
Metformin Alters mRNA Expression of FOXP3, RORC, and TBX21 and Modulates Gut Microbiota in COVID-19 Patients with Type 2 Diabetes





Abstract
1. Introduction
2. Materials and Methods
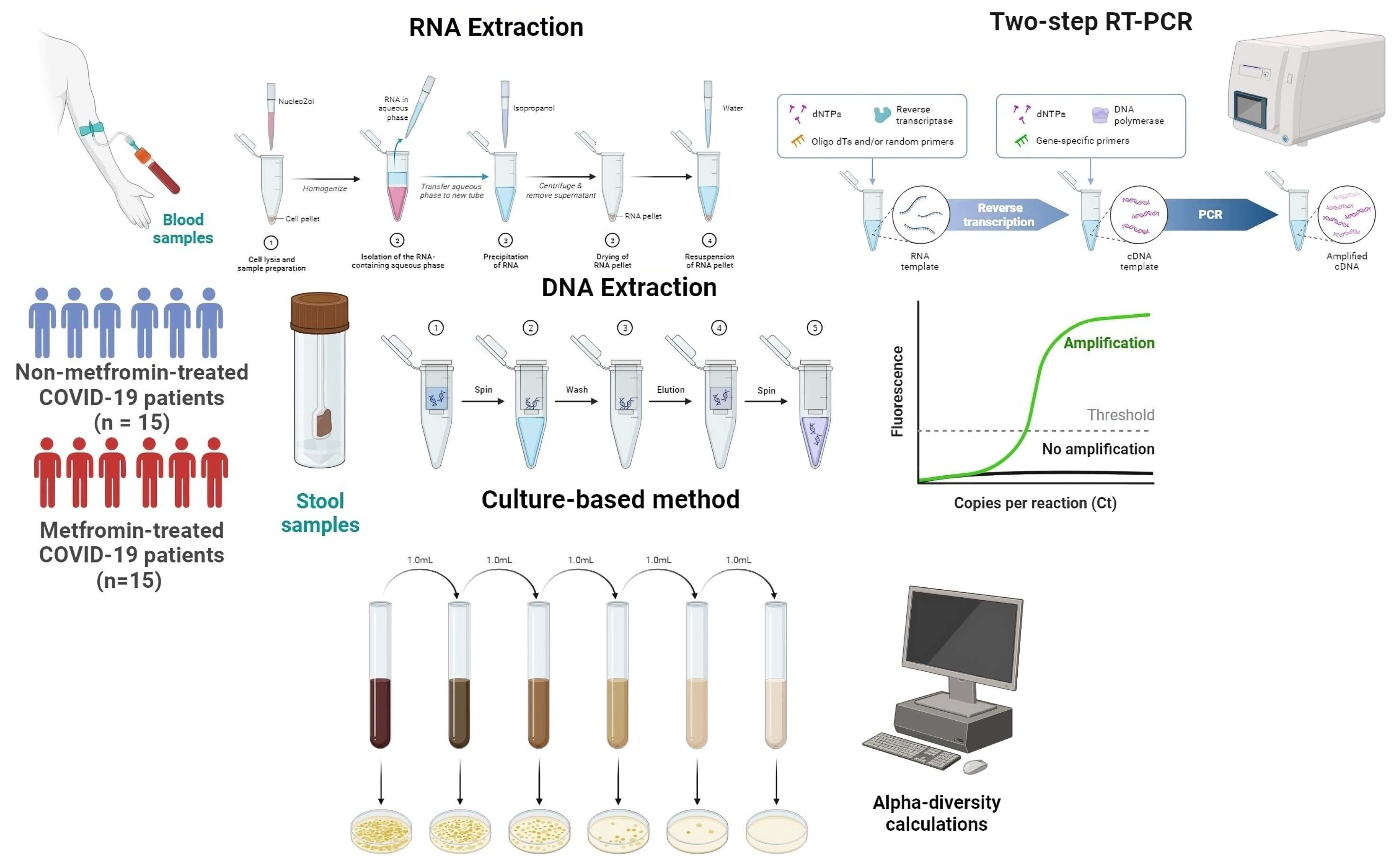
2.1. Sample Collection
2.2. Clinical Data
2.3. Gene Expression Analysis
2.3.1. RNA Extraction and cDNA Synthesis
2.3.2. Real-Time PCR Amplification
2.4. Gut Microbiota Analysis
2.4.1. Firmicutes/Bacteroidetes Ratio
2.4.2. Alpha-Diversity Indices Calculations
2.5. Statistical Analysis
3. Results
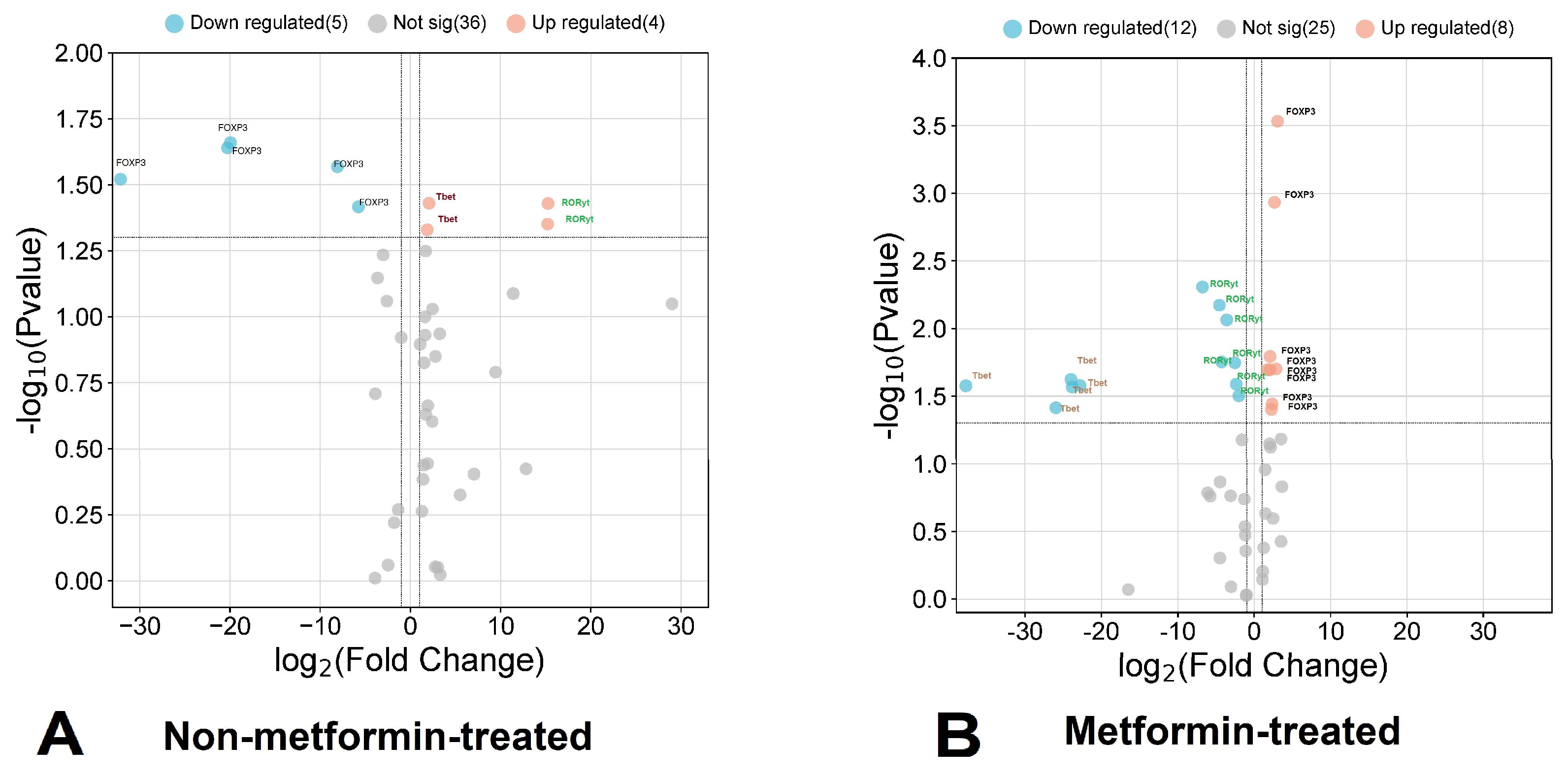
3.1. Relative Expression of FOXP3, RORC, and TBX21 in Metformin-Treated COVID-19 Patients
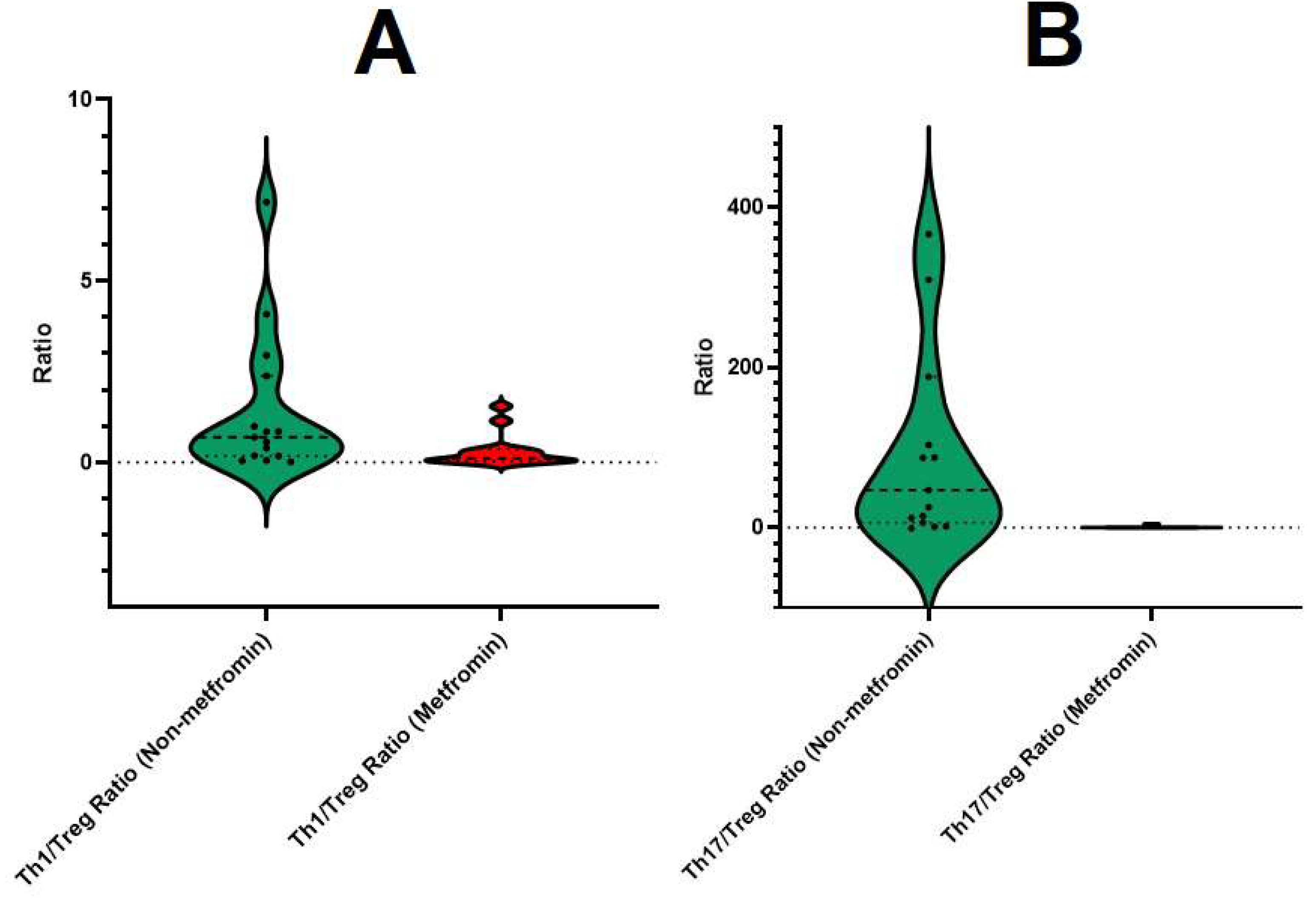
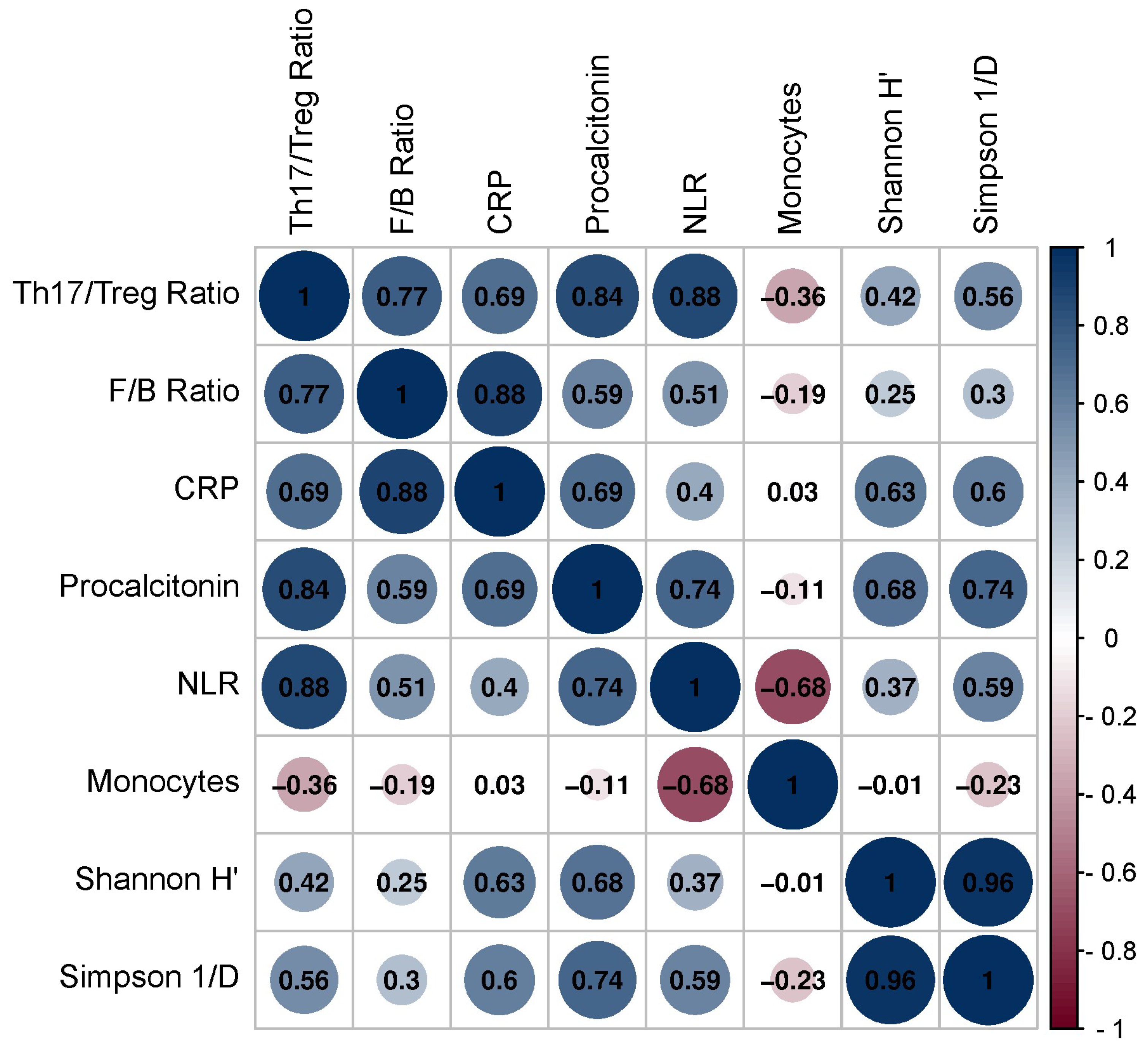
3.2. Correlation between Th1/Treg, Th17/Treg mRNA Ratios, and Gut Microbiota Composition and Hematological Parameters
4. Discussion
5. Conclusions
6. Limitation
- Sample size and homogeneity. The sample size in this study may limit the generalizability of the results. A larger and more diverse cohort would strengthen the statistical power. The study participants were recruited from a single medical center, which may affect the representativeness of the findings. A multi-center approach involving different geographic locations and demographic groups could enhance the external validity of the results.
- Confounders and co-morbidities. The presence of confounding factors and co-morbidities may influence the observed associations. While efforts were made to exclude participants with chronic diseases and other gastrointestinal disorders, the influence of uncontrolled confounders cannot be entirely ruled out. This study did not systematically control for the influence of obesity and dietary habits on the gut microbiota and inflammatory markers. Both obesity and diet are known to be crucial factors influencing microbiome diversity and immune responses.
- Gut microbiota analysis. This study employed a culture-based method for calculating alpha-diversity indices, providing insights into the relative abundance of specific bacterial taxa. However, it is crucial to acknowledge that culture-based methods have limitations in capturing the entire spectrum of microbial diversity present in the gut.
- Cross-sectional design. The cross-sectional design of this study limits the establishment of causal relationships. Longitudinal studies would provide a more dynamic understanding of the relationship between metformin use, gut microbiota, inflammatory markers, and gene expression levels over time.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hiscott, J.; Alexandridi, M.; Muscolini, M.; Tassone, E.; Palermo, E.; Soultsioti, M.; Zevini, A. The global impact of the coronavirus pandemic. Cytokine Growth Factor Rev. 2020, 53, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.worldometers.info/coronavirus/ (accessed on 10 December 2023).
- Looi, M.-K. COVID-19: Scientists sound alarm over new BA.2.86 “Pirola” variant. BMJ Br. Med. J. 2023, 382, 1964. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Behl, T.; Sharma, N.; Singh, S.; Grewal, A.S.; Albarrati, A.; Albratty, M.; Meraya, A.M.; Bungau, S. COVID-19 and diabetes: Association intensify risk factors for morbidity and mortality. Biomed. Pharmacother. 2022, 151, 113089. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Baranova, A.; Wei, X.; Wang, C.; Zhang, F. Bidirectional causal associations between type 2 diabetes and COVID-19. J. Med. Virol. 2023, 95, e28100. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Li, C.; Sun, Y.; Wang, D.W. Insulin treatment is associated with increased mortality in patients with COVID-19 and type 2 diabetes. J. Cell Metab. 2021, 33, 65–77.e2. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Y.; Yi, F.; Zhang, J.; Xu, Z.; Liu, Y.; Tao, Y. Type 2 diabetes mellitus impaired nasal immunity and increased the risk of hyposmia in COVID-19 mild pneumonia patients. Int. Immunopharmacol. 2021, 93, 107406. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Kamyshna, I.; Nykyforuk, A.; Yao, R.; Imbery, J.F.; Oksenych, V.; Korda, M.; Kamyshnyi, A. Immunoregulatory Intestinal Microbiota and COVID-19 in Patients with Type Two Diabetes: A Double-Edged Sword. J. Viruses 2022, 14, 477. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Kamyshna, I.; Kamyshnyi, A. Unveiling the potential pleiotropic effects of metformin in treating COVID-19: A comprehensive review. Front. Mol. Biosci. 2023, 10, 1260633. [Google Scholar] [CrossRef]
- Kamyshnyi, O.; Matskevych, V.; Lenchuk, T.; Strilbytska, O.; Storey, K.; Lushchak, O. Metformin to decrease COVID-19 severity and mortality: Molecular mechanisms and therapeutic potential. Biomed. Pharmacother. 2021, 144, 112230. [Google Scholar] [CrossRef]
- Petakh, P.; Griga, V.; Mohammed, I.B.; Loshak, K.; Poliak, I.; Kamyshnyiy, A. Effects of Metformin, Insulin on Hematological Parameters of COVID-19 Patients with Type 2 Diabetes. Med. Arch. 2022, 76, 329–332. [Google Scholar] [CrossRef]
- Petakh, P.; Loshak, K.; Kamyshnyi, A. Hematological features of patients with type 2 diabetes depending on the variant of SARS-CoV-2. Fiziolohichnyĭ Zhurnal 2023, 69, 35–42. [Google Scholar] [CrossRef]
- Petakh, P.; Isevych, V.; Mohammed, I.; Loshak, K.; Poliak, I.; Kamyshnyiy, A. Association between Use of Metformin and Insulin with Hematological Parameters in COVID-19 Patients with Type 2 Diabetes: A Single Center, Cross-Sectional Study. Clin. Diabetol. 2022, 11, 432–433. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, Y.S.; Wang, L.C.; Huang, J.B. Advances in metformin–based metabolic therapy for non–small cell lung cancer (Review). Oncol. Rep. 2022, 47, 55. [Google Scholar] [CrossRef] [PubMed]
- Chi, H. Regulation and function of mTOR signalling in T cell fate decisions. Nat. Rev. Immunol. 2012, 12, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Blagih, J.; Coulombe, F.; Vincent, E.E.; Dupuy, F.; Galicia-Vázquez, G.; Yurchenko, E.; Raissi, T.C.; van der Windt, G.J.; Viollet, B.; Pearce, E.L.; et al. The Energy Sensor AMPK Regulates T Cell Metabolic Adaptation and Effector Responses In Vivo. Immunity 2015, 42, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Tulipano, G. Integrated or Independent Actions of Metformin in Target Tissues Underlying Its Current Use and New Possible Applications in the Endocrine and Metabolic Disorder Area. Int. J. Mol. Sci. 2021, 22, 13068. [Google Scholar] [CrossRef] [PubMed]
- Ma, E.H.; Poffenberger, M.C.; Wong, A.H.; Jones, R.G. The role of AMPK in T cell metabolism and function. Curr. Opin. Immunol. 2017, 46, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Chen, H. Metformin: A Novel Weapon Against Inflammation. Front. Pharmacol. 2021, 12, 622262. [Google Scholar] [CrossRef] [PubMed]
- Rudensky, A.Y. Regulatory T cells and Foxp3. Immunol. Rev. 2011, 241, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Oestreich, K.J.; Weinmann, A.S. Transcriptional mechanisms that regulate T helper 1 cell differentiation. Curr. Opin. Immunol. 2012, 24, 191–195. [Google Scholar] [CrossRef]
- Capone, A.; Volpe, E. Transcriptional Regulators of T Helper 17 Cell Differentiation in Health and Autoimmune Diseases. Front. Immunol. 2020, 11, 348. [Google Scholar] [CrossRef]
- Martonik, D.; Parfieniuk-Kowerda, A.; Rogalska, M.; Flisiak, R. The Role of Th17 Response in COVID-19. Cells 2021, 10, 1550. [Google Scholar] [CrossRef] [PubMed]
- Kondĕlková, K.; Vokurková, D.; Krejsek, J.; Borská, L.; Fiala, Z.; Ctirad, A. Regulatory T cells (TREG) and their roles in immune system with respect to immunopathological disorders. Acta Medica 2010, 53, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Kobyliak, N.; Kamyshnyi, A. Gut microbiota in patients with COVID-19 and type 2 diabetes: A culture-based method. Front. Cell. Infect. Microbiol. 2023, 13, 1142578. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Kamyshna, I.; Kamyshnyi, A. Effects of metformin on the gut microbiota: A systematic review. Mol. Metab. 2023, 77, 101805. [Google Scholar] [CrossRef] [PubMed]
- Bacchetti De Gregoris, T.; Aldred, N.; Clare, A.S.; Burgess, J.G. Improvement of phylum- and class-specific primers for real-time PCR quantification of bacterial taxa. J. Microbiol. Methods 2011, 86, 351–356. [Google Scholar] [CrossRef]
- Guidelines for Biochemical Identification of Aerobic Bacteria. In Clinical Microbiology Procedures Handbook; ASM Press: Washington, DC, USA, 2016; pp. 3.16.1–3.16.5.
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Polidori, I.; Marullo, L.; Ialongo, C.; Tomassetti, F.; Colombo, R.; di Gaudio, F.; Calugi, G.; Marrone, G.; Noce, A.; Bernardini, S.; et al. Characterization of Gut Microbiota Composition in Type 2 Diabetes Patients: A Population-Based Study. Int. J. Environ. Res. Public Health 2022, 19, 15913. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Oksenych, V.; Kamyshnyi, A. The F/B ratio as a biomarker for inflammation in COVID-19 and T2D: Impact of metformin. Biomed. Pharmacother. 2023, 163, 114892. [Google Scholar] [CrossRef]
- Petakh, P.; Kamyshna, I.; Oksenych, V.; Kainov, D.; Kamyshnyi, A. Metformin Therapy Changes Gut Microbiota Alpha-Diversity in COVID-19 Patients with Type 2 Diabetes: The Role of SARS-CoV-2 Variants and Antibiotic Treatment. Pharmaceuticals 2023, 16, 904. [Google Scholar] [CrossRef]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Hyttinen, J.M.; Kaarniranta, K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: Impact on healthspan and lifespan. J. Mol. Med. 2011, 89, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Petakh, P.; Kamyshna, I.; Kamyshnyi, A. Gene expression of protein kinase AMP-activated catalytic subunit alpha 1 (PRKAA1), solute carrier family 2 member 1 (SLC2A1) and mechanistic target of rapamycin (MTOR) in metformin-treated type 2 diabetes patients with COVID-19: Impact on inflammation markers. Inflammopharmacology 2023, 1–7. [Google Scholar] [CrossRef]
- Ventura-López, C.; Cervantes-Luevano, K.; Aguirre-Sánchez, J.S.; Flores-Caballero, J.C.; Alvarez-Delgado, C.; Bernaldez-Sarabia, J.; Sánchez-Campos, N.; Lugo-Sánchez, L.A.; Rodríguez-Vázquez, I.C.; Sander-Padilla, J.G.; et al. Treatment with metformin glycinate reduces SARS-CoV-2 viral load: An in vitro model and randomized, double-blind, Phase IIb clinical trial. Biomed. Pharmacother. 2022, 152, 113223. [Google Scholar] [CrossRef]
- Reis, G.; Silva, E.A.D.S.M.; Silva, D.C.M.; Thabane, L.; Milagres, A.C.; Ferreira, T.S.; dos Santos, C.V.Q.; Neto, A.D.d.F.; Callegari, E.D.; Savassi, L.C.M.; et al. Effect of early treatment with metformin on risk of emergency care and hospitalization among patients with COVID-19: The TOGETHER randomized platform clinical trial. Lancet Reg. Health Am. 2022, 6, 100142. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Primer Nucleotide Sequence | |
|---|---|---|
| Forward | Reverse | |
| Firmicutes | 928F-firm TGAAACTYAAGGAATTGACG | 1040FirmR ACCATGCACCACCTGTC |
| Bacteroidetes | 798cfbF CRAACAGGATTAGATACCCT | cfb967R GGTAAGGTTCCTCGCGCTAT |
| 16S rRNA gene | 926F AAACTCAAAKGAATTGACGG | 1062R CTCACRRCACGAGCTGAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petakh, P.; Kamyshna, I.; Oksenych, V.; Kamyshnyi, O. Metformin Alters mRNA Expression of FOXP3, RORC, and TBX21 and Modulates Gut Microbiota in COVID-19 Patients with Type 2 Diabetes. Viruses 2024, 16, 281. https://doi.org/10.3390/v16020281
Petakh P, Kamyshna I, Oksenych V, Kamyshnyi O. Metformin Alters mRNA Expression of FOXP3, RORC, and TBX21 and Modulates Gut Microbiota in COVID-19 Patients with Type 2 Diabetes. Viruses. 2024; 16(2):281. https://doi.org/10.3390/v16020281
Chicago/Turabian StylePetakh, Pavlo, Iryna Kamyshna, Valentyn Oksenych, and Oleksandr Kamyshnyi. 2024. "Metformin Alters mRNA Expression of FOXP3, RORC, and TBX21 and Modulates Gut Microbiota in COVID-19 Patients with Type 2 Diabetes" Viruses 16, no. 2: 281. https://doi.org/10.3390/v16020281
APA StylePetakh, P., Kamyshna, I., Oksenych, V., & Kamyshnyi, O. (2024). Metformin Alters mRNA Expression of FOXP3, RORC, and TBX21 and Modulates Gut Microbiota in COVID-19 Patients with Type 2 Diabetes. Viruses, 16(2), 281. https://doi.org/10.3390/v16020281