairs efficacy of bacteriophage targeting vancomycin resistant Enterococcus in a murine model. Nat. Commun. 2024, 15, 2993. [Google Scholar] [CrossRef]
- Liu, P.; Ibaraki, M.; Kapoor, R.; Amin, N.; Das, A.; Miah, R. Development of moore swab and ultrafiltration concentration and detection methods for Salmonella typhi and Salmonella paratyphi A in wastewater and application in Kolkata, India and Dhaka, Bangladesh. Front. Microbiol. 2021, 12, 684094. [Google Scholar] [CrossRef]
- Yanestria, S.M.; Rahmaniar, R.P.; Wibisono, F.J.; Effendi, M.H. Detection of invA gene of Salmonella from milkfish (Chanos chanos) at Sidoarjo wet fish market, Indonesia, using polymerase chain reaction technique. Vet. World 2019, 12, 170–175. [Google Scholar] [CrossRef]
- Alshaheeb, Z.A.; Thabit, Z.A.; Oraibi, A.G.; Baioumy, A.A.; Abedelmaksoud, T.G. Salmonella enterica species isolated from local foodstuff and patients suffering from foodborne illness: Surveillance, antimicrobial resistance and molecular detection. Theory Pract. Meat Process. 2023, 8, 112–123. [Google Scholar] [CrossRef]
- Hawwas, H.A.E.H.; Aboueisha, A.K.M.; Fadel, H.M.; El-Mahallawy, H.S. Salmonella serovars in sheep and goats and their probable zoonotic potential to humans in Suez Canal Area, Egypt. Acta Vet. Scand. 2022, 64, 17. [Google Scholar] [CrossRef] [PubMed]
- Mezal, E.H.; Bae, D.; Khan, A.A. Detection and functionality of the CdtB, PltA, and PltB from Salmonella enterica serovar Javiana. Pathog. Dis. 2014, 72, 95–103. [Google Scholar] [PubMed]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests, 14th ed.; CLSI standard M02; Clinical and Laboratory Standards Institute: Berwyn, PA, USA, 2024. [Google Scholar]
- Ikner, L.A.; Soto-Beltran, M.; Bright, K.R. New method using a positively charged microporous filter and ultrafiltration for concentration of viruses from tap water. Appl. Environ. Microbiol. 2011, 77, 3500–3506. [Google Scholar] [CrossRef]
- Li, C.; Yuan, X.; Li, N.; Wang, J.; Yu, S.; Zeng, H.; Zhang, J.; Wu, Q.; Ding, Y. Isolation and characterization of Bacillus cereus phage vb_bcep-dlc1 reveals the largest member of the Φ29-like phages. Microorganisms 2020, 8, 1750. [Google Scholar] [CrossRef]
- Le Guellec, S.; Pardessus, J.; Bodier-Montagutelli, E.; L’hostis, G.; Dalloneau, E.; Piel, D.; Samaï, H.C.; Guillon, A.; Mujic, E.; Guillot-Combe, E. Administration of bacteriophages via nebulization during mechanical ventilation: In vitro study and lung deposition in macaques. Viruses 2023, 15, 602. [Google Scholar] [CrossRef]
- Kabwe, M.; Brown, T.; Speirs, L.; Ku, H.; Leach, M.; Chan, H.T.; Petrovski, S.; Lock, P.; Tucci, J. Novel bacteriophages capable of disrupting biofilms from clinical strains of Aeromonas hydrophila. Front. Microbiol. 2020, 11, 194. [Google Scholar] [CrossRef]
- Xu, Z.; Ding, Z.; Zhang, Y.; Liu, X.; Wang, Q.; Shao, S.; Liu, Q. Shelf-life prediction and storage stability of Aeromonas bacteriophage vB_AsM_ZHF. Virus Res. 2023, 323, 198997. [Google Scholar] [CrossRef]
- Mulet-Cabero, A.I.; Egger, L.; Portmann, R.; Ménard, O.; Marze, S.; Minekus, M.; Le Feunteun, S.; Sarkar, A.; Grundy, M.M.L.; Carrière, F.; et al. A standardised semi-dynamic: In vitro digestion method suitable for food-an international consensus. Food Funct. 2020, 11, 1702–1720. [Google Scholar] [PubMed]
- Sonalika, J.; Srujana, A.S.; Akhila, D.S.; Juliet, M.R.; Santhosh, K.S. Application of bacteriophages to control Salmonella enteritidis in raw eggs. Iran. J. Vet. Res. 2020, 21, 221. [Google Scholar]
- Akritidou, T.; Akkermans, S.; Smet, C.; Delens, V.; Van Impe, J.F.M. Effect of food structure and buffering capacity on pathogen survival during in vitro digestion. Food Res. Int. 2023, 164, 112305. [Google Scholar] [CrossRef]
- Cieplak, T.; Soffer, N.; Sulakvelidze, A.; Nielsen, D.S. A bacteriophage cocktail targeting Escherichia coli reduces E. coli in simulated gut conditions, while preserving a non-targeted representative commensal normal microbiota. Gut Microbes 2018, 9, 391–399. [Google Scholar] [PubMed]
- Sambrook, J.; Russell, D.W. Extraction of Bacteriophage λ DNA from Large-scale Cultures Using Proteinase K and SDS. Cold Spring Harb. Protoc. 2006, 1, pdb-prot3972. [Google Scholar] [CrossRef] [PubMed]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing phage genomes for therapeutic applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef]
- Turner, D.; Adriaenssens, E.M.; Tolstoy, I.; Kropinski, A.M. Phage annotation guide: Guidelines for assembly and high-quality annotation. PHAGE Ther. Appl. Res. 2021, 2, 170–182. [Google Scholar] [CrossRef]
- Diemert, S.; Yan, T. Municipal wastewater surveillance revealed a high community disease burden of a rarely reported and possibly subclinical Salmonella enterica serovar derby strain. Appl. Environ. Microbiol. 2020, 86, e00814-20. [Google Scholar] [CrossRef] [PubMed]
- Martone-Rocha, S.; Dropa, M.; da Cruz, B.M.C.; Leite, D.B.M.O.; dos Santos, T.P.; Razzolini, M.T.P. Antimicrobial profile of non-typhoidal Salmonella isolated from raw sewage in the Metropolitan Region of São Paulo, Brazil. J. Infect. Dev. Ctries. 2023, 17, 86–92. [Google Scholar] [CrossRef]
- Dougherty, M.W.; Jobin, C. Intestinal bacteria and colorectal cancer: Etiology and treatment. Gut Microbes 2023, 15, 2185028. [Google Scholar] [CrossRef]
- Ruan, H.; Zhang, Z.; Tian, L.; Wang, S.; Hu, S.; Qiao, J.J. The Salmonella effector SopB prevents ROS-induced apoptosis of epithelial cells by retarding TRAF6 recruitment to mitochondria. Biochem. Biophys. Res. Commun. 2016, 478, 618–623. [Google Scholar] [CrossRef]
- Sepe, L.P.; Hartl, K.; Iftekhar, A.; Berger, H.; Kumar, N.; Goosmann, C.; Chopra, S.; Schmidt, S.C.; Gurumurthy, R.K.; Meyer, T.F. Genotoxic effect of Salmonella paratyphi a infection on human primary gallbladder cells. mBio 2020, 11, 1–39. [Google Scholar] [CrossRef]
- Castellanos, L.R.; van Der Graaf-van Bloois, L.; Donado-Godoy, P.; Veldman, K.; Duarte, F.; Acuña, M.; Jarquín, C.; Weill, F.X.; Mevius, D.; Wagenaar, J.; et al. Antimicrobial resistance in Salmonella enterica serovar paratyphi B variant Java in poultry from Europe and Latin America. Emerg. Infect. Dis. 2020, 26, 1164–1173. [Google Scholar] [CrossRef]
- Amancha, G.; Celis, Y.; Irazabal, J.; Falconi, M.; Villacis, K.; Thekkur, P.; Nair, D.; Perez, F.; Verdonck, K. High levels of antimicrobial resistance in Escherichia coli and Salmonella from poultry in Ecuador. Rev. Panam. De Salud Publica/Pan Am. J. Public Health 2023, 47, e15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chu, M.; Liao, Y.T.; Salvador, A.; Wu, V.C.H. Characterization of two novel Salmonella phages having biocontrol potential against Salmonella spp. in gastrointestinal conditions. Sci. Rep. 2024, 14, 12294. [Google Scholar] [CrossRef] [PubMed]
- Ge, H.; Xu, Y.; Hu, M.; Zhang, K.; Zhang, S.; Chen, X. Isolation, characterization, and application in poultry products of a Salmonella-specific bacteriophage, S55. J. Food Prot. 2021, 84, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Hu, Y.; Mizan, M.F.R.; Yan, T.; Nime, I.; Zhou, Y.; Li, J. Characterization of Salmonella phage LPST153 that effectively targets most prevalent Salmonella serovars. Microorganisms 2020, 8, 1089. [Google Scholar] [CrossRef] [PubMed]
- Teklemariam, A.D.; Alharbi, M.G.; Al-Hindi, R.R.; Alotibi, I.; Aljaddawi, A.A.; Azhari, S.A.; Esmael, A. Isolation and characterization of Chi-like Salmonella bacteriophages infecting two Salmonella enterica Serovars, Typhimurium and Enteritidis. Pathogens 2022, 11, 1480. [Google Scholar] [CrossRef]
- Jurczak-Kurek, A.; Gąsior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Elsayed, M.M.; Elkenany, R.M.; EL-Khateeb, A.Y.; Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Isolation and encapsulation of bacteriophage with chitosan nanoparticles for biocontrol of multidrug-resistant methicillin-resistant Staphylococcus aureus isolated from broiler poultry farms. Sci. Rep. 2024, 14, 4702. [Google Scholar] [CrossRef]
- Wójcicki, M.; Średnicka, P.; Błażejak, S.; Gientka, I.; Kowalczyk, M.; Emanowicz, P.; Świder, O.; Sokołowska, B.; Juszczuk-Kubiak, E. Characterization and genome study of novel lytic bacteriophages against prevailing saprophytic bacterial microflora of minimally processed plant-based food products. Int. J. Mol. Sci. 2021, 22, 12460. [Google Scholar] [CrossRef]
- Unverdi, A.; Erol, H.B.; Kaskatepe, B.; Babacan, O. Characterization of Salmonella phages isolated from poultry coops and its effect with nisin on food bio-control. Food Sci. Nutr. 2024, 12, 2760–2771. [Google Scholar] [CrossRef]
- Jeon, G.; Ahn, J. Evaluation of phage adsorption to Salmonella Typhimurium exposed to different levels of pH and antibiotic. Microb. Pathog. 2021, 150, 12460. [Google Scholar] [CrossRef]
- Shang, Y.; Sun, Q.; Chen, H.; Wu, Q.; Chen, M.; Yang, S.; Du, M.; Zha, F.; Ye, Q.; Zhang, J. Isolation and characterization of a novel Salmonella phage vB_SalP_TR2. Front. Microbiol. 2021, 12, 664810. [Google Scholar] [CrossRef]
- Khan, M.A.S.; Islam, Z.; Barua, C.; Sarkar, M.M.H.; Ahmed, M.F.; Rahman, S.R. Phenotypic characterization and genomic analysis of a Salmonella phage L223 for biocontrol of Salmonella spp. in poultry. Sci. Rep. 2024, 14, 15347. [Google Scholar] [CrossRef] [PubMed]
- Holtappels, D.; Alfenas-Zerbini, P.; Koskella, B. Drivers and consequences of bacteriophage host range. FEMS Microbiol. Rev. 2023, 47, fuad038. [Google Scholar] [CrossRef] [PubMed]
- Whittle, M.J.; Bilverstone, T.W.; Van Esveld, R.J.; Lücke, A.C.; Lister, M.M.; Kuehne, S.A.; Minton, N.P. A novel bacteriophage with broad host range against Clostridioides difficile ribotype 078 supports slpa as the likely phage receptor. Microbiol. Spectr. 2022, 10, e02295-21. [Google Scholar] [CrossRef] [PubMed]
- Suleman, M.; Clark, J.R.; Bull, S.; Jones, J.D. Ethical argument for establishing good manufacturing practice for phage therapy in the UK. J. Med. Ethics 2024, 10. [Google Scholar] [CrossRef] [PubMed]
- Pinto, G.; Shetty, S.A.; Zoetendal, E.G.; Gonçalves, R.F.; Pinheiro, A.C.; Almeida, C.; Azeredo, J.; Smidt, H. An in vitro fermentation model to study the impact of bacteriophages targeting Shiga toxin-encoding Escherichia coli on the colonic microbiota. NPJ Biofilms Microbiomes 2022, 8, 74. [Google Scholar] [CrossRef]
- Dlamini, S.B.; Gigante, A.M.; Hooton, S.P.T.; Atterbury, R.J. Efficacy of different encapsulation techniques on the viability and stability of diverse phage under simulated gastric conditions. Microorganisms 2023, 11, 2389. [Google Scholar] [CrossRef]
- Abdelsattar, A.S.; Abdelrahman, F.; Dawoud, A.; Connerton, I.F.; El-Shibiny, A. Encapsulation of E. coli phage ZCEC5 in chitosan–alginate beads as a delivery system in phage therapy. AMB Express 2019, 9, 87. [Google Scholar] [CrossRef]
- Chen, C.; Tao, Z.; Li, T.; Chen, H.; Zhao, Y.; Sun, X. Isolation and characterization of novel bacteriophage vB_KpP_HS106 for Klebsiella pneumonia K2 and applications in foods. Front. Microbiol. 2023, 14, 1227147. [Google Scholar] [CrossRef]
- Harrell, J.E.; Hahn, M.M.; D’Souza, S.J.; Vasicek, E.M.; Sandala, J.L.; Gunn, J.S.; McLachlan, J.B. Salmonella biofilm formation, chronic infection, and immunity within the intestine and hepatobiliary tract. Front. Cell. Infect. Microbiol. 2021, 10, 624622. [Google Scholar]
- Fernández, L.; Gutiérrez, D.; García, P.; Rodríguez, A. The perfect bacteriophage for therapeutic applications—A quick guide. Antibiotics 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide | Sequence | Gene | Fragment Size (bp) |
|---|---|---|---|
| AvrA-F | CCTGTATTGTTGAGCGTCTGG | avrA | 422 |
| AvrA-R | AGAAGAGCTTCGTTGAATGTCC | ||
| SopB-F | TCAGAAGRCGTCTAACCACTC | sopB | 517 |
| SopB-R | TACCGTCCTCATGCACACTC | ||
| CdtB-F | GAAACAAGTCAGGCATTGCC | cdtB | 819 |
| CdtB-R | GAATGGCTCATAAACACGCC | ||
| PltA-F | GTGGGACTATCATCGTGCAG | pltA | 729 |
| PltA-R | AGGGTGATCAACGTAACCAC | ||
| PltB-F | GCCGGAAGTACCTGTGTTAT | pltB | 414 |
| PltB-R | AGTAGTGAAAACCCATCGCG |
| Reagent | Oral Phase (mmol/L) | Gastric Phase (mmol/L) | Intestinal Phase (mmol/L) |
|---|---|---|---|
| KCl | 15.1 | 6.9 | 6.8 |
| KH2PO4 | 3.7 | 0.9 | 0.8 |
| NaHCO3 | 13.6 | 25 | 85 |
| NaCl | - | 47.2 | 38.4 |
| MgCl2(H2O)6 | 0.15 | 0.12 | 0.33 |
| (NH4)2CO3 | 0.06 | 0.5 | - |
| CaCl2(H2O)2 | 1.5 | 0.15 | 0.6 |
| Enzymes | |||
| α-amylase | 150 U/mL | - | - |
| Pepsin | - | 4000 U/mL | - |
| Lipase | - | 120 U/mL | - |
| Pancreatin | - | - | 200 U/mL (based on trypsin activity) |
| Bile salts | - | - | 10 mM |
| Antimicrobials and Average Inhibition Diameter (mm) | Virulence Genes Associated with the Potential to Induce Precancerous Lesions | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacterial Strain | IMP | SXT | GEN | AMP | CIP | NAL | CHL | TET | COL | AMC | CFP | AMK | avrA | sopB | cdtB | pltA | pltB |
| Salmonella sal01 | 26 | 21 | 16 | 15 | 16 | 14 | 24 | 19 | 8 | 21 | 24 | 17 | - | + | + | - | + |
| Salmonella sal02 | 26 | 24 | 18 | 19 | 25 | 21 | 25 | 22 | 9 | 10 | 24 | 18 | + | + | + | + | + |
| Salmonella sal03 | 22 | 18 | 19 | 16 | 22 | 21 | 17 | 19 | 8 | 17 | 22 | 19 | + | - | + | + | + |
| Salmonella sal04 | 25 | 24 | 18 | 14 | 22 | 17 | 22 | 18 | 7 | 18 | 21 | 18 | + | - | + | - | + |
| Salmonella sal05 | 29 | 26 | 21 | 22 | 31 | 21 | 23 | 20 | 8 | 22 | 24 | 19 | + | + | + | + | + |
| Salmonella sal06 | 26 | 22 | 18 | 21 | 30 | 21 | 23 | 20 | 9 | 22 | 17 | 17 | + | - | - | - | + |
| Salmonella sal07 | 27 | 25 | 18 | 21 | 31 | 21 | 31 | 19 | 20 | 17 | 8 | 16 | + | + | + | + | + |
| Salmonella sal08 | 15 | 18 | 17 | 15 | 14 | 12 | 21 | 27 | 8 | 22 | 19 | 19 | + | - | + | + | + |
| Salmonella sal09 | 29 | 25 | 23 | 31 | 22 | 24 | 29 | 9 | 24 | 19 | 25 | 19 | + | + | + | + | + |
| Salmonella sal10 | 26 | 24 | 18 | 23 | 30 | 22 | 26 | 22 | 9 | 22 | 24 | 19 | + | + | + | - | - |
| Salmonella sal11 | 26 | 25 | 16 | 21 | 29 | 20 | 24 | 19 | 9 | 22 | 26 | 19 | + | + | + | + | + |
| Salmonella sal12 | 26 | 24 | 18 | 21 | 25 | 21 | 25 | 22 | 9 | 23 | 24 | 18 | + | + | + | + | + |
| Salmonella sal13 | 22 | 22 | 19 | 21 | 30 | 21 | 24 | 19 | 9 | 21 | 22 | 19 | + | - | + | - | + |
| Salmonella sal14 | 25 | 23 | 18 | 22 | 32 | 22 | 22 | 18 | 9 | 22 | 28 | 18 | + | - | - | + | + |
| Salmonella sal15 | 29 | 20 | 21 | 22 | 31 | 21 | 23 | 20 | 8 | 22 | 24 | 19 | + | + | + | + | + |
| Salmonella sal16 | 26 | 24 | 18 | 21 | 30 | 21 | 23 | 20 | 9 | 22 | 23 | 17 | + | + | + | + | - |
| Salmonella sal17 | 21 | 19 | 12 | 21 | 22 | 21 | 31 | 21 | 25 | 18 | 8 | 22 | + | - | + | + | + |
| Salmonella sal19 | 25 | 26 | 17 | 22 | 32 | 20 | 21 | 27 | 8 | 22 | 23 | 19 | + | + | + | + | + |
| Salmonella sal20 | 22 | 18 | 16 | 21 | 19 | 20 | 18 | 19 | 10 | 18 | 26 | 20 | + | + | + | + | + |
| Salmonella sal21 | 26 | 24 | 18 | 21 | 25 | 21 | 24 | 22 | 9 | 23 | 24 | 18 | + | + | + | + | + |
| Salmonella sal21 | 22 | 24 | 19 | 21 | 30 | 21 | 22 | 19 | 9 | 21 | 22 | 19 | + | - | + | + | - |
| Salmonella sal23 | 25 | 24 | 18 | 22 | 32 | 22 | 22 | 18 | 9 | 22 | 28 | 18 | + | + | + | + | + |
| Salmonella sal24 | 29 | 26 | 21 | 22 | 31 | 21 | 23 | 20 | 8 | 22 | 24 | 19 | + | - | + | + | + |
| Salmonella sal25 | 26 | 24 | 18 | 21 | 30 | 21 | 23 | 20 | 9 | 22 | 23 | 17 | + | + | + | + | + |
| Salmonella sal26 | 21 | 19 | 18 | 21 | 15 | 21 | 31 | 21 | 15 | 18 | 8 | 21 | + | - | + | + | + |
| Salmonella sal27 | 22 | 26 | 17 | 18 | 17 | 12 | 21 | 27 | 8 | 22 | 23 | 19 | + | - | + | + | + |
| Salmonella sal28 | 12 | 14 | 13 | 14 | 9 | 12 | 20 | 9 | 8 | 10 | 14 | 13 | + | + | + | + | + |
| Salmonella sal29 | 26 | 24 | 18 | 23 | 30 | 22 | 26 | 22 | 9 | 22 | 24 | 19 | + | - | + | - | + |
| Salmonella sal30 | 26 | 25 | 16 | 21 | 29 | 20 | 24 | 19 | 9 | 22 | 26 | 18 | + | + | + | + | + |
| Salmonella sal31 | 19 | 20 | 18 | 17 | 14 | 19 | 17 | 22 | 9 | 19 | 16 | 18 | + | + | + | + | + |
| Salmonella sal32 | 18 | 16 | 17 | 18 | 18 | 16 | 20 | 19 | 9 | 19 | 22 | 19 | + | - | + | + | + |
| Salmonella sal33 | 24 | 24 | 18 | 22 | 32 | 22 | 22 | 18 | 9 | 22 | 28 | 18 | + | - | + | + | + |
| Salmonella sal34 | 26 | 22 | 21 | 22 | 31 | 21 | 23 | 20 | 8 | 22 | 24 | 19 | + | + | + | + | + |
| Salmonella sal35 | 26 | 24 | 18 | 21 | 30 | 21 | 23 | 20 | 9 | 22 | 23 | 17 | + | + | + | - | + |
| Salmonella sal36 | 21 | 25 | 18 | 21 | 31 | 21 | 31 | 21 | 25 | 18 | 8 | 22 | + | + | + | + | + |
| Salmonella sal37 | 25 | 18 | 17 | 22 | 32 | 14 | 18 | 16 | 8 | 22 | 23 | 12 | + | + | + | - | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amarillas, L.; Padilla-Lafarga, F.; León Chan, R.G.; Padilla, J.; Lugo-Melchor, Y.; López Avendaño, J.E.; Lightbourn-Rojas, L.; Estrada-Acosta, M. Isolation and Characterization of a Bacteriophage with Potential for the Control of Multidrug-Resistant Salmonella Strains Encoding Virulence Factors Associated with the Promotion of Precancerous Lesions. Viruses 2024, 16, 1711. https://doi.org/10.3390/v16111711
Amarillas L, Padilla-Lafarga F, León Chan RG, Padilla J, Lugo-Melchor Y, López Avendaño JE, Lightbourn-Rojas L, Estrada-Acosta M. Isolation and Characterization of a Bacteriophage with Potential for the Control of Multidrug-Resistant Salmonella Strains Encoding Virulence Factors Associated with the Promotion of Precancerous Lesions. Viruses. 2024; 16(11):1711. https://doi.org/10.3390/v16111711
Chicago/Turabian StyleAmarillas, Luis, Fedra Padilla-Lafarga, Rubén Gerardo León Chan, Jorge Padilla, Yadira Lugo-Melchor, Jesús Enrique López Avendaño, Luis Lightbourn-Rojas, and Mitzi Estrada-Acosta. 2024. "Isolation and Characterization of a Bacteriophage with Potential for the Control of Multidrug-Resistant Salmonella Strains Encoding Virulence Factors Associated with the Promotion of Precancerous Lesions" Viruses 16, no. 11: 1711. https://doi.org/10.3390/v16111711
APA StyleAmarillas, L., Padilla-Lafarga, F., León Chan, R. G., Padilla, J., Lugo-Melchor, Y., López Avendaño, J. E., Lightbourn-Rojas, L., & Estrada-Acosta, M. (2024). Isolation and Characterization of a Bacteriophage with Potential for the Control of Multidrug-Resistant Salmonella Strains Encoding Virulence Factors Associated with the Promotion of Precancerous Lesions. Viruses, 16(11), 1711. https://doi.org/10.3390/v16111711