
Torque Teno Virus DNA Load in Blood as an Immune Status Biomarker in Adult Hematological Patients: The State of the Art and Future Prospects
 , and
, and
Abstract
1. Introduction
2. Methodological Considerations
3. TTV DNA Load in Blood as a Biomarker of the Net State of Immunosuppression in Allo-SCT
3.1. Frequency of TTV DNA Detection in Allo-SCT Recipients
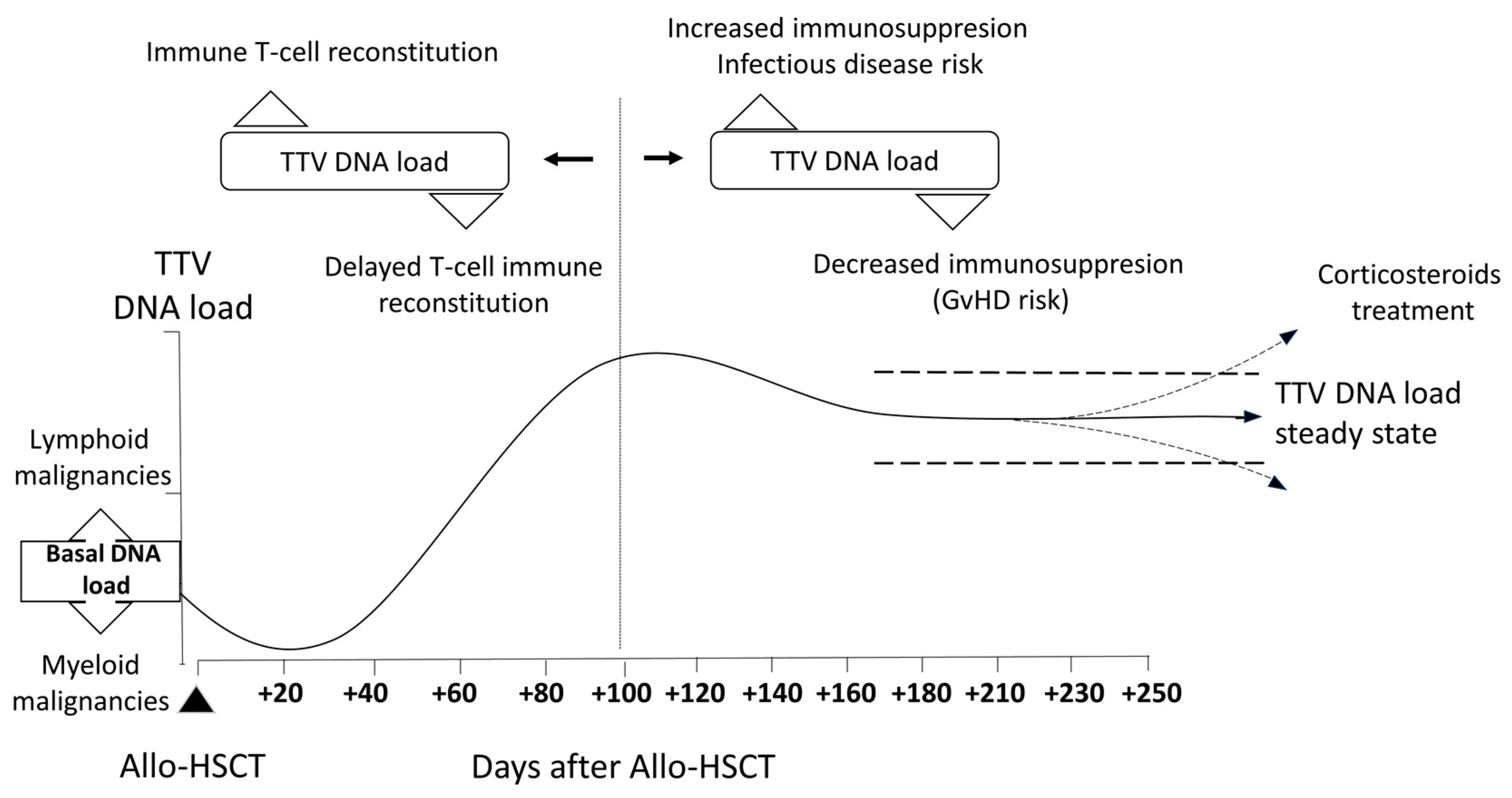
3.2. TTV DNAemia Kinetics in Allo-SCT Recipients
3.3. TTV DNA Load and Occurrence of Clinically Relevant Events following Allo-SCT
3.3.1. TTV DNA Load and Opportunistic Viral Infections
3.3.2. TTV DNA Load and Graft versus Host Disease
3.3.3. TTV DNA Load and Other Clinical Events
4. Potential Clinical Value of TTV DNA Load Assessment in Other Hematological Patients
5. Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Bendinelli, M.; Pistello, M.; Maggi, F.; Fornai, C.; Freer, G.; Vatteroni, M.L. Molecular properties, biology, and clinical implications of TT virus, a recently identified widespread infectious agent of humans. Clin. Microbiol. Rev. 2001, 14, 98–113. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Opriessnig, T.; Celer, V.; Maggi, F.; Okamoto, H.; Blomström, A.L.; Cadar, D.; Harrach, B.; Biagini, P.; Kraberger, S. Taxonomic update for mammalian anelloviruses (family Anelloviridae). Arch. Virol. 2021, 166, 2943–2953. [Google Scholar] [CrossRef] [PubMed]
- Varsani, A.; Kraberger, S.; Opriessnig, T.; Maggi, F.; Celer, V.; Okamoto, H.; Biagini, P. Anelloviridae taxonomy update 2023. Arch. Virol. 2023, 168, 277. [Google Scholar] [CrossRef] [PubMed]
- Arze, C.A.; Springer, S.; Dudas, G.; Patel, S.; Bhattacharyya, A.; Swaminathan, H.; Brugnara, C.; Delagrave, S.; Ong, T.; Kahvejian, A.; et al. Global genome analysis reveals a vast and dynamic anellovirus landscape within the human virome. Cell Host Microbe 2021, 29, 1305–1315.e6. [Google Scholar] [CrossRef]
- Focosi, D.; Antonelli, G.; Pistello, M.; Maggi, F. Torquetenovirus: The human virome from bench to bedside. Clin. Microbiol. Infect. 2016, 22, 589–593. [Google Scholar] [CrossRef]
- Focosi, D.; Spezia, P.G.; Macera, L.; Salvadori, S.; Navarro, D.; Lanza, M.; Antonelli, G.; Pistello, M.; Maggi, F. Assessment of prevalence and load of torquetenovirus viraemia in a large cohort of healthy blood donors. Clin. Microbiol. Infect. 2020, 26, 1406–1410. [Google Scholar] [CrossRef]
- Kaczorowska, J.; Deijs, M.; Klein, M.; Bakker, M.; Jebbink, M.F.; Sparreboom, M.; Kinsella, C.M.; Timmerman, A.L.; van der Hoek, L. Diversity and Long-Term Dynamics of Human Blood Anelloviruses. J. Virol. 2022, 96, e0010922. [Google Scholar] [CrossRef]
- Maggi, F.; Fornai, C.; Zaccaro, L.; Morrica, A.; Vatteroni, M.L.; Isola, P.; Marchi, S.; Ricchiuti, A.; Pistello, M.; Bendinelli, M. TT virus (TTV) loads associated with different peripheral blood cell types and evidence for TTV replication in activated mononuclear cells. J. Med. Virol. 2001, 64, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.; Pal, R.; Deshmukh, R.; Banker, D. Replication of TT virus in hepatocyte and leucocyte cell lines. J. Med. Virol. 2005, 77, 136–143. [Google Scholar] [CrossRef]
- Mariscal, L.F.; López-Alcorocho, J.M.; Rodríguez-Iñigo, E.; Ortiz-Movilla, N.; de Lucas, S.; Bartolomé, J.; Carreño, V. T virus replicates in stimulated but not in nonstimulated peripheral blood mononuclear cells. Virology 2002, 301, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, V.; Laperche, S.; Cheval, J.; Muth, E.; Dubois, M.; Boizeau, L.; Hebert, C.; Lionnet, F.; Lefrere, J.J.; Eloit, M. Viral metagenomics applied to blood donors and recipients at high risk for blood-borne infections. Blood Transfus. 2016, 14, 400–407. [Google Scholar]
- Takahashi, M.; Asabe, S.; Gotanda, Y.; Kishimoto, J.; Tsuda, F.; Okamoto, H. TT virus is distributed in various leukocyte subpopulations at distinct levels, with the highest viral load in granulocytes. Biochem. Biophys. Res. Commun. 2002, 290, 242–248. [Google Scholar] [CrossRef]
- Kikuchi, K.; Miyakawa, H.; Abe, K.; Kako, M.; Katayama, K.; Fukushi, S.; Mishiro, S. Indirect evidence of TTV replication in bone marrow cells, but not in hepatocytes, of a subacute hepatitis/aplastic anemia patient. J. Med. Virol. 2000, 61, 165–170. [Google Scholar] [CrossRef]
- Kaczorowska, J.; van der Hoek, L. Human anelloviruses: Diverse, omnipresent and commensal members of the virome. FEMS Microbial. Rev. 2020, 44, 305–313. [Google Scholar] [CrossRef]
- De Vlaminck, I.; Khush, K.K.; Strehl, C.; Kohli, B.; Luikart, H.; Neff, N.F.; Okamoto, J.; Snyder, T.M.; Cornfield, D.N.; Nicolls, M.R.; et al. Temporal response of the human virome to immunosuppression and antiviral therapy. Cell 2013, 155, 1178–1187. [Google Scholar] [CrossRef]
- Redondo, N.; Navarro, D.; Aguado, J.M.; Fernández-Ruiz, M. Viruses, friends, and foes: The case of Torque Teno Virus and the net state of immunosuppression. Transpl. Infect. Dis. 2022, 24, e1377. [Google Scholar] [CrossRef]
- van Rijn, A.L.; Roos, R.; Dekker, F.W.; Rotmans, J.I.; Feltkamp, M. Torque teno virus load as marker of rejection and infection in solid organ transplantation—A systematic review and meta-analysis. Rev. Med. Virol. 2023, 33, e2393. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Pifferi, M.; Fornai, C.; Andreoli, E.; Tempestini, E.; Vatteroni, M.; Presciuttini, S.; Marchi, S.; Pietrobelli, A.; Boner, A.; et al. TT virus in the nasal secretions of children with acute respiratory diseases: Relations to viremia and disease severity. J. Virol. 2003, 77, 2418–2425. [Google Scholar] [CrossRef]
- Ninomiya, M.; Takahashi, M.; Nishizawa, T.; Shimosegawa, T.; Okamoto, H. Development of PCR assays with nested primers specific for differential detection of three humananelloviruses and early acquisition of dual or triple infection during infancy. J. Clin. Microbiol. 2008, 46, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Macera, L.; Spezia, P.G.; Medici, C.; Rofi, E.; Del Re, M.; Focosi, D.; Mazzetti, P.; Navarro, D.; Antonelli, G.; Danesi, R.; et al. Comparative evaluation of molecular methods for the quantitative measure of torquetenovirus viremia, the new surrogate marker of immune competence. J. Med. Virol. 2022, 94, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Giménez, E.; Monzó, C.; Albert, E.; Fuentes-Trillo, A.; Seda, E.; Piñana, J.L.; Hernández Boluda, J.C.; Solano, C.; Chaves, J.; Navarro, D. Diversity and dynamic changes of anelloviruses in plasma following allogeneic hematopoietic stem cell transplantation. J. Med. Virol. 2021, 93, 5167–5172. [Google Scholar] [CrossRef]
- Zanella, M.C.; Cordey, S.; Kaiser, L. Beyond Cytomegalovirus and Epstein-Barr Virus: A Review of Viruses Composing the Blood Virome of Solid Organ Transplant and Hematopoietic Stem Cell Transplant Recipients. Clin. Microbiol. Rev. 2020, 33, e00027-20. [Google Scholar] [CrossRef] [PubMed]
- Leijonhufvud, G.; Bajalan, A.; Teixeira Soratto, T.A.; Gustafsson, B.; Bogdanovic, G.; Bjerkner, A.; Allander, T.; Ljungman, G.; Andersson, B. Better detection of Torque teno virus in children with leukemia by metagenomic sequencing than by quantitative PCR. J. Med. Virol. 2022, 94, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Pou, C.; Barrientos-Somarribas, M.; Marin-Juan, S.; Bogdanovic, G.; Bjerkner, A.; Allander, T.; Gustafsson, B.; Andersson, B. Virome definition in cerebrospinal fluid of patients with neurological complications after hematopoietic stem cell transplantation. J. Clin. Virol. 2018, 108, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Edridge, A.W.D.; Deijs, M.; van Zeggeren, I.E.; Kinsella, C.M.; Jebbink, M.F.; Bakker, M.; van de Beek, D.; Brouwer, M.C.; van der Hoek, L. Viral Metagenomics on Cerebrospinal Fluid. Genes 2019, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Masouridi-Levrat, S.; Pradier, A.; Simonetta, F.; Kaiser, L.; Chalandon, Y.; Roosnek, E. Torque teno virus in patients undergoing allogeneic hematopoietic stem cell transplantation for hematological malignancies. Bone Marrow Transplant. 2016, 51, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Albert, E.; Solano, C.; Pascual, T.; Torres, I.; Macera, L.; Focosi, D.; Maggi, F.; Giménez, E.; Amat, P.; Navarro, D. Dynamics of Torque Teno virus plasma DNAemia in allogeneic stem cell transplant recipients. J. Clin. Virol. 2017, 94, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Wohlfarth, P.; Leiner, M.; Schoergenhofer, C.; Hopfinger, G.; Goerzer, I.; Puchhammer-Stoeckl, E.; Rabitsch, W. Torquetenovirus Dynamics and Immune Marker Properties in Patients Following Allogeneic Hematopoietic Stem Cell Transplantation: A Prospective Longitudinal Study. Biol. Blood Marrow Transplant. 2018, 24, 194–199. [Google Scholar] [CrossRef]
- Schmitz, J.; Kobbe, G.; Kondakci, M.; Schuler, E.; Magorsch, M.; Adams, O. The Value of Torque Teno Virus (TTV) as a Marker for the Degree of Immunosuppression in Adult Patients after Hematopoietic Stem Cell Transplantation (HSCT). Biol. Blood Marrow Transplant. 2020, 26, 643–650. [Google Scholar] [CrossRef]
- Spiertz, A.; Tsakmaklis, A.; Farowski, F.; Knops, E.; Heger, E.; Wirtz, M.; Kaiser, R.; Holtick, U.; Vehreschild, M.J.G.T.; Di Cristanziano, V. Torque teno virus-DNA load as individual cytomegalovirus risk assessment parameter upon allogeneic hematopoietic stem cell transplantation. Eur. J. Haematol. 2023, 111, 963–969. [Google Scholar] [CrossRef]
- Zanella, M.C.; Vu, D.L.; Hosszu-Fellous, K.; Neofytos, D.; Van Delden, C.; Turin, L.; Poncet, A.; Simonetta, F.; Masouridi-Levrat, S.; Chalandon, Y.; et al. Longitudinal Detection of Twenty DNA and RNA Viruses in Allogeneic Hematopoietic Stem Cell Transplant Recipients Plasma. Viruses 2023, 15, 928. [Google Scholar] [CrossRef] [PubMed]
- Mouton, W.; Conrad, A.; Bal, A.; Boccard, M.; Malcus, C.; Ducastelle-Lepretre, S.; Balsat, M.; Barraco, F.; Larcher, M.V.; Fossard, G.; et al. Torque Teno Virus Viral Load as a Marker of Immune Function in Allogeneic Haematopoietic Stem Cell Transplantation Recipients. Viruses 2020, 12, 1292. [Google Scholar] [CrossRef] [PubMed]
- Gilles, R.; Herling, M.; Holtick, U.; Heger, E.; Awerkiew, S.; Fish, I.; Höller, K.; Sierra, S.; Knops, E.; Kaiser, R.; et al. Dynamics of Torque Teno virus viremia could predict risk of complications after allogeneic hematopoietic stem cell transplantation. Med. Microbiol. Immunol. 2017, 206, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Focosi, D.; Albani, M.; Lanini, L.; Vatteroni, M.L.; Petrini, M.; Ceccherini-Nelli, L.; Pistello, M.; Bendinelli, M. Role of hematopoietic cells in the maintenance of chronic human torquetenovirus plasma viremia. J. Virol. 2010, 84, 6891–6893. [Google Scholar] [CrossRef]
- Albert, E.; Torres, I.; Talaya, A.; Giménez, E.; Piñana, J.L.; Hernández-Boluda, J.C.; Focosi, D.; Macera, L.; Maggi, F.; Solano, C.; et al. Kinetics of torque teno virus DNA load in saliva and plasma following allogeneic hematopoietic stem cell transplantation. J. Med. Virol. 2018, 90, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- Albert, E.; Solano, C.; Giménez, E.; Focosi, D.; Pérez, A.; Macera, L.; Piñana, J.L.; Mateo, E.M.; Boluda, J.C.H.; Maggi, F.; et al. Kinetics of Alphatorquevirus plasma DNAemia at late times after allogeneic hematopoietic stem cell transplantation. Med. Microbiol. Immunol. 2019, 208, 253–258. [Google Scholar] [CrossRef]
- Pradier, A.; Masouridi-Levrat, S.; Bosshard, C.; Dantin, C.; Vu, D.-L.; Zanella, M.-C.; Boely, E.; Tapparel, C.; Kaiser, L.; Chalandon, Y.; et al. Torque teno virus as a potential biomarker for complications and survival after allogeneic hematopoietic stem cell transplantation. Front. Immunol. 2020, 11, 998. [Google Scholar] [CrossRef]
- Forqué, L.; Albert, E.; Piñana, J.L.; Pérez, A.; Hernani, R.; Solano, C.; Navarro, D.; Giménez, E. Monitoring of plasma Torque teno virus, total Anelloviridae and Human Pegivirus 1 viral load for the prediction of infectious events and acute graft versus host disease in the allogeneic hematopoietic stem cell transplantation setting. J. Med. Virol. 2023, 95, e29107. [Google Scholar] [CrossRef]
- Storek, J.; Dawson, M.A.; Storer, B.; Stevens-Ayers, T.; Maloney, D.G.; Marr, K.A.; Witherspoon, R.P.; Bensinger, W.; Flowers, M.E.; Martin, P.; et al. Immune reconstitution after allogeneic marrow transplantation compared with blood stem cell transplantation. Blood 2001, 97, 3380–3389. [Google Scholar] [CrossRef]
- Storek, J.; Geddes, M.; Khan, F.; Huard, B.; Helg, C.; Chalandon, Y.; Passweg, J.; Roosnek, E. Reconstitution of the immune system after hematopoietic stem cell transplantation in humans. Semin. Immunopathol. 2008, 30, 425–437. [Google Scholar] [CrossRef]
- Focosi, D.; Bestagno, M.; Burrone, O.; Petrini, M. CD57+ T lymphocytes and functional immune deficiency. J. Leukoc. Biol. 2010, 87, 107–116. [Google Scholar] [CrossRef]
- Albert, E.; Solano, C.; Giménez, E.; Focosi, D.; Pérez, A.; Macera, L.; Piñana, J.L.; Hernández-Boluda, J.C.; Maggi, F.; Navarro, D. The kinetics of torque teno virus plasma DNA load shortly after engraftment predicts the risk of high-level CMV DNAemia in allogeneic hematopoietic stem cell transplant recipients. Bone Marrow Transplant. 2018, 53, 180–187. [Google Scholar] [CrossRef]
- Legoff, J.; Resche-Rigon, M.; Bouquet, J.; Robin, M.; Naccache, S.N.; Mercier-Delarue, S.; Federman, S.; Samayoa, E.; Rousseau, C.; Piron, P.; et al. The eukaryotic gut virome in hematopoietic stem cell transplantation: New clues in enteric graft-versus-host disease. Nat. Med. 2017, 23, 1080–1085. [Google Scholar] [CrossRef]
- Bueno, F.; Albert, E.; Piñana, J.L.; Pérez, A.; Úbeda, C.; Gómez, M.D.; Hernández-Boluda, J.C.; Gonzalez-Barberá, E.M.; Montoro, J.; Giménez, E.; et al. Kinetics of Torque Teno virus DNA in stools may predict occurrence of acute intestinal graft versus host disease early after allogeneic hematopoietic stem cell transplantation. Transpl. Infect. Dis. 2021, 23, e13507. [Google Scholar] [CrossRef] [PubMed]
- Focosi, D.; Maggi, F.; Albani, M.; Macera, L.; Ricci, V.; Gragnani, S.; Di Beo, S.; Ghimenti, M.; Antonelli, G.; Bendinelli, M.; et al. Torquetenovirus viremia kinetics after autologous stem cell transplantation are predictable and may serve as a surrogate marker of functional immune reconstitution. J. Clin. Virol. 2010, 47, 189–192. [Google Scholar] [CrossRef] [PubMed]
- Mitra, A.; Barua, A.; Huang, L.; Ganguly, S.; Feng, Q.; He, B. From bench to bedside: The history and progress of CAR T cell therapy. Front. Immunol. 2023, 14, 1188049. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Cao, Y.J. Engineered T Cell Therapy for Cancer in the Clinic. Front. Immunol. 2019, 10, 2250. [Google Scholar] [CrossRef]
- Turtle, C.J.; Hanafi, L.-A.; Berger, C.; Gooley, T.A.; Cherian, S.; Hudecek, M.; Sommermeyer, D.; Melville, K.; Pender, B.; Budiarto, T.M.; et al. CD19 CAR-T cells of defined CD4+:CD8+ composition in adult B cell ALL patients. J. Clin. Investig. 2016, 126, 2123–2138. [Google Scholar] [CrossRef] [PubMed]
- Wudhikarn, K.; Palomba, M.L.; Pennisi, M.; Garcia-Recio, M.; Flynn, J.R.; Devlin, S.M.; Devlin, S.M.; Afuye, A.; Silverberg, M.L.; Maloy, M.A.; et al. Infection during the first year in patients treated with CD19 CAR T cells for diffuse large B cell lymphoma. Blood Cancer J. 2020, 10, 79. [Google Scholar] [CrossRef]
- Baird, J.H.; Epstein, D.J.; Tamaresis, J.S.; Ehlinger, Z.; Spiegel, J.Y.; Craig, J.; Claire, G.K.; Frank, M.J.; Muffly, L.; Shiraz, P.; et al. Immune reconstitution and infectious complications following axicabtagene ciloleucel therapy for large B-cell lymphoma. Blood Adv. 2021, 5, 143–155. [Google Scholar] [CrossRef]
- Benzaquén, A.; Giménez, E.; Iacoboni, G.; Guerreiro, M.; Hernani, R.; Albert, E.; Carpio, C.; Balaguer, A.; Pérez, A.; Solano de la Asunción, C.; et al. Torque Teno Virus plasma DNA load: A novel prognostic biomarker in CAR-T therapy. Bone Marrow Transplant. 2024, 59, 93–100. [Google Scholar] [CrossRef]
- Hernani, R.; Benzaquén, A.; Solano, C. Toxicities following CAR-T therapy for hematological malignancies. Cancer Treat. Rev. 2022, 111, 102479. [Google Scholar] [CrossRef]
- Kunte, A.S.; Gagelmann, N.; Badbaran, A.; Berger, C.; Massoud, R.; Klyuchnikov, E. Viral Reactivation and Immune Reconstitution after CAR-T Cell Treatment in Patients with Hematologic Malignancies. Blood 2022, 140 (Suppl. S1), 7531–7532. [Google Scholar] [CrossRef]
- Logue, J.M.; Zucchetti, E.; Bachmeier, C.A.; Krivenko, G.S.; Larson, V.; Ninh, D.; Grillo, G.; Cao, B.; Kim, J.; Chavez, J.C.; et al. Immune reconstitution and associated infections following axicabtagene ciloleucel in relapsed or refractory large B-cell lymphoma. Haematologica 2021, 106, 978–986. [Google Scholar] [CrossRef]
- Cordeiro, A.; Bezerra, E.D.; Hirayama, A.V.; Hill, J.A.; Wu, Q.V.; Voutsinas, J.; Sorror, M.L.; Turtle, C.J.; Maloney, D.G.; Bar, M. Late Events after Treatment with CD19-Targeted Chimeric Antigen Receptor Modified T Cells. Biol. Blood Marrow Transplant. 2020, 26, 26–33. [Google Scholar] [CrossRef]
- Sochacka-Ćwikła, A.; Mączyński, M.; Regiec, A. FDA-approved drugs for hematological malignancies-the last decade review. Cancers 2021, 14, 87. [Google Scholar] [CrossRef]
- Ruiz-Camps, I.; Aguilar-Company, J. Risk of infection associated with targeted therapies for solid organ and hematological malignancies. Ther. Adv. Infect. Dis. 2021, 8, 2049936121989548. [Google Scholar] [CrossRef]
- de la Asunción, C.S.; Giménez, E.; Hernández-Boluda, J.C.; Terol, M.J.; Albert, E.; López, J.; García-Gutiérrez, V.; Andreu, R.; Malo, M.D.G.; Fox, M.L.; et al. Assessment of the potential value of plasma Torque Teno virus DNA load monitoring to predict cytomegalovirus DNAemia in patients with hematological malignancies treated with small molecule inhibitors: A proof-of-concept study. J. Med. Virol. 2023, 95, e28933. [Google Scholar] [CrossRef] [PubMed]
- de la Asunción Carlos, S.; Giménez, E.; Hernández-Boluda, J.C.; Terol, M.J.; Albert, E.; López-Jiménez, J.; García-Gutiérrez, V.; Andreu, R.; García, D.; Fox, M.L.; et al. Immunobiology of cytomegalovirus infection in patients with haematological malignancies undergoing treatment with small molecule inhibitors. Br. J. Haematol. 2023, 200, e58–e61. [Google Scholar] [CrossRef] [PubMed]
- Solano de la Asunción, C.; Terol, M.J.; Saus, A.; Olea, B.; Giménez, E.; Albert, E.; López-Jiménez, J.; Andreu, R.; García, D.; Fox, L.; et al. Cytomegalovirus-specific T-cell immunity and DNAemia in patients with chronic lymphocytic leukaemia undergoing treatment with ibrutinib. Br. J. Haematol. 2021, 195, 637–641. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Study | Sample Size and Underlying Disease | Monitoring Time Points | Main Findings |
|---|---|---|---|
| Maggi et al. (2010) [34] | Four patients with acute leukemia | Before total body irradiation (TBI) and after allo-SCT (within the first 30 days; in 2 patients at days +50, +80, and +110 after transplantation) |
|
| Masouridi-Levrat et al. (2016) [26] | A total of 121 patients. AML (58), ALL (15), MDS (12), NHL (10), MPS (6), MM (9), HL (5), CML (3), CLL (1), and MDPS (1) | At baseline (days +4.3 ± 5.4 post-transplant) and at 1 and 3 months afterward. |
|
| Albert, et al. (2017) [27] | A total of 72 patients. HL (5), NHL (15), ALL (7), CLL (6), AML (19), CML (1), MM (5), MDS (10), and others (4) | Before conditioning, and at days +20, +30, +60, and +90 after allo-SCT. |
|
| Wohlfarth et al. (2018) [28] | A total of 50 patients. ALM (25); ALL (9) MDS (6), NHL (3), and others (7) | Before conditioning, at the time of allo-SCT and days +10, +30, +50, +80, +120, +160, +200,+ 250, +300, and +365 after allo-SCT |
|
| Albert et al. (2018) [35] | A total of 38 patients. NHL (14), HL (1), AML (4), CLL (3), AML (6), MM (2), MDPS (6), and others (2) | Before the initiation of conditioning (day −7) and days +30, +50, +90 after allo-SSCT. |
|
| Albert et al. (2019) [36] | A total of 33 patients: Lymphoma (12), leukemia (12), MM (4), MDS (3), and myelofibrosis (2) | Before conditioning and at days +20, +30, +40, +50, +60, +90, +120, +180, +210 after allo-SCT |
|
| Mouton et al. (2020) [32] | A total of 41 patients. Myeloid neoplasm and acute leukemia (37), Mature lymphoid/histiocytic, and dendritic neoplasms (4) | Patients were enrolled a median of 6 months (IQR, 5–8) post transplant. |
|
| Schmitz et al. (2020) [29] | A total of 123 patients. AML (58), ALL (9), MDS (33), NHL (11), and others (12) | A total of 18 different time points were examined from before allo-HSCT to 345 days post. |
|
| Pradier et al. (2020) [37] | A total of 168 patients. AML (78), ALL (17), MDS (22), MPS (11), Lymphoma (12), MM (11), and others (17). | Peripheral blood samples were collected at days 0, +50, +100, +150, +200, +300, +400, +547, and 2 to 9 years post-allo-SCT. |
|
| Spiertz et al. (2023) [30] | A total of 59 patients. AML (31) ALL (6), CML (2), CLL (4), MDS (10), and others (6) | Upon infusion of hematopoietic cells (between day −7 and +10), and at days +14, +21, +28, +56, +90, +365 after allo-SCT |
|
| Forque et al. (2023) [38] | A total of 75 patients. AML (28), H (13) NHL (15) MDS (4) MF (4), CML (3), CLL (3), and others (5) | Before conditioning, at baseline, and after allo-SCT (+30, +60, +90, +120, and +180) |
|
| Study | Sample Size and Underlying Disease | Relevant Monitoring Time Points before or after Allo-SCT | Main Findings |
|---|---|---|---|
| Albert E et al. (2018) [42] | A total of 72 patients. HL (5), NHL (15), ALL (7), CLL (6), AML (19), CML (1), MM (5), MDS (10), and others (4). | Days +20, +30, +40, +50 after allo-SCT |
|
| Legoff et al. (2017) [43] | A total of 44 patients. Underlying disease not reported. | Longitudinal follow-up of the enteric virome (by metagenomics) |
|
| Gilles et al. (2017) [33] | A total of 23 patients. AML (10), Lymphoma (7), MDS (1), Myelofibrosis (1), MM (1), CLL (1), prolymphocytic leukemia (1), chronic myelomonocytic leukemia (1) Low risk (9), High risk (14) | Days +30, +100, and +200 |
|
| Schmitz et al. (2020) [29] | A total of 123 patients. AML (58), ALL (9), MDS (33), NHL (11), and others (12) | Days + 0–15, +16–30, +31–45, +46–60, +61–80, +81–99, +100–119, +120–140, +141–160, +161–180, +181–200, +201–219, +221–239, +240–260, +261–280, +281–300, +301–320, +321–345 |
|
| Wohlfarth et al. (2018) [28] | A total of 50 patients. ALM (25); ALL (9) MDS (6), NHL (3), and others (7) | Days +10, +30, +50, +80, +120, +160, +200,+ 250, +300 and +365 |
|
| Mouton W et al. (2020) [32] | A total of 41 patients. Myeloid neoplasm and acute leukemia (37), Mature lymphoid, histiocytic, and dendritic neoplasms (4) | Five to eight months post transplant |
|
| Pradier et al. (2020) [37] | A total of 133 patients. AML (78), ALL (17), MDS (22), MPS (11), Lymphoma (12), Myeloma (11), and others (17) | Days +50, +100, +150, +200, +300, +400, +547, and 2 to 9 years post-allo-SCT. |
|
| Bueno et al. (2021) [44] | A total of 83 patients. ALL (49), CL (4), Lymphoma (4), MDS (19), Myelofibrosis (5), and MM (2) | TTV DNA was quantified in paired stool and plasma samples collected a median of 2 days before cell infusion and a median of 14 days after allo-HSCT by real-time PCR. |
|
| Spiertz et al. (2023) [30] | A total of 59 patients. AML (31), ALL (6), CML (2), CLL (4), MDS (10), and others (6) | Days +14, +21, +28, +56, +90 and +365 |
|
| Forque et al. (2023) [38] | A total of 75 patients. AML (28), HL (13), NHL (15), MDS (4), MF (4), CML (3), CLL (3), and others (5) | Preconditioning, at baseline (day 0) and days after allo-SCT (+30, +60, +90, +120, and +180) |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albert, E.; Giménez, E.; Hernani, R.; Piñana, J.L.; Solano, C.; Navarro, D. Torque Teno Virus DNA Load in Blood as an Immune Status Biomarker in Adult Hematological Patients: The State of the Art and Future Prospects. Viruses 2024, 16, 459. https://doi.org/10.3390/v16030459
Albert E, Giménez E, Hernani R, Piñana JL, Solano C, Navarro D. Torque Teno Virus DNA Load in Blood as an Immune Status Biomarker in Adult Hematological Patients: The State of the Art and Future Prospects. Viruses. 2024; 16(3):459. https://doi.org/10.3390/v16030459
Chicago/Turabian StyleAlbert, Eliseo, Estela Giménez, Rafael Hernani, José Luis Piñana, Carlos Solano, and David Navarro. 2024. "Torque Teno Virus DNA Load in Blood as an Immune Status Biomarker in Adult Hematological Patients: The State of the Art and Future Prospects" Viruses 16, no. 3: 459. https://doi.org/10.3390/v16030459
APA StyleAlbert, E., Giménez, E., Hernani, R., Piñana, J. L., Solano, C., & Navarro, D. (2024). Torque Teno Virus DNA Load in Blood as an Immune Status Biomarker in Adult Hematological Patients: The State of the Art and Future Prospects. Viruses, 16(3), 459. https://doi.org/10.3390/v16030459