
Genetic Characterization of Small Ruminant Lentiviruses Isolated from Dairy Sheep in Greece

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Blood Samples, and DNA Extraction
2.2. PCR, Sequencing, and Phylogenetic Analyses
3. Results
3.1. Pairwise Nucletide and Amino Acid Sequence Comparisons
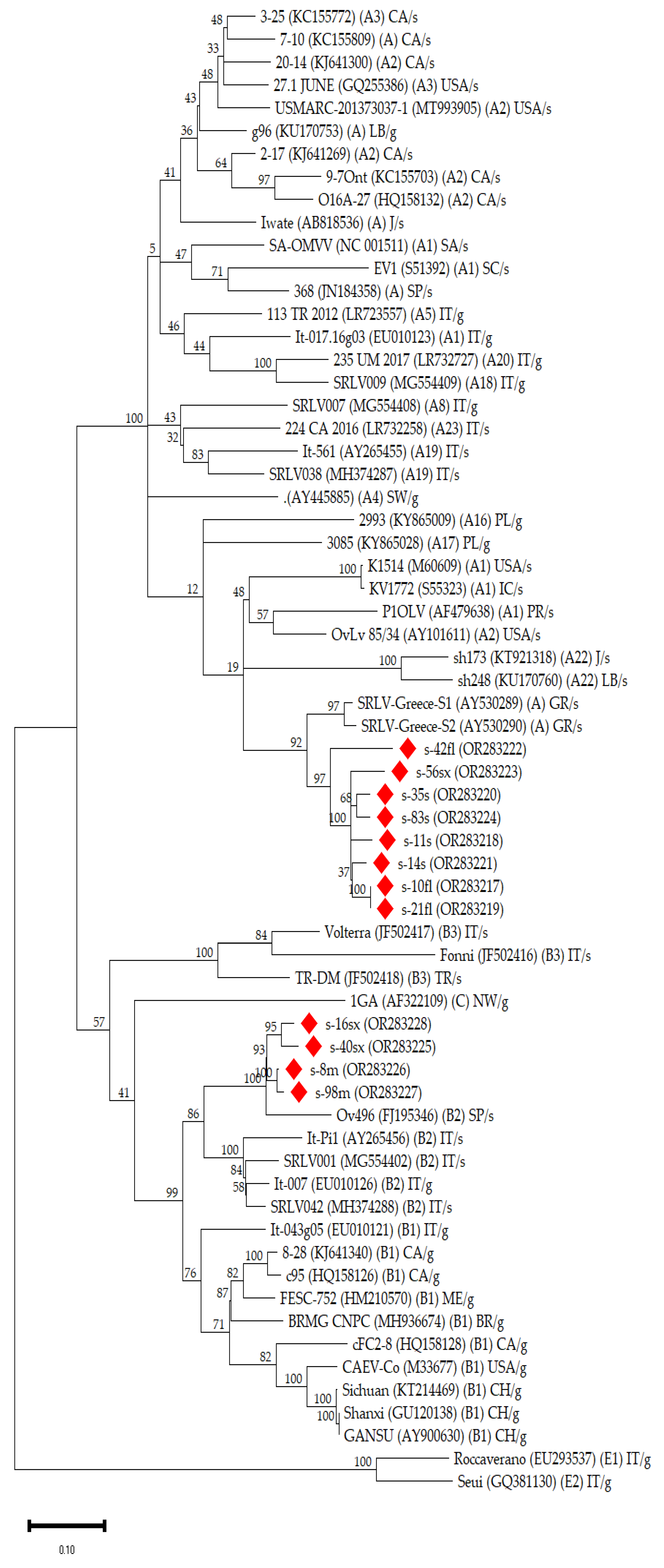
3.2. Phylogenetic Analysis
3.3. Comparative Assessment of immunodominant Linear Epitopes of Capsid Protein
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pépin, M.; Vitu, C.; Russo, P.; Mornex, J.F.; Peterhans, E. Maedi-Visna Virus Infection in Sheep: A Review. Vet. Res. 1998, 29, 341–367. [Google Scholar] [PubMed]
- Minguijón, E.; Reina, R.; Pérez, M.; Polledo, L.; Villoria, M.; Ramírez, H.; Leginagoikoa, I.; Badiola, J.J.; García-Marín, J.F.; de Andrés, D.; et al. Small Ruminant Lentivirus Infections and Diseases. Vet. Microbiol. 2015, 181, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Peterhans, E.; Greenland, T.; Badiola, J.; Harkiss, G.; Bertoni, G.; Amorena, B.; Eliaszewicz, M.; Juste, R.A.; Kraßnig, R.; Lafont, J.-P.; et al. Routes of Transmission and Consequences of Small Ruminant Lentiviruses (SRLVs) Infection and Eradication Schemes. Vet. Res. 2004, 35, 257–274. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Stavropoulos, I.; Chaintoutis, S.C.; Bossis, I.; Gelasakis, A.I. Serological, Molecular and Culture-based Diagnosis of Lentiviral Infections in Small Ruminants. Viruses 2021, 13, 1711. [Google Scholar] [CrossRef] [PubMed]
- L’Homme, Y.; Leboeuf, A.; Arsenault, J.; Fras, M. Identification and Characterization of an Emerging Small Ruminant Lentivirus Circulating Recombinant Form (CRF). Virology 2015, 475, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Santry, L.A.; de Jong, J.; Gold, A.C.; Walsh, S.R.; Menzies, P.I.; Wootton, S.K. Genetic Characterization of Small Ruminant Lentiviruses Circulating in Naturally Infected Sheep and Goats in Ontario, Canada. Virus Res. 2013, 175, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Quasso, A.; Profiti, M.; Lacerenza, D.; Muz, D.; Rosati, S. Genetic Characterization of Small Ruminant Lentivirus in Italian Mixed Flocks: Evidence for a Novel Genotype Circulating in a Local Goat Population. J. Gen. Virol. 2007, 88, 3423–3427. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.; Böni, J.; Huder, J.B.; Vogt, H.R.; Mühlherr, J.; Zanoni, R.; Miserez, R.; Lutz, H.; Schüpbach, J. Phylogenetic Analysis and Reclassification of Caprine and Ovine Lentiviruses Based on 104 New Isolates: Evidence for Regular Sheep-to-Goat Transmission and Worldwide Propagation through Livestock Trade. Virology 2004, 319, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Reina, R.; Mora, M.I.; Glaria, I.; García, I.; Solano, C.; Luján, L.; Badiola, J.J.; Contreras, A.; Berriatua, E.; Juste, R.; et al. Molecular Characterization and Phylogenetic Study of Maedi Visna and Caprine Arthritis Encephalitis Viral Sequences in Sheep and Goats from Spain. Virus Res. 2006, 121, 189–198. [Google Scholar] [CrossRef]
- Kalogianni, A.I.; Bossis, I.; Ekateriniadou, L.V.; Gelasakis, A.I. Etiology, Epizootiology and Control of Maedi-Visna in Dairy Sheep: A Review. Animals 2020, 10, 616. [Google Scholar] [CrossRef]
- Leroux, C.; Cruz, J.C.M.; Mornex, J.-F. SRLVs: A Genetic Continuum of Lentiviral Species in Sheep and Goats with Cumulative Evidence of Cross Species Transmission. Curr. HIV Res. 2010, 8, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Da Cruz, J.C.M.; Singh, D.K.; Lamara, A.; Chebloune, Y. Small Ruminant Lentiviruses (SRLVs) Break the Species Barrier to Acquire New Host Range. Viruses 2013, 5, 1867–1884. [Google Scholar] [CrossRef] [PubMed]
- Michiels, R.; Adjadj, N.R.; De Regge, N. Phylogenetic Analysis of Belgian Small Ruminant Lentiviruses Supports Cross Species Virus Transmission and Identifies New Subtype B5 Strains. Pathogens 2020, 9, 183. [Google Scholar] [CrossRef]
- Gjerset, B.; Rimstad, E.; Teige, J.; Soetaert, K.; Jonassen, C.M. Impact of Natural Sheep-Goat Transmission on Detection and Control of Small Ruminant Lentivirus Group C Infections. Vet. Microbiol. 2009, 135, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Gjerset, B.; Storset, A.K.; Rimstad, E. Genetic Diversity of Small-Ruminant Lentiviruses: Characterization of Norwegian Isolates of Caprine Arthritis Encephalitis Virus. J. Gen. Virol. 2006, 87, 573–580. [Google Scholar] [CrossRef]
- Rosati, S.; Mannelli, A.; Merlo, T.; Ponti, N. Characterization of the Immunodominant Cross-Reacting Epitope of Visna Maedi Virus and Caprine Arthritis-Encephalitis Virus Capsid Antigen. Virus Res. 1999, 61, 177–183. [Google Scholar] [CrossRef]
- Gomez-Lucia, E.; Barquero, N.; Domenech, A. Maedi-Visna Virus: Current Perspectives. Vet. Med. Res. Rep. 2018, 9, 11–21. [Google Scholar] [CrossRef]
- De Andrés, D.; Klein, D.; Watt, N.J.; Berriatua, E.; Torsteinsdottir, S.; Blacklaws, B.A.; Harkiss, G.D. Diagnostic Tests for Small Ruminant Lentiviruses. Vet. Microbiol. 2005, 107, 49–62. [Google Scholar] [CrossRef]
- Herrmann-Hoesing, L.M. Diagnostic Assays Used to Control Small Ruminant Lentiviruses. J. Vet. Diagn. Investig. 2010, 22, 843–855. [Google Scholar] [CrossRef]
- Karanikolaou, K.; Angelopoulou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Papadopoulos, O.; Koptopoulos, G. Detection of Small Ruminant Lentiviruses by PCR and Serology Tests in Field Samples of Animals from Greece. Small Rumin. Res. 2005, 58, 181–187. [Google Scholar] [CrossRef]
- Brellou, G.D.; Angelopoulou, K.; Poutahidis, T.; Vlemmas, I. Detection of Maedi-Visna Virus in the Liver and Heart of Naturally Infected Sheep. J. Comp. Pathol. 2007, 136, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Brellou, G.D.; Vlemmas, I. Detection of Maedi-Visna Virus in the Kidneys of Naturally Infected Sheep. J. Comp. Pathol. 2006, 134, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Kalogianni, A.I.; Bouzalas, I.; Bossis, I.; Gelasakis, A.I. Seroepidemiology of Maedi-Visna in Intensively Reared Dairy. Animals 2023, 13, 2273. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Karanikolaou, K.; Papanastasopoulou, M.; Koumpati-Artopiou, M.; Vlemmas, I.; Papadopoulos, O.; Koptopoulos, G. First Partial Characterisation of Small Ruminant Lentiviruses from Greece. Vet. Microbiol. 2005, 109, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Chassalevris, T.; Chaintoutis, S.C.; Apostolidi, E.D.; Giadinis, N.D.; Vlemmas, I.; Brellou, G.D.; Dovas, C.I. A Highly Sensitive Semi-Nested Real-Time PCR Utilizing Oligospermine-Conjugated Degenerate Primers for the Detection of Diverse Strains of Small Ruminant Lentiviruses. Mol. Cell. Probes 2020, 51, 101528. [Google Scholar] [CrossRef] [PubMed]
- Eltahir, Y.M.; Dovas, C.I.; Papanastassopoulou, M.; Koumbati, M.; Giadinis, N.; Verghese-Nikolakaki, S.; Koptopoulos, G. Development of a Semi-Nested PCR Using Degenerate Primers for the Generic Detection of Small Ruminant Lentivirus Proviral DNA. J. Virol. Methods 2006, 135, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Kalogianni, A.I.; Bouzalas, I.; Bossis, I.; Gelasakis, A.I. A Longitudinal Cohort Study of Risk Factors Associated with Small Ruminant Lentivirus Seropositivity in Intensively Reared Dairy Ewes in Greece. Pathogens 2023, 12, 1200. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Crandall, K.A. Phylogeny Estimation and Hypothesis Testing Using Maximum Likelihood. Annu. Rev. Ecol. Syst. 1997, 28, 437–466. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 9780199881222. [Google Scholar]
- Olech, M.; Rachid, A.; Croisé, B.; Kuźmak, J.; Valas, S. Genetic and Antigenic Characterization of Small Ruminant Lentiviruses Circulating in Poland. Virus Res. 2012, 163, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Hodor, D.; Toma, C.; Negoescu, A.; Taulescu, M. First Molecular Characterization of Small Ruminant Lentiviruses Detected in Romania. Animals 2023, 13, 3718. [Google Scholar] [CrossRef] [PubMed]
- Fras, M.; Leboeuf, A.; Labrie, F.M.; Laurin, M.A.; Singh Sohal, J.; L’Homme, Y. Phylogenetic Analysis of Small Ruminant Lentiviruses in Mixed Flocks: Multiple Evidence of Dual Infection and Natural Transmission of Types A2 and B1 between Sheep and Goats. Infect. Genet. Evol. 2013, 19, 97–104. [Google Scholar] [CrossRef]
- Ramírez, H.; Glaria, I.; de Andrés, X.; Martínez, H.; Hernández, M.; Reina, R.; Iráizoz, E.; Crespo, H.; Berriatua, E.; Vázquez, J.; et al. Recombinant Small Ruminant Lentivirus Subtype B1 in Goats and Sheep of Imported Breeds in Mexico. Vet. J. 2011, 190, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, W.P.S.; Tórtora, J.L.; Martínez, H.A.; García, M.M.; Cuevas-Romero, S.; Cerriteño, J.L.; Ramírez, H. Genotyping Based on the LTR Region of Small Ruminant Lentiviruses from Naturally Infected Sheep and Goats from Mexico. BioMed Res. Int. 2019, 2019, 4279573. [Google Scholar] [CrossRef]
- Martín-Sánchez, A.M.; Ciro-Gómez, G.; Sayas, E.; Vilella-Esplá, J.; Ben-Abda, J.; Pérez-Alvarez, J.Á. Date Palm By-Products as a New Ingredient for the Meat Industry: Application to Pork Liver Pâté. MESC 2013, 93, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Olech, M.; Kuźmak, J. Molecular Characterization of Small Ruminant Lentiviruses in Polish Mixed Flocks Supports Evidence of Cross Species Transmission, Dual Infection, a Recombination Event, and Reveals the Existence of New Subtypes within Group A. Viruses 2021, 13, 2529. [Google Scholar] [CrossRef]
- Barquero, N.; Arjona, A.; Domenech, A.; Toural, C.; De Las Heras, A.; Fernández-Garayzabal, J.F.; Ruiz-Santa Quiteria, J.A.; Gomez-Lucia, E. Paper: Diagnostic Performance of PCR and ELISA on Blood and Milk Samples and Serological Survey for Small Ruminant Lentiviruses in Central Spain. Vet. Rec. 2011, 168, 20. [Google Scholar] [CrossRef]
- De Regge, N.; Cay, B. Development, Validation and Evaluation of Added Diagnostic Value of a q(RT)-PCR for the Detection of Genotype A Strains of Small Ruminant Lentiviruses. J. Virol. Methods 2013, 194, 250–257. [Google Scholar] [CrossRef]
- Barquero, N.; Domenech, A.; Arjona, A.; Fernández-Garayzabal, J.F.; Ruiz-Santa-Quiteria, J.A.; Gomez-Lucia, E. Comparison of Two PCR and One ELISA Techniques for the Detection of Small Ruminant Lentiviruses (SRLVs) in Milk of Sheep and Goats. Res. Vet. Sci. 2013, 94, 817–819. [Google Scholar] [CrossRef] [PubMed]
- Dolfini, T.; Conrad, L.F.; Flores, I.V.C.; Ravazzolo, A.P. Comparison of Primer Pairs: Greater Degeneracy Improves Small Ruminant Lentivirus (SRLVs) Detection by Seminested PCR. Small Rumin. Res. 2015, 123, 189–192. [Google Scholar] [CrossRef]
- Zanoni, R.G.; Nauta, I.M.; Kuhnert, P.; Pauli, U.; Pohl, B.; Peterhans, E. Genomic Heterogeneity of Small Ruminant Lentiviruses Detected by PCR. Vet. Microbiol. 1992, 33, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, K.; Brellou, G.D.; Greenland, T.; Vlemmas, I. A Novel Deletion in the LTR Region of a Greek Small Ruminant Lentivirus May Be Associated with Low Pathogenicity. Virus Res. 2006, 118, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Grego, E.; Bertolotti, L.; Carrozza, M.L.; Profiti, M.; Mazzei, M.; Tolari, F.; Rosati, S. Genetic and Antigenic Characterization of the Matrix Protein of Two Genetically Distinct Ovine Lentiviruses. Vet. Microbiol. 2005, 106, 179–185. [Google Scholar] [CrossRef]
- Glaria, I.; Reina, R.; Crespo, H.; de Andrés, X.; Ramírez, H.; Biescas, E.; Pérez, M.M.; Badiola, J.; Luján, L.; Amorena, B.; et al. Phylogenetic Analysis of SRLV Sequences from an Arthritic Sheep Outbreak Demonstrates the Introduction of CAEV-like Viruses among Spanish Sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Germain, K.; Valas, S. Distribution and Heterogeneity of Small Ruminant Lentivirus Envelope Subtypes in Naturally Infected French Sheep. Virus Res. 2006, 120, 156–162. [Google Scholar] [CrossRef]
- Molaee, V.; Bazzucchi, M.; De Mia, G.M.; Otarod, V.; Abdollahi, D.; Rosati, S.; Lühken, G. Phylogenetic Analysis of Small Ruminant Lentiviruses in Germany and Iran Suggests Their Expansion with Domestic Sheep. Sci. Rep. 2020, 10, 2243. [Google Scholar] [CrossRef]
- Carrozza, M.L.; Niewiadomska, A.M.; Mazzei, M.; Abi-Said, M.R.; Hué, S.; Hughes, J.; Gatseva, A.; Gifford, R.J. Emergence and Pandemic Spread of Small Ruminant Lentiviruses. Virus Evol. 2023, 9, vead005. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, L.; Shi, X.; Duan, X.; Li, H.; Liu, S. Next-Generation Sequencing for the Genetic Characterization of Maedi/Visna Virus Isolated from the Northwest of China. J. Vet. Sci. 2021, 22, e66. [Google Scholar] [CrossRef]
- Laamanen, I.; Jakava-Viljanen, M.; Sihvonen, L. Genetic Characterization of Maedi-Visna Virus (MVV) Detected in Finland. Vet. Microbiol. 2007, 122, 357–365. [Google Scholar] [CrossRef]
- Grego, E.; Profiti, M.; Giammarioli, M.; Giannino, L.; Rutili, D.; Woodall, C.; Rosati, S. Genetic Heterogeneity of Small Ruminant Lentiviruses Involves Immunodominant Epitope of Capsid Antigen and Affects Sensitivity of Single-Strain-Based Immunoassay. Clin. Diagn. Lab. Immunol. 2002, 9, 828–832. [Google Scholar] [CrossRef]
- L’Homme, Y.; Ouardani, M.; Lévesque, V.; Bertoni, G.; Simard, C.; Pisoni, G. Molecular Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Isolated from Canadian Sheep and Goats. Virol. J. 2011, 8, 271. [Google Scholar] [CrossRef] [PubMed]
- Lacerenza, D.; Giammarioli, M.; Grego, E.; Marini, C.; Profiti, M.; Rutili, D.; Rosati, S. Antibody Response in Sheep Experimentally Infected with Different Small Ruminant Lentivirus Genotypes. Vet. Immunol. Immunopathol. 2006, 112, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Reina, R.; Grego, E.; Profiti, M.; Glaria, I.; Robino, P.; Quasso, A.; Amorena, B.; Rosati, S. Development of Specific Diagnostic Test for Small Ruminant Lentivirus Genotype E. Vet. Microbiol. 2009, 138, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Olech, M. The Genetic Variability of Small-Ruminant Lentiviruses and Its Impact on Tropism, the Development of Diagnostic Tests and Vaccines and the Effectiveness of Control Programmes. J. Vet. Res. 2023, 67, 479–502. [Google Scholar] [CrossRef] [PubMed]
- Nogarol, C.; Bertolotti, L.; Klevar, S.; Profiti, M.; Gjerset, B.; Rosati, S. Serological Characterization of Small Ruminant Lentiviruses: A Complete Tool for Serotyping Lentivirus Infection in Goat. Small Rumin. Res. 2019, 176, 42–46. [Google Scholar] [CrossRef]
- Acevedo Jiménez, G.E.; Tórtora Pérez, J.L.; Rodríguez Murillo, C.; Arellano Reynoso, B.; Ramírez Álvarez, H. Serotyping versus Genotyping in Infected Sheep and Goats with Small Ruminant Lentiviruses. Vet. Microbiol. 2021, 252, 108931. [Google Scholar] [CrossRef] [PubMed]
- Arcangeli, C.; Torricelli, M.; Sebastiani, C.; Lucarelli, D.; Ciullo, M.; Passamonti, F.; Giammarioli, M.; Biagetti, M. Genetic Characterization of Small Ruminant Lentiviruses (SRLVs) Circulating in Naturally Infected Sheep in Central Italy. Viruses 2022, 14, 686. [Google Scholar] [CrossRef]
- Gayo, E.; Cuteri, V.; Preziuso, S. Genetic Characterization and Phylogenetic Analysis of Small Ruminant Lentiviruses Detected in Spanish Assaf Sheep with Different Mammary Lesions. Viruses 2018, 10, 315. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Sample ID | Breed | Age (Years) | Serological Status | Farm | Location | Accession Number | Genotype |
|---|---|---|---|---|---|---|---|
| s-16sx | Lacaune | 3.5 | + | A | Central Greece | OR283228 | B |
| s-40sx | Lacaune | 2.0 | − | A | OR283225 | B | |
| s-56sx | Chios | 7.0 | + | A | OR283223 | A | |
| s-11s | Chios | 2.5 | + | B | Epirus | OR283218 | A |
| s-14s | Chios | 7.0 | + | B | OR283221 | A | |
| s-35s | Chios | 1.5 | − | B | OR283220 | A | |
| s-83s | Chios | 7.0 | + | B | OR283224 | A | |
| s-10fl | Chios | 4.0 | + | C | Peloponnese | OR283217 | A |
| s-21fl | Chios | 3.0 | − | C | OR283219 | A | |
| s-42fl | Chios | 3.0 | − | C | OR283222 | A | |
| s-8m | Lacaune | 3.0 | − | D | Western Greece | OR283226 | B |
| s-98m | Lacaune | 3.0 | + | D | OR283227 | B |
| s-16sx | s-40sx | s-56sx | s-11s | s-14s | s-35s | s-83s | s-10fl | s-21fl | s-42fl | s-8m | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| s-40sx | 3.9 | - | - | - | - | - | - | - | - | - | - |
| 2.2 | - | - | - | - | - | - | - | - | - | - | |
| s-56sx | 26.8 | 27.3 | - | - | - | - | - | - | - | - | - |
| 13.9 | 15.2 | - | - | - | - | - | - | - | - | - | |
| s-11s | 24.2 | 24.9 | 6.8 | - | - | - | - | - | - | - | - |
| 14.6 | 15.8 | 3.8 | - | - | - | - | - | - | - | - | |
| s-14s | 24.7 | 25.2 | 5.4 | 4.7 | - | - | - | - | - | - | - |
| 13.0 | 14.2 | 2.5 | 2.0 | - | - | - | - | - | - | - | |
| s-35s | 24.7 | 25.0 | 5.7 | 5.2 | 4.8 | - | - | - | - | - | - |
| 13.8 | 15.0 | 3.4 | 3.2 | 2.0 | - | - | - | - | - | - | |
| s-83s | 23.0 | 24.3 | 4.7 | 4.7 | 4.7 | 3.6 | - | - | - | - | - |
| 12.6 | 13.8 | 3.4 | 3.2 | 2.0 | 2.4 | - | - | - | - | - | |
| s-10fl | 25.4 | 26.1 | 6.5 | 5.1 | 4.1 | 4.7 | 5.1 | - | - | - | - |
| 14.2 | 15.4 | 3.8 | 2.8 | 2.4 | 2.4 | 3.6 | - | - | - | - | |
| s-21fl | 25.4 | 26.1 | 6.5 | 5.1 | 4.1 | 4.7 | 5.1 | - | - | - | - |
| 14.2 | 15.4 | 3.8 | 2.8 | 2.4 | 2.4 | 3.6 | - | - | - | - | |
| s-42fl | 28.9 | 29.2 | 11.9 | 11.6 | 10.5 | 11.2 | 11.4 | 11.1 | 11.1 | - | - |
| 15.9 | 17.1 | 5.5 | 6.1 | 4.9 | 5.3 | 6.5 | 6.1 | 6.1 | - | - | |
| s-8m | 4.9 | 5.6 | 25.2 | 23.0 | 24.4 | 23.9 | 22.6 | 24.3 | 24.3 | 26.5 | - |
| 2.4 | 2.8 | 14.3 | 14.6 | 13.8 | 14.6 | 13.4 | 15.0 | 15.0 | 15.9 | - | |
| s-98m | 5.6 | 6.3 | 25.6 | 23.2 | 24.4 | 23.8 | 22.3 | 24.4 | 24.4 | 26.4 | 1.2 |
| 2.4 | 2.8 | 14.3 | 14.3 | 13.5 | 14.3 | 13.5 | 14.7 | 14.7 | 15.9 | 0.0 |
| s-16sx | s-40sx | s-56sx | s-11s | s-14s | s-35s | s-83s | s-10fl | s-21fl | s-42fl | s-8m | s-98m | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| EV1 (A1/SC) | 27.9 | 29.3 | 24.2 | 21.2 | 22.4 | 21.7 | 21.9 | 22.8 | 22.8 | 24.7 | 26.5 | 25.8 |
| 18.7 | 19.9 | 12.2 | 13.3 | 12.9 | 13.3 | 11.2 | 14.1 | 14.1 | 15.5 | 19.1 | 18.4 | |
| K1514 (A1/USA) | 23.3 | 24.8 | 22.0 | 19.3 | 21.2 | 19.6 | 19.8 | 20.5 | 20.5 | 23.8 | 24.2 | 23.8 |
| 14.2 | 15.4 | 8.0 | 8.4 | 6.4 | 7.6 | 6.8 | 8.8 | 8.8 | 8.5 | 14.6 | 14.7 | |
| SA-OMVV (A1/SA) | 24.1 | 25.7 | 21.3 | 19.6 | 19.8 | 19.1 | 20.6 | 20.0 | 20.0 | 21.5 | 23.9 | 23.9 |
| 18.3 | 19.5 | 8.4 | 11.6 | 10.0 | 11.6 | 10.4 | 12.0 | 12.0 | 12.2 | 18.7 | 18.0 | |
| KV1772 (A1/IC) | 23.4 | 25.0 | 22.2 | 19.5 | 21.3 | 19.8 | 20.0 | 20.7 | 20.7 | 23.6 | 24.3 | 24.0 |
| 13.8 | 15.0 | 7.6 | 8.0 | 6.0 | 7.2 | 6.4 | 8.4 | 8.4 | 8.1 | 14.2 | 14.3 | |
| OvLv 85/34 (A2/USA) | 20.5 | 22.7 | 19.0 | 18.7 | 18.5 | 17.7 | 17.2 | 18.2 | 18.2 | 20.1 | 20.9 | 21.4 |
| 15.4 | 16.6 | 7.6 | 8.4 | 6.4 | 7.6 | 6.8 | 8.8 | 8.8 | 9.3 | 15.0 | 14.7 | |
| SRLV007 (A8/IT) | 27.3 | 28.9 | 20.7 | 18.6 | 19.8 | 18.8 | 18.1 | 19.6 | 19.6 | 21.0 | 26.2 | 26.2 |
| 17.4 | 18.6 | 10.5 | 12.8 | 11.6 | 12.8 | 12.0 | 12.0 | 12.0 | 13.8 | 18.2 | 17.6 | |
| SRLV009 (A18/IT) | 26.8 | 29.6 | 22.7 | 20.4 | 21.1 | 20.4 | 20.9 | 21.3 | 21.3 | 23.7 | 26.3 | 24.8 |
| 19.0 | 20.2 | 11.8 | 14.4 | 13.2 | 14.0 | 13.6 | 14.8 | 14.8 | 15.9 | 19.4 | 18.8 | |
| SRLV038 (A19/IT) | 25.0 | 26.9 | 20.1 | 18.4 | 18.7 | 17.7 | 19.1 | 17.6 | 17.6 | 19.5 | 24.5 | 24.4 |
| 17.4 | 18.6 | 7.1 | 10.0 | 9.2 | 10.0 | 9.6 | 10.0 | 10.0 | 11.0 | 17.8 | 17.1 | |
| It-561 (A19/IT) | 27.4 | 27.5 | 20.7 | 19.8 | 20.2 | 20.8 | 20.6 | 19.5 | 19.5 | 20.3 | 24.9 | 25.3 |
| 18.2 | 19.4 | 9.7 | 11.5 | 10.7 | 11.5 | 11.1 | 11.5 | 11.5 | 11.5 | 19.0 | 19.0 | |
| sh248 (A22/LB) | 28.3 | 28.7 | 24.2 | 23.5 | 21.9 | 21.5 | 22.4 | 21.7 | 21.7 | 24.0 | 27.4 | 28.5 |
| 15.8 | 17.0 | 11.3 | 11.6 | 10.0 | 11.6 | 10.0 | 11.2 | 11.2 | 12.6 | 17.0 | 17.1 | |
| sh173 (A22/J) | 30.0 | 30.4 | 26.2 | 24.8 | 23.4 | 22.8 | 24.9 | 23.7 | 23.7 | 25.1 | 28.6 | 28.1 |
| 15.8 | 17.0 | 10.5 | 10.8 | 9.6 | 11.2 | 11.2 | 11.2 | 11.2 | 11.0 | 16.6 | 16.3 | |
| SRLV-Greece-S1 (A/GR) | 22.4 | 24.2 | 12.6 | 10.0 | 11.5 | 10.0 | 9.6 | 10.4 | 10.4 | 11.1 | 23.1 | 22.1 |
| 10.6 | 12.2 | 2.2 | 1.7 | 0.6 | 2.2 | 1.1 | 2.8 | 2.8 | 3.3 | 11.1 | 11.1 | |
| SRLV-Greece-S2 (A/GR) | 22.9 | 25.5 | 12.6 | 10.0 | 11.0 | 10.4 | 9.1 | 10.4 | 10.4 | 11.3 | 23.4 | 22.4 |
| 11.7 | 13.3 | 3.9 | 3.3 | 2.2 | 2.8 | 2.8 | 3.9 | 3.9 | 3.9 | 12.2 | 12.2 | |
| CAEV-Co (B1/USA) | 15.1 | 15.0 | 27.2 | 25.2 | 23.9 | 26.1 | 24.8 | 24.9 | 24.9 | 26.0 | 13.2 | 13.1 |
| 8.4 | 9.2 | 14.3 | 15.3 | 13.7 | 14.1 | 14.1 | 14.5 | 14.5 | 15.5 | 7.2 | 6.5 | |
| GANSU (B1/CH) | 15.1 | 15.4 | 26.9 | 24.1 | 23.7 | 24.4 | 24.2 | 23.9 | 23.9 | 27.9 | 14.1 | 14.5 |
| 10.0 | 10.8 | 17.3 | 16.9 | 15.3 | 14.5 | 15.7 | 16.1 | 16.1 | 17.6 | 9.6 | 9.3 | |
| Ov496 (B2/SP) | 9.9 | 9.6 | 26.8 | 24.2 | 24.6 | 24.9 | 24.2 | 25.2 | 25.2 | 28.5 | 8.7 | 9.5 |
| 2.8 | 3.2 | 15.2 | 16.1 | 14.5 | 15.3 | 14.1 | 15.7 | 15.7 | 16.7 | 2.0 | 2.0 | |
| SRLV042 (B2/IT) | 7.4 | 8.5 | 28.1 | 24.9 | 25.7 | 25.4 | 24.5 | 25.9 | 25.9 | 28.3 | 6.2 | 6.9 |
| 10.1 | 10.5 | 17.4 | 19.4 | 19.0 | 20.2 | 18.5 | 19.8 | 19.8 | 21.7 | 9.3 | 8.5 | |
| It-007 (B2/IT) | 7.7 | 8.6 | 28.1 | 25.2 | 25.1 | 25.4 | 24.6 | 26.4 | 26.4 | 29.4 | 6.4 | 7.2 |
| 10.4 | 10.8 | 16.9 | 19.3 | 18.5 | 19.7 | 18.1 | 19.3 | 19.3 | 21.2 | 9.6 | 8.9 | |
| It-Pi1 (B2/IT) | 8.3 | 8.9 | 27.5 | 26.0 | 25.8 | 26.1 | 25.2 | 26.1 | 26.1 | 29.3 | 6.3 | 7.1 |
| 15.7 | 16.1 | 20.3 | 22.9 | 22.1 | 23.3 | 21.7 | 22.9 | 22.9 | 24.9 | 14.9 | 14.2 | |
| SRLV001 (B2/IT) | 8.8 | 8.6 | 27.6 | 25.0 | 26.5 | 26.6 | 25.3 | 27.3 | 27.3 | 29.2 | 6.9 | 7.2 |
| 10.8 | 11.2 | 17.7 | 19.7 | 19.3 | 20.5 | 18.9 | 20.1 | 20.1 | 21.2 | 10.0 | 9.3 | |
| Volterra (B3/IT) | 23.9 | 26.2 | 26.4 | 26.5 | 27.7 | 26.3 | 26.7 | 26.2 | 26.2 | 28.8 | 22.8 | 23.3 |
| 14.6 | 15.4 | 17.9 | 20.3 | 19.5 | 20.7 | 19.1 | 20.3 | 20.3 | 21.5 | 14.6 | 13.9 | |
| TR-DM (B3/TR) | 25.2 | 25.7 | 27.9 | 28.6 | 27.4 | 26.9 | 28.5 | 28.5 | 28.5 | 31.6 | 23.9 | 23.9 |
| 14.1 | 14.9 | 16.5 | 18.5 | 17.7 | 18.9 | 18.1 | 18.5 | 18.5 | 20.4 | 13.7 | 13.0 | |
| 1GA (C/NW) | 25.9 | 25.7 | 32.0 | 32.2 | 30.4 | 31.3 | 30.4 | 31.5 | 31.5 | 31.6 | 23.5 | 24.2 |
| 20.6 | 21.9 | 23.3 | 25.0 | 23.8 | 23.8 | 23.8 | 24.2 | 24.2 | 25.4 | 20.6 | 20.0 | |
| Roccaverano (E1/IT) | 35.5 | 37.8 | 41.4 | 39.8 | 39.9 | 38.7 | 37.9 | 38.4 | 38.4 | 40.1 | 36.4 | 37.3 |
| 27.2 | 28.5 | 26.7 | 28.6 | 27.8 | 28.2 | 27.4 | 28.2 | 28.2 | 28.3 | 27.6 | 27.0 | |
| Seui (E2/IT) | 38.3 | 40.4 | 40.8 | 37.4 | 36.7 | 37.1 | 36.1 | 36.4 | 36.4 | 40.3 | 38.6 | 39.3 |
| 27.2 | 28.5 | 27.5 | 29.4 | 28.2 | 29.4 | 27.8 | 29.0 | 29.0 | 29.9 | 28.0 | 27.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalogianni, A.I.; Bouzalas, I.; Marka, S.; Zografaki, M.-E.; Mavrikou, S.; Gelasakis, A.I. Genetic Characterization of Small Ruminant Lentiviruses Isolated from Dairy Sheep in Greece. Viruses 2024, 16, 547. https://doi.org/10.3390/v16040547
Kalogianni AI, Bouzalas I, Marka S, Zografaki M-E, Mavrikou S, Gelasakis AI. Genetic Characterization of Small Ruminant Lentiviruses Isolated from Dairy Sheep in Greece. Viruses. 2024; 16(4):547. https://doi.org/10.3390/v16040547
Chicago/Turabian StyleKalogianni, Aphrodite I., Ilias Bouzalas, Sofia Marka, Maria-Eleftheria Zografaki, Sofia Mavrikou, and Athanasios I. Gelasakis. 2024. "Genetic Characterization of Small Ruminant Lentiviruses Isolated from Dairy Sheep in Greece" Viruses 16, no. 4: 547. https://doi.org/10.3390/v16040547
APA StyleKalogianni, A. I., Bouzalas, I., Marka, S., Zografaki, M.-E., Mavrikou, S., & Gelasakis, A. I. (2024). Genetic Characterization of Small Ruminant Lentiviruses Isolated from Dairy Sheep in Greece. Viruses, 16(4), 547. https://doi.org/10.3390/v16040547