

Nipah Virus: A Multidimensional Update


Abstract
:1. Introduction
2. Nipah Virus Outbreaks
2.1. Malaysia-Singapore 1998–1999
2.2. Bangladesh
2.3. India
2.4. Philippines
3. Nipah Virus Characterization
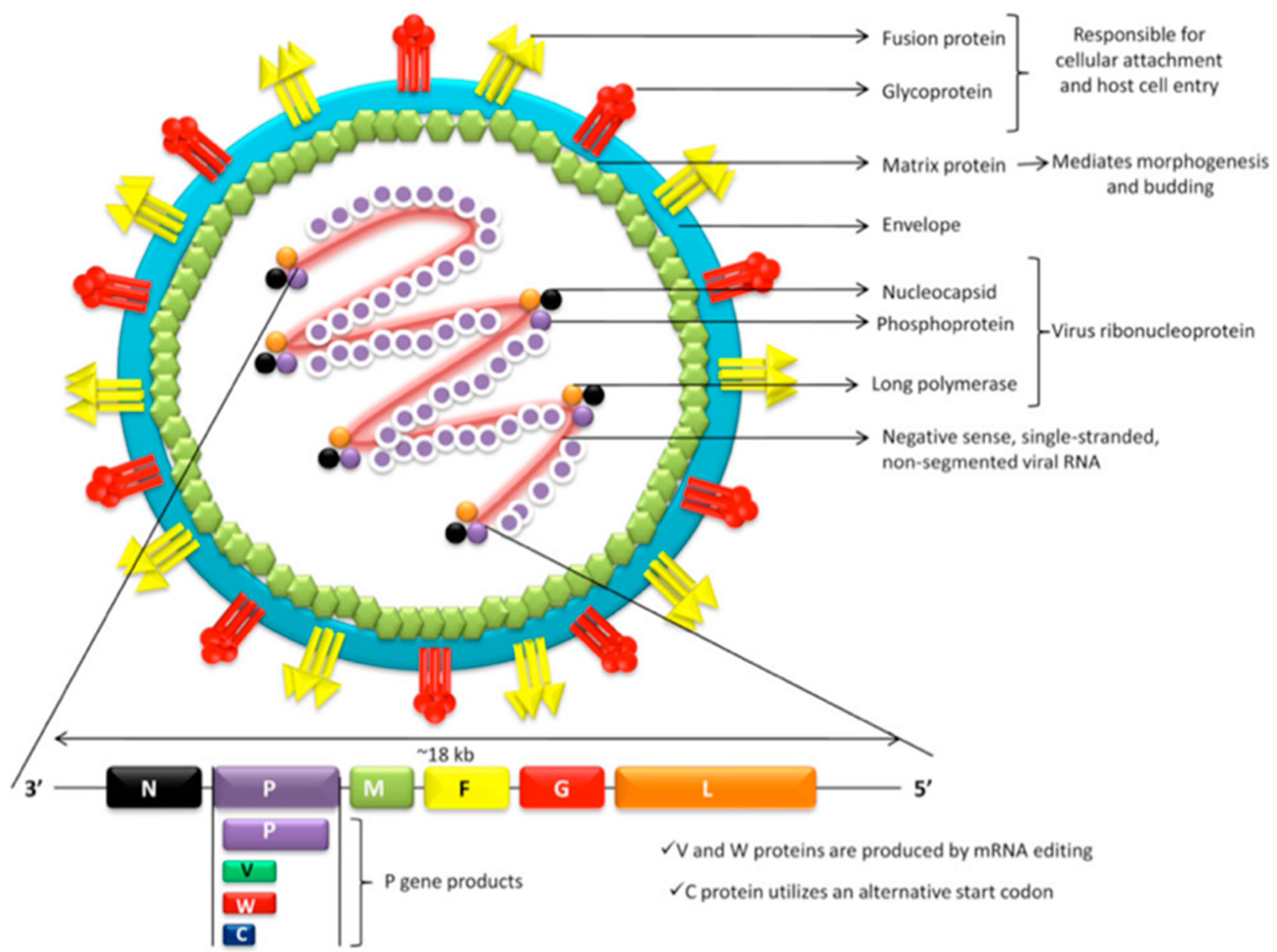
3.1. Viral Structure
3.2. Phylogeny and Viral Strains
4. Nipah Virus Reservoir and Host Range
4.1. Natural Reservoir
4.2. Intermediate Amplifier Hosts
5. Nipah Virus Infection
5.1. Pathogeny
5.2. Viral Immune Evasion
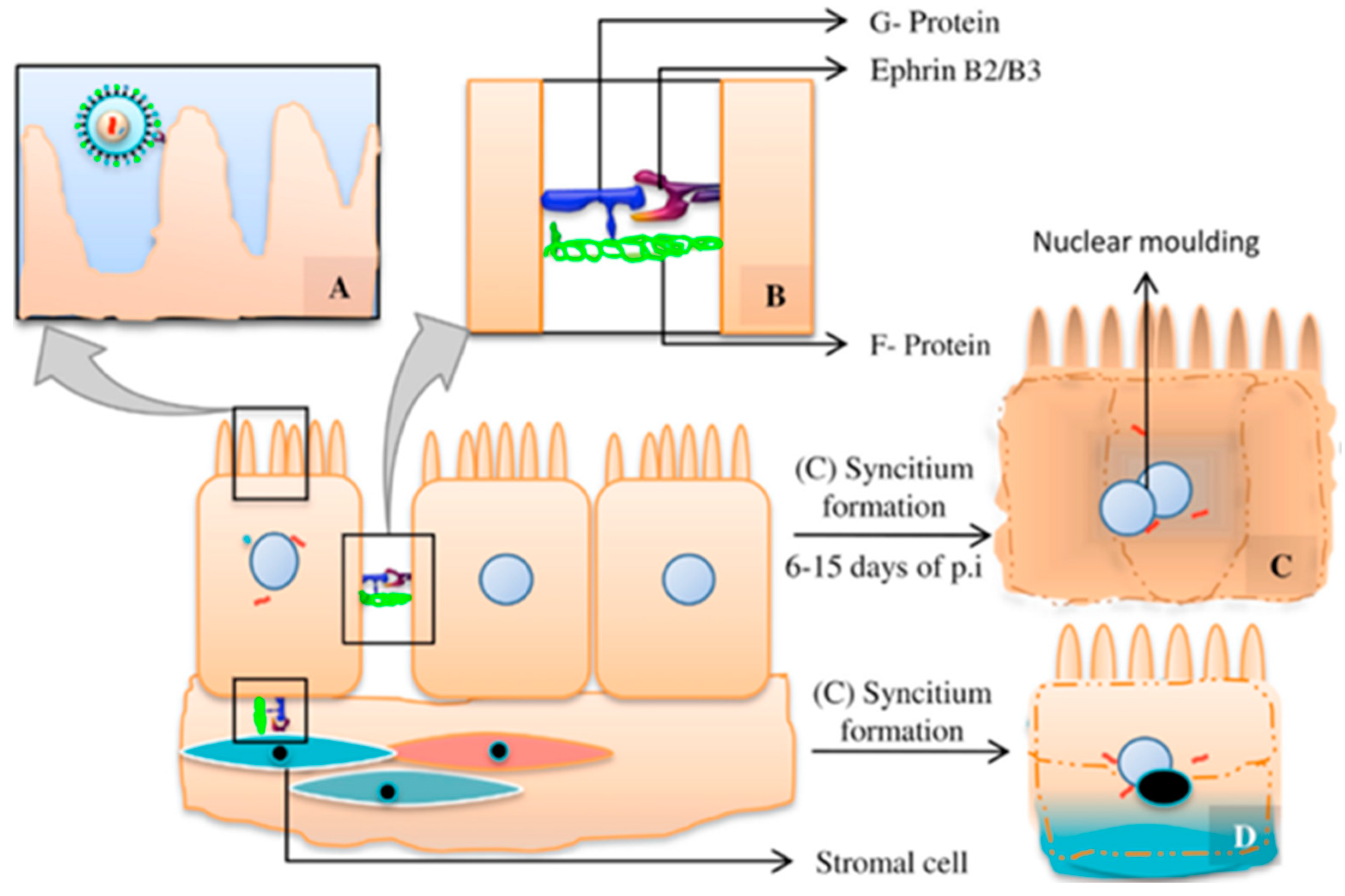
5.3. Viral Tropism and Tissular Lesion
5.4. Clinical Presentation
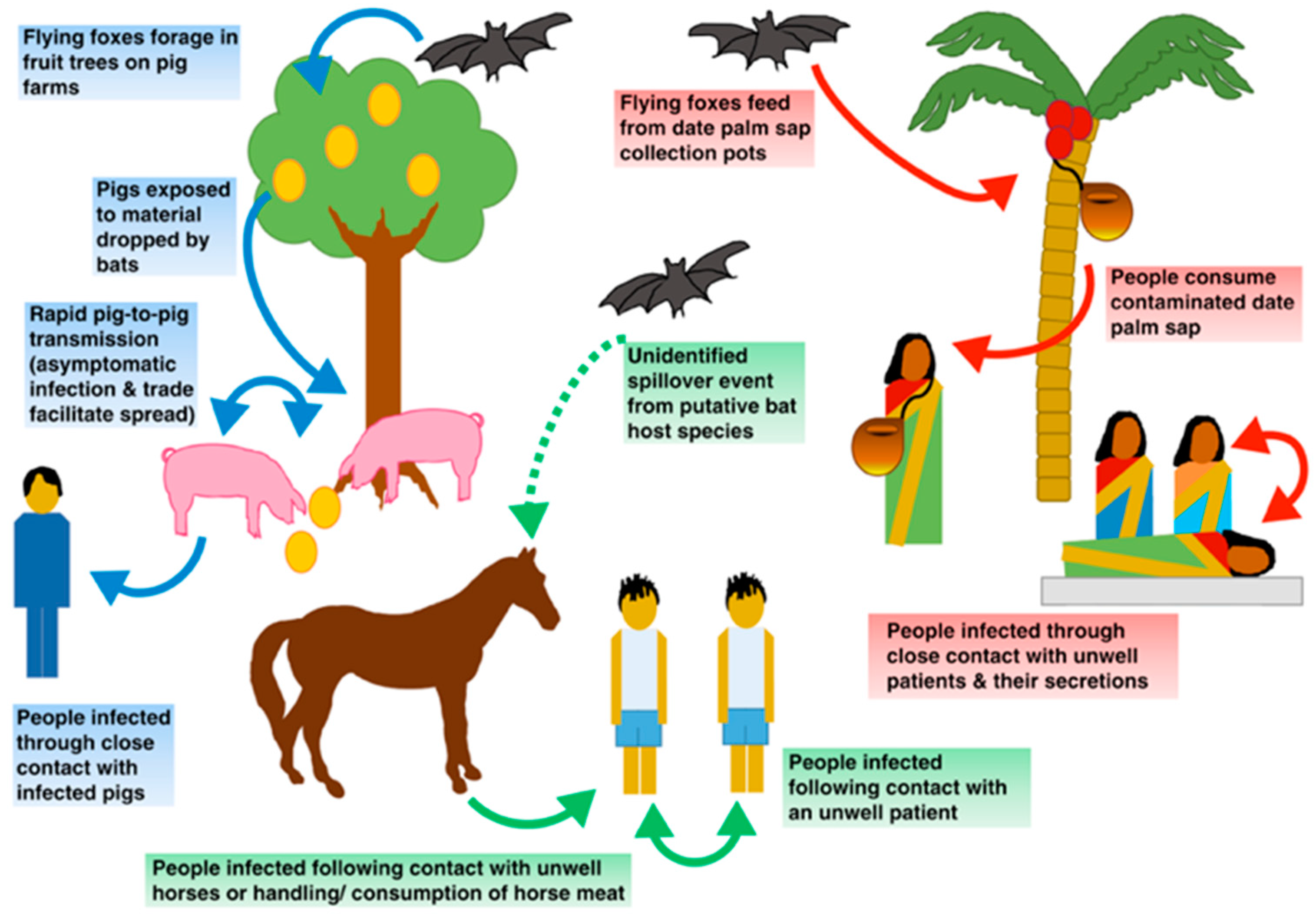
6. Nipah Virus Transmission
6.1. Interspecies Transmission

6.2. Human-to-Human Transmission
7. Countermeasure Requirements, Epidemic/Pandemic Potential of Nipah Virus and Public Health Implications
7.1. Implications of Bangladesh’s Outbreaks
7.2. Large-Scale Implications
- I exclusive to animals;
- II primary human infections only;
- III limited interhuman transmission;
- IV sustained interhuman transmission.
8. Therapeutic Agents
8.1. Nucleoside/Nucleotide Analogs
- Remdesivir: broad-spectrum RNA polymerase inhibitor antiviral prodrug. It has demonstrated potent replication-inhibitory activity in in vitro and in vivo experimentation. It constitutes a potential therapeutic candidate and a possible viral clearance adjuvant for survivors as a recurrence preventive measure. High seropositive maintained titers relate to incomplete viral clearance in patients and increased risk of infection recurrence; the intense viral clearance observed in experimental animals treated with remdesivir supports its use on selected patients [40].
- Favipiravir: selective RNA polymerase inhibitor antiviral drug. Great replication- and transcription-inhibitory capacity observed in vitro even with low drug concentrations. Experimental animal models with Golden Hamster reveal complete protection against lethal NiV doses. Considered as a highly potential human therapeutic agent as well as a post-exposure prophylaxis candidate [39].
- Balapiravir: it has demonstrated in vitro efficacy against NiV, currently pending in vivo experimentation [39].
8.2. Monoclonal Antibodies
- m102.4: a highly potent and promising human monoclonal antibody directed to the G viral glycoprotein–Ephrin B2/B3 interaction surface. For its synthesis, it has been selected by the screening of G-soluble glycoprotein form antibody library and exposed to a posterior affinity maturation by light chain reconstruction and random mutation of heavy chain variable regions [40,41]. It exhibits extensive neutralizing in vitro activity against all tested NiV and HeV strains. In vivo efficacy against NiV lethal doses has been confirmed in African Green Monkey and ferret experimental animal models in therapeutic conditions within several post-viral inoculation drug administration intervals (including after the initiation of clinical signs and circulating virus detection). It maintains an appropriate in vivo stability, and its biological activity extends until eight days post-infusion in animal models. Since 2010, m102.4 has been employed in humans for compassionate use for post-exposure treatment against NiV and HeV in USA, India, and Australia; however, real human efficacy remains unknown [6,39,42].
- h5B3.1: a humanized monoclonal antibody directed to the prefusion conformation of F viral glycoprotein; able to prevent virus–host cell membrane attachment and viral penetration by junction with quaternary F glycoprotein epitopes. Considered as a potential candidate for post-exposure treatment/prophylaxis. Currently pending in vivo experimentation [42].
- nAH1.3: a broadly neutralizing antibody interfering with the F fusion-triggering mechanism and a potency comparable to that of m102.4. They both have distinct antigenic sites that are non-competitive and could be a potential treatment option in combination [43].
8.3. Fusion-Inhibitory Peptides
- Cholesterol-tagged fusion-inhibitory peptides: designed to block F glycoprotein conformational changes leading to viral pore-mediated host cell penetration. Cholesterol tag provides a drastic increase in antiviral efficacy by directing peptides to target host cell membrane. Golden Hamster experimental models reveal an 80% survival rate with lethal viral doses when administered at the same time as viral inoculation, showing elevated penetration and high drug concentration in CNS, lung, and vascular endothelium. A remarkable decrease in survival rate (40%) has been observed when infusion is delayed by 48 h [45].
- Inhaled fusion-inhibitory lipopeptides: as an advantage, the inhaled administration route allows focused respiratory system delivery, covering the main viral entry pathway. In vivo experimentation has been conducted in Golden Hamster and African Green Monkey, a 33% relative mortality risk reduction was observed for the African Green Monkey model [40,46].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Efficacy | Endemic Region Availability | Safety | Advantages |
|---|---|---|---|---|
| Remdesivir |
|
|
|
|
| Favipiravir |
|
|
|
|
| m102.4 |
|
|
|
|
| Fusion-inhibitory peptides/lipopeptides |
|
|
|
|
9. Preventive and Outbreak Control Strategies
9.1. Surveillance and Outbreak Detection Strategies
- Populational surveillance systems: several strategies have been implemented previously in Bangladesh with this aim, including Nipah belt-focused hospital surveillance (plus Nipah season intensification), 24 h hotline implementation for adverse health events notification, and mass media information monitoring [21,47]. Mass media monitoring was implemented from 2010–2011 by using Bangladesh’s National Media-Based Public Surveillance System, and it represented a highly effective, low-cost, sustainable outbreak detection strategy, which suits low-income countries with scarce health infrastructure development, such as Bangladesh. It is based on the extensive monitoring of the principal media information sources for the early detection and investigation of possible NiV outbreaks [47].
- Exposure-based screening: the use of brief hospital admission questionnaires to assess patients’ previous risk exposure was demonstrated to be highly effective and efficient when implemented in Bangladesh during 2012–2013 for early NiV-encephalitis case detection and interhuman transmission prevention. Asking for sap consumption and history of contact with febrile patients with altered cognition during the previous 30 days since symptom onset proved to be a useful NiV screening tool, especially during Nipah season (observed negative predictive value NPV = 99% during wintertime). In addition, screening questionnaire implementation allows a more efficient use of NiV transmission prevention resources in hospital settings, which are frequently scarce [36].
9.2. Human Infection and Transmission Preventive Interventions
| Strategy | Efficacy | Effectiveness | Cost Efficiency | Endemic Region Availability | Comments |
|---|---|---|---|---|---|
| Populational surveillance systems |
|
|
|
|
|
| Exposure-based screening |
|
|
|
|
|
| Populational/community educational intervention |
|
|
|
|
|
| Physical barrier bamboo skit method |
|
|
|
|
|
- Populational/community educational intervention: a multilevel information campaign adjusted for specific risk factors in a targeted population (i.e., fresh sap consumption in rural areas, direct contact transmission in hospital settings, etc.) (Table 3). This strategy makes the most of educative measures to impact the prevention of infectious diseases [21]. During 2009, several messages were disseminated in rural Bangladesh to dissuade the population from sap consumption; the “only safe sap” campaign represented the best approach, achieving a great improvement in disease knowledge and transmission awareness. The message is a harm reduction approach that recognizes abstinence as an ideal outcome but accepts alternatives that reduce harm (“Only safe sap” refers to consumption of physical barrier harvested palm sap) [31].
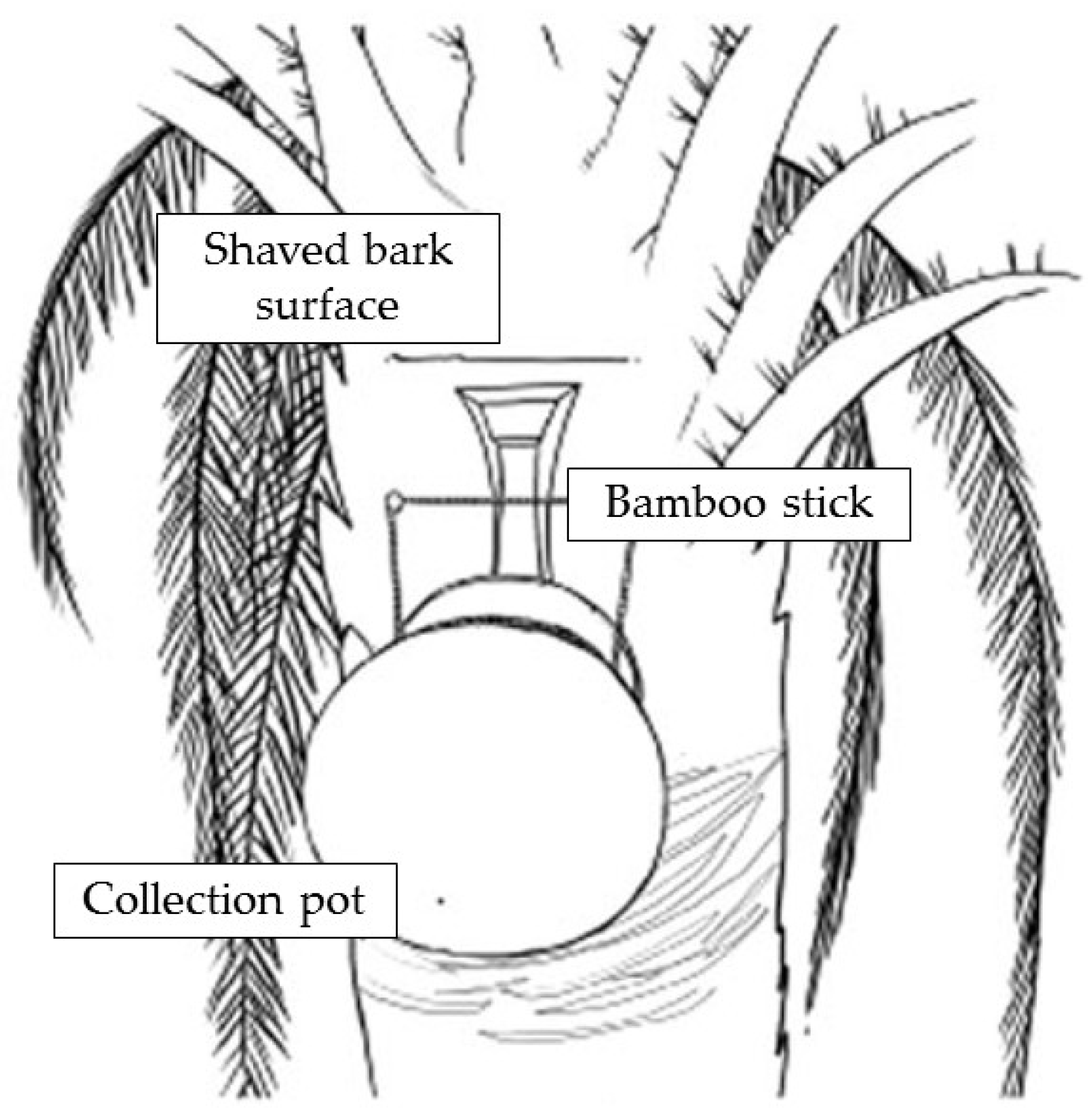
- Physical barrier strategies for bat-to-human transmission prevention: the fresh date palm sap harvesting period overlaps the Nipah outbreak season as contaminated sap consumption is the main route for NiV emergence in Bangladesh. Local harvesters, known as “gachhis”, collect and sell fresh sap during the early morning, and consumers typically ingest the fresh sap within the same day. Under these conditions, viable viral load reduction in contaminated sap is minimal; hence, interventions that prevent sap contamination from Pteropus spp. Stand out as valuable strategies [21,31,51]. Traditionally “gachhis” have used several methods to prevent sap deterioration from bat urine and excrements, including the use of tree branches to cover the sap circulation area, bark impregnation with lime, and the placing of bamboo skirts. Infrared camera evidence has demonstrated that only bamboo skirts, locally known as “banas”, are capable of preventing bat–sap flow physical contact [35,51]. Camera evidence proves Pteropus spp. frequently visit date palm trees and contaminate the collecting sap flow by directly contacting and feeding from it (Figure 5). Although further experimentation is needed to assess whether the “bana” method offers large-scale effectiveness, it still remains as a high potential preventive strategy. Zoonotic transmission prevention is considered the most efficient and cost-effective strategy to prevent human outbreaks. Furthermore, it is a low-cost intervention, easy to install, and accepted by “gachhis” when asked, which allows the conservation of the traditional practice of fresh sap harvesting and consumption [13,35,51].
9.3. Vaccination
- Subunit vaccines: epitopes or viral peptides, highly specific, easy to produce low-cost vaccines. Bioinformatics tools predict the main epitopes are able to trigger a sufficient immune response; F and G glycoprotein fractions stand out as the best candidates [15]. Animal experimentation in African Green Monkey models reveals complete protection and development of high IgG levels against NiV with a subunit vaccine based on the oligomeric soluble form of G recombinant HeV glycoprotein (sGHeV). The elevated immunogenicity observed and its exceptional efficacy support future evaluation and, eventually, authorization for human use [5,9].
- Vector vaccines: attenuated virus able to express G/F NiV recombinant glycoproteins. Promising results have been observed in in vivo experimentation with pig and Golden Hamster animal models, showing adequate serological response with several viral vectors (canarypox virus, vesicular stomatitis virus, and Venezuelan equine encephalitis virus). Virus-like particles derived from mammal cells expressing F, G, and M viral proteins have also demonstrated the ability to induce a potent neutralizing response and complete protection against lethal doses of NiV in Golden Hamster models [9].
- mRNA-1215 vaccine: a candidate mRNA vaccine under evaluation in a Phase 1 clinical trial has been developed by NIAID and Moderna. It encodes for the prefusion state of the F protein covalently linked to the G protein monomer (pre-F/G) of the Malaysian strain NiV [52].
10. Comparison of Strategies Focused on Endemic Regions
11. Future Directions
- Bangladesh, and specifically the Nipah belt, should be prioritized as the main target of anti-Nipah interventions because of the frequency and regularity of viral emergences. Human outbreaks in other Asiatic regions have depended on the presence of intermediate amplification hosts which facilitated the transmission to humans. For this to occur, the harmonization of several conditions were required, including the presence of a bat-to-amplification mammal interface and a subsequent amplification mammal-to-human interface. This defines an extraordinarily tough to predict temporo-spatial intersection, owing to the vast intercontinental geographic distribution of NiV-disseminating Pteropus spp. bats. Because of this, the rare viral emergences that may occur in other countries are very difficult to prevent. In contrast, Bangladesh’s frequent outbreaks are tightly linked to a specific risk exposure (date palm sap consumption); thus, an evident bat-to-human nexus determining the viral transmission is established. This also provides an obvious source of outbreak prevention by blocking bat-to-human transmission, which would minimize both regional and global outbreak-derived potential adversities.
- Further research and development of therapeutic agents is required due to the urgent need of treatment implementation in order to confront the extremely high mortality and morbidity of NiV human infection in endemic regions. A number of drugs and monoclonal antibodies have been developed and require future research in animal models and clinical trials for an eventual authorization for human use.
- Current economic investment in endemic regions should focus on low-cost, effective, and efficient preventive strategies able to interrupt interspecies transmission in the Nipah belt. The recommendations of the WHO and CEPI to assign priority to the development of therapeutic and preventive tools against NiV should serve to enhance the development and future application of effective measures in the countries with the greatest impact of the disease.
- A pragmatic consideration of the real epidemic/pandemic risk of novel emerging viruses is required, which relates to the need of infection contention assistance by the international community to certain regions with special risk of novel pathogen emergence and dissemination. Furthermore, as has been demonstrated with the COVID-19 pandemic, surveillance for potential pandemic agents, such as NiV, is essential for the implementation of early measures in local outbreaks or in broader circulation of the virus.
- The persistent human–wildlife interaction derived from environment modification by human activities will certainly promote the emergence of novel pathogens in the future, hence exposing large human populations to unpredictable threats.
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pulliam, J.R.; Epstein, J.H.; Dushoff, J.; Rahman, S.A.; Bunning, M.; Jamaluddin, A.A.; Hyatt, A.D.; Field, H.E.; Dobson, A.P.; Daszak, P. Agricultural intensification, priming for persistence and the emergence of Nipah virus: A lethal bat-borne zoonosis. J. R. Soc. Interface 2012, 9, 89–101. [Google Scholar] [CrossRef]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proceedings. Biol. Sci. 2015, 282, 2014–2124. [Google Scholar] [CrossRef] [PubMed]
- Angeletti, S.; Lo Presti, A.; Cella, E.; Ciccozzi, M. Molecular epidemiology and phylogeny of Nipah virus infection: A mini review. Asian Pac. J. Trop. Med. 2016, 9, 630–634. [Google Scholar] [CrossRef] [PubMed]
- Clayton, B.A. Nipah virus: Transmission of a zoonotic paramyxovirus. Curr. Opin. Virol. 2017, 22, 97–104. [Google Scholar] [CrossRef]
- Bossart, K.N.; Rockx, B.; Feldmann, F.; Brining, D.; Scott, D.; LaCasse, R.; Geisbert, J.B.; Feng, Y.R.; Chan, Y.P.; Hickey, A.C.; et al. A Hendra virus G glycoprotein subunit vaccine protects African green monkeys from Nipah virus challenge. Sci. Transl. Med. 2012, 4, 146ra107. [Google Scholar] [CrossRef] [PubMed]
- Geisbert, T.W.; Mire, C.E.; Geisbert, J.B.; Chan, Y.P.; Agans, K.N.; Feldmann, F.; Fenton, K.A.; Zhu, Z.; Dimitrov, D.S.; Scott, D.P.; et al. Therapeutic treatment of Nipah virus infection in nonhuman primates with a neutralizing human monoclonal antibody. Sci. Transl. Med. 2014, 6, 242–258. [Google Scholar] [CrossRef]
- Lo, M.K.; Rota, P.A. The emergence of Nipah virus, a highly pathogenic paramyxovirus. J. Clin. Virol. 2008, 43, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.S.; Sazzad, H.M.; Satter, S.M.; Sultana, S.; Hossain, M.J.; Hasan, M.; Rahman, M.; Campbell, S.; Cannon, D.L.; Ströher, U.; et al. Nipah Virus Transmission from Bats to Humans Associated with Drinking Traditional Liquor Made from Date Palm Sap, Bangladesh, 2011–2014. Emerg. Infect. Dis. 2016, 22, 664–670. [Google Scholar] [CrossRef]
- Thakur, N.; Bailey, D. Advances in diagnostics, vaccines and therapeutics for Nipah virus. Microbes Infect. 2019, 21, 278–286. [Google Scholar] [CrossRef]
- Cortes, M.C.; Cauchemez, S.; Lefrancq, N.; Luby, S.P.; Jahangir Hossain, M.; Sazzad, H.M.S.; Rahman, M.; Daszak, P.; Salje, H.; Gurley, E.S. Characterization of the Spatial and Temporal Distribution of Nipah Virus Spillover Events in Bangladesh, 2007–2013. J. Infect. Dis. 2018, 217, 1390–1394. [Google Scholar] [CrossRef]
- Spiropoulou, C.F. Nipah Virus Outbreaks: Still Small but Extremely Lethal. J. Infect. Dis. 2019, 219, 1855–1857. [Google Scholar] [CrossRef]
- Plowright, R.K.; Becker, D.J.; Crowley, D.E.; Washburne, A.D.; Huang, T.; Nameer, P.O.; Gurley, E.S.; Han, B.A. Prioritizing surveillance of Nipah virus in India. PLoS Neglected Trop. Dis. 2019, 13, e0007393. [Google Scholar] [CrossRef]
- Weatherman, S.; Feldmann, H.; de Wit, E. Transmission of henipaviruses. Curr. Opin. Virol. 2018, 28, 7–11. [Google Scholar] [CrossRef]
- Kerry, R.G.; Malik, S.; Redda, Y.T.; Sahoo, S.; Patra, J.K.; Majhi, S. Nano-based approach to combat emerging viral (NIPAH virus) infection. Nanomedicine 2019, 18, 196–220. [Google Scholar] [CrossRef]
- Kamthania, M.; Sharma, D.K. Screening and structure-based modeling of T-cell epitopes of Nipah virus proteome: An immunoinformatic approach for designing peptide-based vaccine. 3 Biotech 2015, 5, 877–882. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, M.; Guillaume, V.; Sato, H.; Fujita, K.; Georges-Courbot, M.C.; Ikeda, F.; Omi, M.; Muto-Terao, Y.; Wild, T.F.; Kai, C. The nonstructural proteins of Nipah virus play a key role in pathogenicity in experimentally infected animals. PLoS ONE 2010, 5, e12709. [Google Scholar] [CrossRef]
- Singh, R.K.; Dhama, K.; Chakraborty, S.; Tiwari, R.; Natesan, S.; Khandia, R.; Munjal, A.; Vora, K.S.; Latheef, S.K.; Karthik, K.; et al. Nipah virus: Epidemiology, pathology, immunobiology and advances in diagnosis, vaccine designing and control strategies—A comprehensive review. Vet. Q. 2019, 39, 26–55. [Google Scholar] [CrossRef] [PubMed]
- Luby, S.P. The pandemic potential of Nipah virus. Antivir. Res. 2013, 100, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Nikolay, B.; Salje, H.; Hossain, M.J.; Khan, A.K.M.D.; Sazzad, H.M.S.; Rahman, M.; Daszak, P.; Ströher, U.; Pulliam, J.R.C.; Kilpatrick, A.M.; et al. Transmission of Nipah Virus—14 Years of Investigations in Bangladesh. N. Engl. J. Med. 2019, 380, 1804–1814. [Google Scholar] [CrossRef]
- Hossain, M.J.; Gurley, E.S.; Montgomery, J.M.; Bell, M.; Carroll, D.S.; Hsu, V.P.; Formenty, P.; Croisier, A.; Bertherat, E.; Faiz, M.A.; et al. Clinical presentation of nipah virus infection in Bangladesh. Clin. Infect. Dis. 2008, 46, 977–984. [Google Scholar] [CrossRef]
- World Health Organization. National Guideline for Management, Prevention and Control of Nipah Virus Infection including Encephalitis. 2011. Available online: https://www.who.int/docs/default-source/searo/bangladesh/pdf-reports/year-2007-2012/national-guideline-for-management-prevention-and-control-of-nipah-virus-infection-including-encephalitis.pdf?sfvrsn=a5801092_2 (accessed on 30 December 2023).
- Rahman, S.A.; Hassan, S.S.; Olival, K.J.; Mohamed, M.; Chang, L.Y.; Hassan, L.; Saad, N.M.; Shohaimi, S.A.; Mamat, Z.C.; Naim, M.S.; et al. Characterization of Nipah virus from naturally infected Pteropus vampyrus bats, Malaysia. Emerg. Infect. Dis. 2010, 16, 1990–1993. [Google Scholar] [CrossRef]
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of Humans, Bats, Trees, and Culture in Nipah Virus Transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453. [Google Scholar] [CrossRef]
- Donaldson, H.; Lucey, D. Enhancing preparation for large Nipah outbreaks beyond Bangladesh: Preventing a tragedy like Ebola in West Africa. Int. J. Infect. Dis. 2018, 72, 69–72. [Google Scholar] [CrossRef]
- Wong, J.J.W.; Young, T.A.; Zhang, J.; Liu, S.; Leser, G.P.; Komives, E.A.; Lamb, R.A.; Zhou, Z.H.; Salafsky, J.; Jardetzky, T.S. Monomeric ephrinB2 binding induces allosteric changes in Nipah virus G that precede its full activation. Nat. Commun. 2017, 8, 781–791. [Google Scholar] [CrossRef]
- Vogt, C.; Eickmann, M.; Diederich, S.; Moll, M.; Maisner, A. Endocytosis of the Nipah virus glycoproteins. J. Virol. 2005, 79, 3865–3872. [Google Scholar] [CrossRef] [PubMed]
- Maisner, A.; Neufeld, J.; Weingartl, H. Organ- and endotheliotropism of Nipah virus infections in vivo and in vitro. Thromb. Haemost. 2009, 102, 1014–1023. [Google Scholar] [CrossRef]
- Dietzel, E.; Kolesnikova, L.; Sawatsky, B.; Heiner, A.; Weis, M.; Kobinger, G.P.; Becker, S.; von Messling, V.; Maisner, A. Nipah Virus Matrix Protein Influences Fusogenicity and Is Essential for Particle Infectivity and Stability. J. Virol. 2015, 90, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Bowden, T.A.; Aricescu, A.R.; Gilbert, R.J.; Grimes, J.M.; Jones, E.Y.; Stuart, D.I. Structural basis of Nipah and Hendra virus attachment to their cell-surface receptor ephrin-B2. Nat. Struct. Mol. Biol. 2008, 15, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Gurley, E.S.; Montgomery, J.M.; Hossain, M.J.; Bell, M.; Azad, A.K.; Islam, M.R.; Molla, M.A.; Carroll, D.S.; Ksiazek, T.G.; Rota, P.A.; et al. Person-to-person transmission of Nipah virus in a Bangladeshi community. Emerg. Infect. Dis. 2007, 13, 1031–1037. [Google Scholar] [CrossRef]
- Nahar, N.; Paul, R.C.; Sultana, R.; Sumon, S.A.; Banik, K.C.; Abedin, J.; Asaduzzaman, M.; Garcia, F.; Zimicki, S.; Rahman, M.; et al. A Controlled Trial to Reduce the Risk of Human Nipah Virus Exposure in Bangladesh. Ecohealth 2017, 14, 501–517. [Google Scholar] [CrossRef]
- Sejvar, J.J.; Hossain, J.; Saha, S.K.; Gurley, E.S.; Banu, S.; Hamadani, J.D.; Faiz, M.A.; Siddiqui, F.M.; Mohammad, Q.D.; Mollah, A.H.; et al. Long-term neurological and functional outcome in Nipah virus infection. Ann. Neurol. 2007, 62, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Luby, S.P.; Rahman, M.; Hossain, M.J.; Blum, L.S.; Husain, M.M.; Gurley, E.; Khan, R.; Ahmed, B.N.; Rahman, S.; Nahar, N.; et al. Foodborne transmission of Nipah virus, Bangladesh. Emerg. Infect. Dis. 2006, 12, 1888–1894. [Google Scholar] [CrossRef]
- Epstein, J.H.; Field, H.E.; Luby, S.; Pulliam, J.R.C.; Daszak, P. Nipah Virus: Impact, Origins, and Causes of Emergence. Curr. Infect. Dis. Rep. 2006, 8, 59–65. [Google Scholar] [CrossRef]
- Nahar, N.; Sultana, R.; Gurley, E.S.; Hossain, M.J.; Luby, S.P. Date palm sap collection: Exploring opportunities to prevent Nipah transmission. Ecohealth 2010, 7, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Sazzad, H.M.; Luby, S.P.; Ströher, U.; Daszak, P.; Sultana, S.; Afroj, S.; Rahman, M.; Gurley, E.S. Exposure-based screening for Nipah virus encephalitis, Bangladesh. Emerg. Infect. Dis. 2015, 21, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Reina González, G.; Toledo Atucha, E.; Carlos Chillerón, S. Enfermedades transmisibles: Aspectos generales. In Conceptos de Salud Pública y Estrategias Preventivas, 2nd ed.; Martínez-González, M.A., Ed.; Elsevier: Barcelona, Spain, 2018; pp. 171–176. [Google Scholar]
- Negrete, O.A.; Levroney, E.L.; Aguilar, H.C.; Bertolotti-Ciarlet, A.; Nazarian, R.; Tajyar, S.; Lee, B. EphrinB2 is the entry receptor for Nipah virus, an emergent deadly paramyxovirus. Nature 2005, 436, 401–4055. [Google Scholar] [CrossRef]
- Dawes, B.E.; Kalveram, B.; Ikegami, T.; Juelich, T.; Smith, J.K.; Zhang, L.; Park, A.; Lee, B.; Komeno, T.; Furuta, Y.; et al. Favipiravir (T-705) protects against Nipah virus infection in the hamster model. Sci. Rep. 2018, 8, 7604–7615. [Google Scholar] [CrossRef]
- Lo, M.K.; Feldmann, F.; Gary, J.M.; Jordan, R.; Bannister, R.; Cronin, J.; Patel, N.R.; Klena, J.D.; Nichol, S.T.; Cihlar, T.; et al. Remdesivir (GS-5734) protects African green monkeys from Nipah virus challenge. Sci. Transl. Med. 2019, 11, eaau9242. [Google Scholar] [CrossRef]
- Zhu, Z.; Bossart, K.N.; Bishop, K.A.; Crameri, G.; Dimitrov, A.S.; McEachern, J.A.; Feng, Y.; Middleton, D.; Wang, L.F.; Broder, C.C.; et al. Exceptionally potent cross-reactive neutralization of Nipah and Hendra viruses by a human monoclonal antibody. J. Infect. Dis. 2008, 197, 846–853. [Google Scholar] [CrossRef]
- Dang, H.V.; Chan, Y.P.; Park, Y.J.; Snijder, J.; Da Silva, S.C.; Vu, B.; Yan, L.; Feng, Y.R.; Rockx, B.; Geisbert, T.W.; et al. An antibody against the F glycoprotein inhibits Nipah and Hendra virus infections. Nat. Struct. Mol. Biol. 2019, 26, 980–987. [Google Scholar] [CrossRef]
- Wang, Z.; Amaya, M.; Addetia, A.; Dang, H.V.; Reggiano, G.; Yan, L.; Hickey, A.C.; DiMaio, F.; Broder, C.C.; Veesler, D. Architecture and antigenicity of the Nipah virus attachment glycoprotein. Science 2022, 375, 1373–1378. [Google Scholar] [CrossRef]
- Mishra, G.; Prajapat, V.; Nayak, D. Advancements in Nipah virus treatment: Analysis of current progress in vaccines, antivirals, and therapeutics. Immunology, 2023; online ahead of print. [Google Scholar] [CrossRef]
- Porotto, M.; Rockx, B.; Yokoyama, C.C.; Talekar, A.; Devito, I.; Palermo, L.M.; Liu, J.; Cortese, R.; Lu, M.; Feldmann, H.; et al. Inhibition of Nipah virus infection in vivo: Targeting an early stage of paramyxovirus fusion activation during viral entry. PLoS Pathog. 2010, 6, e1001168. [Google Scholar] [CrossRef]
- Mathieu, C.; Porotto, M.; Figueira, T.N.; Horvat, B.; Moscona, A. Fusion Inhibitory Lipopeptides Engineered for Prophylaxis of Nipah Virus in Primates. J. Infect. Dis. 2018, 218, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Ao, T.T.; Rahman, M.; Haque, F.; Chakraborty, A.; Hossain, M.J.; Haider, S.; Alamgir, A.S.; Sobel, J.; Luby, S.P.; Gurley, E.S. Low-Cost National Media-Based Surveillance System for Public Health Events, Bangladesh. Emerg. Infect. Dis. 2016, 22, 720–722. [Google Scholar] [CrossRef] [PubMed]
- Luby, S.P.; Gurley, E.S.; Hossain, M.J. Transmission of human infection with Nipah virus. Clin. Infect. Dis. 2009, 49, 1743–1748. [Google Scholar] [CrossRef]
- Chadha, M.S.; Comer, J.A.; Lowe, L.; Rota, P.A.; Rollin, P.E.; Bellini, W.J.; Ksiazek, T.G.; Mishra, A. Nipah virus-associated encephalitis outbreak, Siliguri, India. Emerg. Infect. Dis. 2006, 12, 235–240. [Google Scholar] [CrossRef]
- Montgomery, J.M.; Hossain, M.J.; Gurley, E.; Carroll, G.D.; Croisier, A.; Bertherat, E.; Asgari, N.; Formenty, P.; Keeler, N.; Comer, J.; et al. Risk factors for Nipah virus encephalitis in Bangladesh. Emerg. Infect. Dis. 2008, 14, 1526–1532. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Hossain, J.; Gurley, E.S.; Nahar, N.; Sultana, R.; Luby, S.P. Use of infrared camera to understand bats’ access to date palm sap: Implications for preventing Nipah virus transmission. Ecohealth 2010, 7, 517–525. [Google Scholar] [CrossRef]
- Rzymski, P.; Szuster-Ciesielska, A.; Dzieciątkowski, T.; Gwenzi, W.; Fal, A. mRNA vaccines: The future of prevention of viral infections? J. Med. Virol. 2023, 95, e28572. [Google Scholar] [CrossRef]
- Guillén-Grima, F.; Álvarez-Ruiz, J.C.; Núñez-Córdoba, J.; Pinto Prades, J.L. Evaluación económica y análisis de decisiones. In Conceptos de Salud Pública y Estrategias Preventivas, 2nd ed.; Martínez-González, M.A., Ed.; Elsevier: Barcelona, Spain, 2018; pp. 73–77. [Google Scholar]
- Núñez-Córdoba, J.M.; Rodríguez-Rieiro, C.; Díez-Espino, J. Gestión de la calidad, planificación y programación sanitaria. In Conceptos de Salud Pública y Estrategias Preventivas, 2nd ed.; Martínez-González, M.A., Ed.; Elsevier: Barcelona, Spain, 2018; pp. 339–342. [Google Scholar]




|
|
|
|
|
|
|
|
| Use personal protection equipment |
|
| Hand hygiene |
|
| Use disposable items |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faus-Cotino, J.; Reina, G.; Pueyo, J. Nipah Virus: A Multidimensional Update. Viruses 2024, 16, 179. https://doi.org/10.3390/v16020179
Faus-Cotino J, Reina G, Pueyo J. Nipah Virus: A Multidimensional Update. Viruses. 2024; 16(2):179. https://doi.org/10.3390/v16020179
Chicago/Turabian StyleFaus-Cotino, Javier, Gabriel Reina, and Javier Pueyo. 2024. "Nipah Virus: A Multidimensional Update" Viruses 16, no. 2: 179. https://doi.org/10.3390/v16020179
APA StyleFaus-Cotino, J., Reina, G., & Pueyo, J. (2024). Nipah Virus: A Multidimensional Update. Viruses, 16(2), 179. https://doi.org/10.3390/v16020179