

Cardiometabolic Differences in People Living with HIV Receiving Integrase Strand Transfer Inhibitors Compared to Non-nucleoside Reverse Transcriptase Inhibitors: Implications for Current ART Strategies
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Data Collection
2.2. Ethics
2.3. Untargeted Metabolomics
2.4. Targeted Lipoproteomics
2.5. Statistics
3. Results
3.1. Participant Selection and Baseline Characteristics
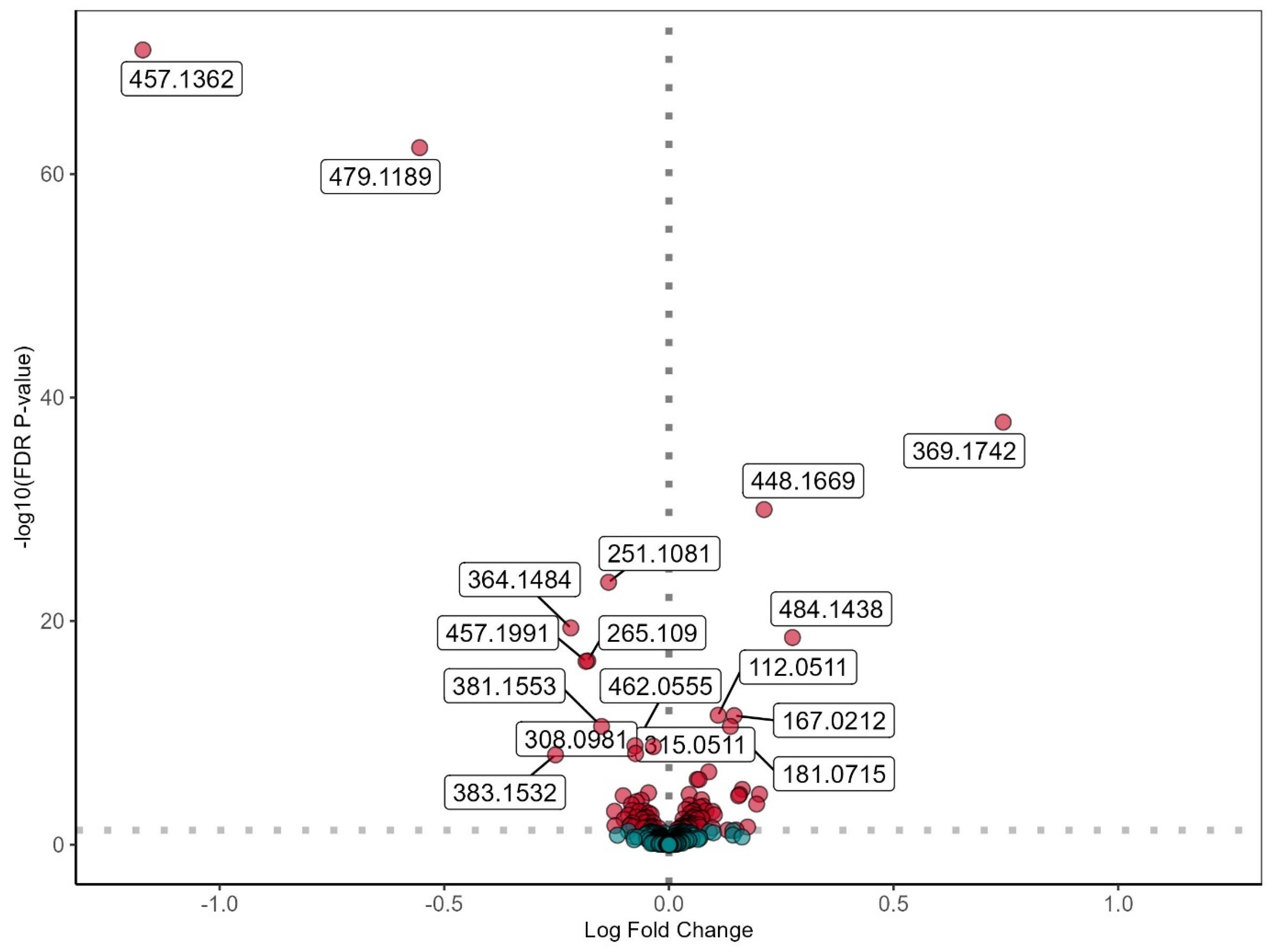
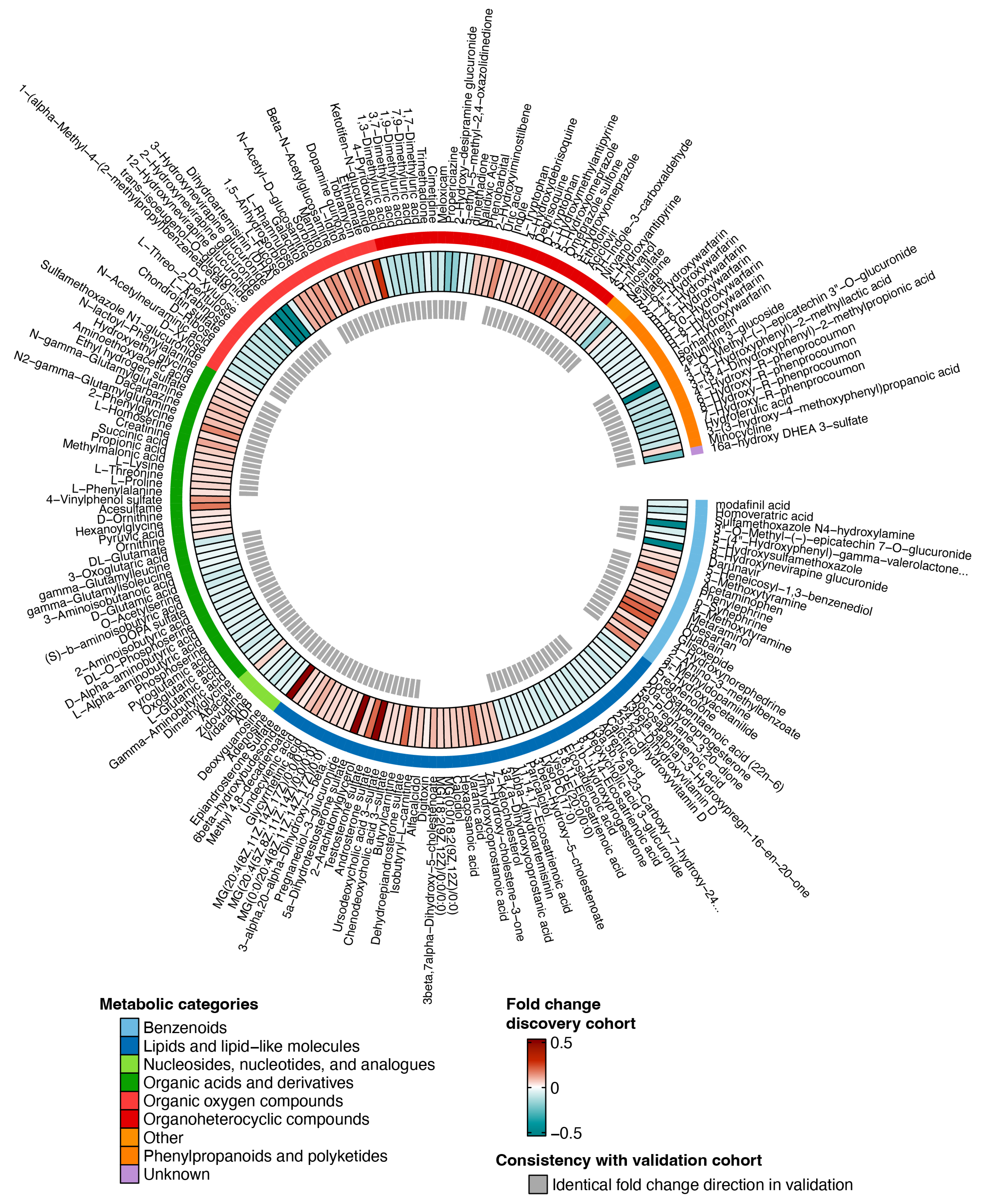
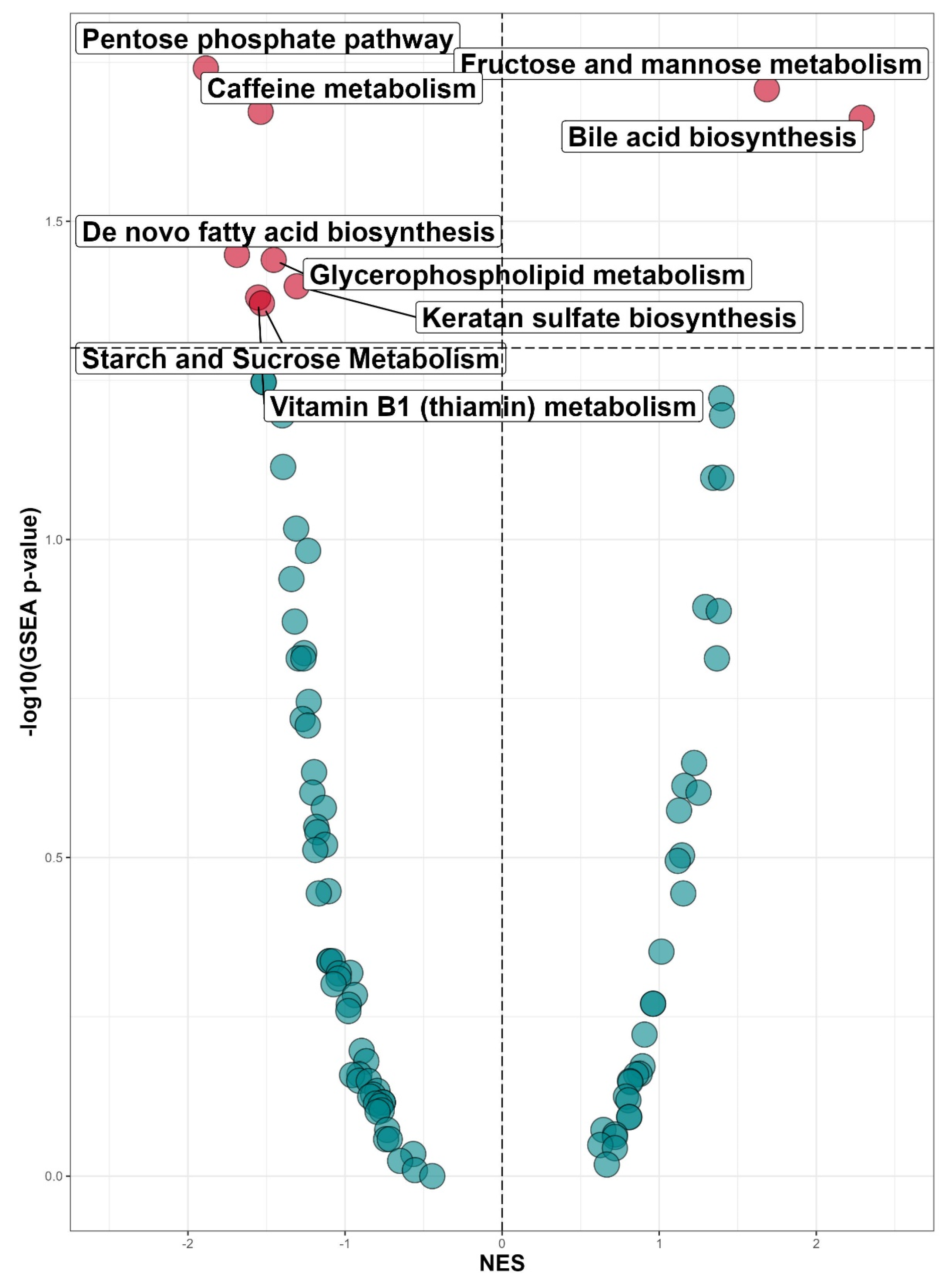
3.2. Untargeted Metabolomic Profiling in INSTI and NNRTI Users
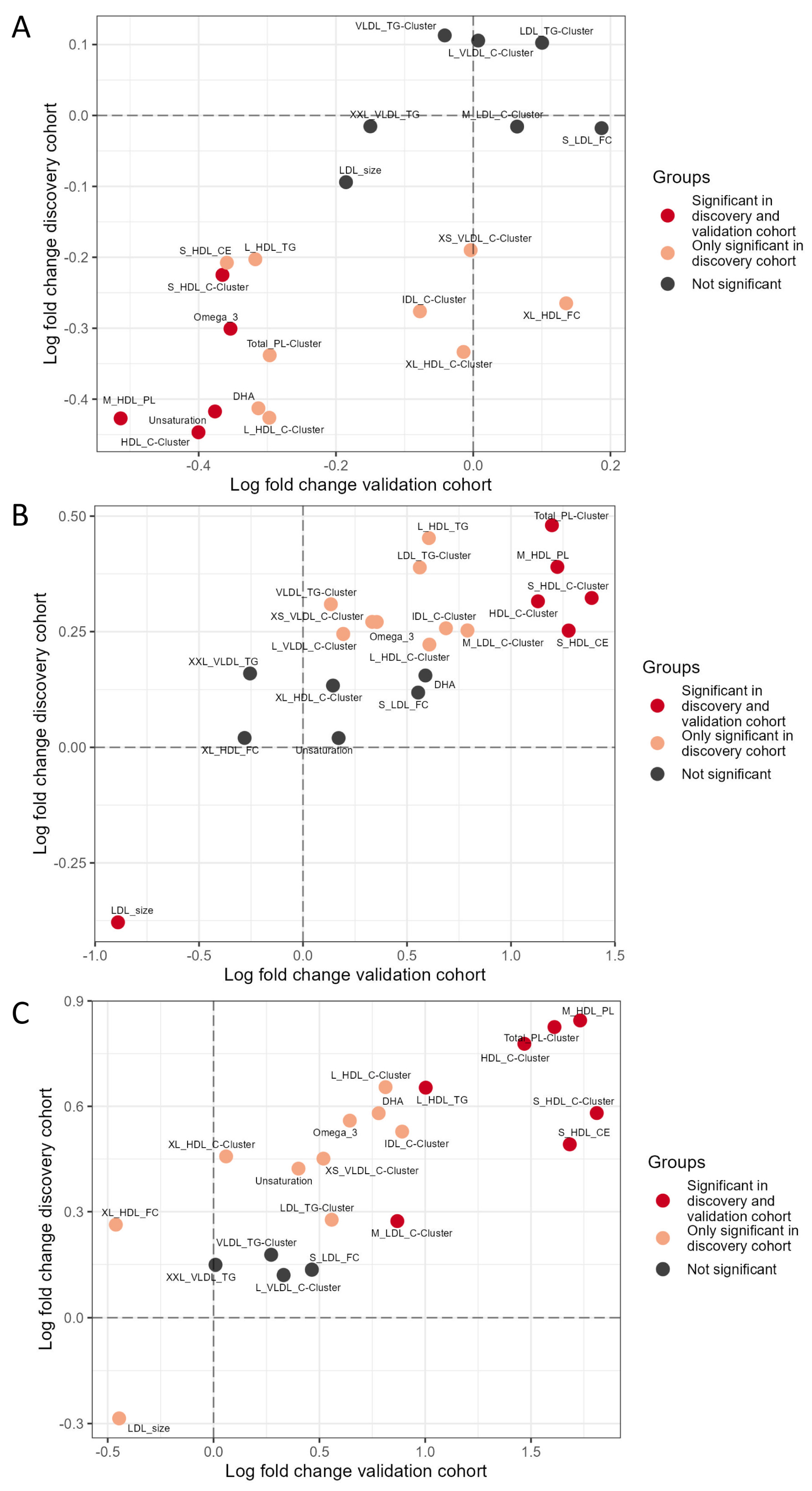
3.3. Targeted Lipoproteomic Profiling in INSTI and NNRTI Users
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zicari, S.; Sessa, L.; Cotugno, N.; Ruggiero, A.; Morrocchi, E.; Concato, C.; Rocca, S.; Zangari, P.; Manno, E.C.; Palma, P. Immune Activation, Inflammation, and Non-AIDS Co-Morbidities in HIV-Infected Patients under Long-Term ART. Viruses 2019, 11, 200. [Google Scholar] [CrossRef] [PubMed]
- CDC. Guidelines for the Use of Antiretroviral Agents in Adults and Adolescents with HIV; CDC: Atlanta, GA, USA, 2022. [Google Scholar]
- Sax, P.E.; Erlandson, K.M.; Lake, J.E.; McComsey, G.A.; Orkin, C.; Esser, S.; Brown, T.T.; Rockstroh, J.K.; Wei, X.; Carter, C.C.; et al. Weight Gain Following Initiation of Antiretroviral Therapy: Risk Factors in Randomized Comparative Clinical Trials. Clin. Infect. Dis. 2020, 71, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Norwood, J.; Turner, M.; Bofill, C.; Rebeiro, P.; Shepherd, B.; Bebawy, S.; Hulgan, T.; Raffanti, S.; Haas, D.W.; Sterling, T.R.; et al. Brief Report: Weight Gain in Persons with HIV Switched from Efavirenz-Based to Integrase Strand Transfer Inhibitor-Based Regimens. J. Acquir. Immune Defic. Syndr. 2017, 76, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Hill, A. Risks of metabolic syndrome and diabetes with integrase inhibitor-based therapy: Republication. Curr. Opin. HIV AIDS 2021, 16, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Neesgaard, B.; Greenberg, L.; Miro, J.M.; Grabmeier-Pfistershammer, K.; Wandeler, G.; Smith, C.; De Wit, S.; Wit, F.; Pelchen-Matthews, A.; Mussini, C.; et al. Associations between integrase strand-transfer inhibitors and cardiovascular disease in people living with HIV: A multicentre prospective study from the RESPOND cohort consortium. Lancet HIV 2022, 9, e474–e485. [Google Scholar] [CrossRef] [PubMed]
- Wood, B.R.; Huhn, G.D. Excess Weight Gain with Integrase Inhibitors and Tenofovir Alafenamide: What Is the Mechanism and Does It Matter? Open Forum Infect. Dis. 2021, 8, ofab542. [Google Scholar] [CrossRef] [PubMed]
- Lake, J.E.; Trevillyan, J. Impact of Integrase inhibitors and tenofovir alafenamide on weight gain in people with HIV. Curr. Opin. HIV AIDS 2021, 16, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Jacob, M.; Lopata, A.L.; Dasouki, M.; Abdel Rahman, A.M. Metabolomics toward personalized medicine. Mass. Spectrom. Rev. 2019, 38, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Kishton, R.J.; Rathmell, J. A guide to immunometabolism for immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef]
- Saez-Cirion, A.; Sereti, I. Immunometabolism and HIV-1 pathogenesis: Food for thought. Nat. Rev. Immunol. 2021, 21, 5–19. [Google Scholar] [PubMed]
- Munshi, S.U.; Rewari, B.B.; Bhavesh, N.S.; Jameel, S. Nuclear magnetic resonance based profiling of biofluids reveals metabolic dysregulation in HIV-infected persons and those on anti-retroviral therapy. PLoS ONE 2013, 8, e64298. [Google Scholar] [CrossRef] [PubMed]
- Deme, P.; Rubin, L.H.; Yu, D.; Xu, Y.; Nakigozi, G.; Nakasujja, N.; Anok, A.; Kisakye, A.; Quinn, T.C.; Reynolds, S.J.; et al. Immunometabolic Reprogramming in Response to HIV Infection Is Not Fully Normalized by Suppressive Antiretroviral Therapy. Viruses 2022, 14, 1313. [Google Scholar] [CrossRef] [PubMed]
- Vos, W.; Groenendijk, A.L.; Blaauw, M.J.T.; van Eekeren, L.E.; Navas, A.; Cleophas, M.C.P.; Vadaq, N.; Matzaraki, V.; Dos Santos, J.C.; Meeder, E.M.G.; et al. The 2000HIV study: Design, multi-omics methods and participant characteristics. Front. Immunol. 2022, 13, 982746. [Google Scholar] [CrossRef]
- Boender, T.S.; Smit, C.; Sighem, A.V.; Bezemer, D.; Ester, C.J.; Zaheri, S.; Wit, F.; Reiss, P. AIDS Therapy Evaluation in the Netherlands (ATHENA) national observational HIV cohort: Cohort profile. BMJ Open 2018, 8, e022516. [Google Scholar] [CrossRef]
- Fuhrer, T.; Heer, D.; Begemann, B.; Zamboni, N. High-throughput, accurate mass metabolome profiling of cellular extracts by flow injection-time-of-flight mass spectrometry. Anal. Chem. 2011, 83, 7074–7080. [Google Scholar] [CrossRef]
- Ter Horst, R.; van den Munckhof, I.C.L.; Schraa, K.; Aguirre-Gamboa, R.; Jaeger, M.; Smeekens, S.P.; Brand, T.; Lemmers, H.; Dijkstra, H.; Galesloot, T.E.; et al. Sex-Specific Regulation of Inflammation and Metabolic Syndrome in Obesity. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef] [PubMed]
- Shytaj, I.L.; Procopio, F.A.; Tarek, M.; Carlon-Andres, I.; Tang, H.Y.; Goldman, A.R.; Munshi, M.; Kumar Pal, V.; Forcato, M.; Sreeram, S.; et al. Glycolysis downregulation is a hallmark of HIV-1 latency and sensitizes infected cells to oxidative stress. EMBO Mol. Med. 2021, 13, e13901. [Google Scholar] [CrossRef]
- Crater, J.M.; Nixon, D.F.; Furler O’Brien, R.L. HIV-1 replication and latency are balanced by mTOR-driven cell metabolism. Front. Cell Infect. Microbiol. 2022, 12, 1068436. [Google Scholar] [CrossRef]
- Lahouassa, H.; Daddacha, W.; Hofmann, H.; Ayinde, D.; Logue, E.C.; Dragin, L.; Bloch, N.; Maudet, C.; Bertrand, M.; Gramberg, T.; et al. SAMHD1 restricts the replication of human immunodeficiency virus type 1 by depleting the intracellular pool of deoxynucleoside triphosphates. Nat. Immunol. 2012, 13, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Kochetov, G.A.; Solovjeva, O.N. Structure and functioning mechanism of transketolase. Biochim. Biophys. Acta 2014, 1844, 1608–1618. [Google Scholar] [CrossRef] [PubMed]
- Schenk, G.; Duggleby, R.G.; Nixon, P.F. Properties and functions of the thiamin diphosphate dependent enzyme transketolase. Int. J. Biochem. Cell Biol. 1998, 30, 1297–1318. [Google Scholar] [PubMed]
- Park, S.Y.; Faraci, G.; Nanda, S.; Ter-Saakyan, S.; Love, T.M.T.; Mack, W.J.; Dube, M.P.; Lee, H.Y. Gut microbiome in people living with HIV is associated with impaired thiamine and folate syntheses. Microb. Pathog. 2021, 160, 105209. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Keratan sulfate: An up-to-date review. Int. J. Biol. Macromol. 2015, 72, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Kot, M.; Daniel, W.A. Caffeine as a marker substrate for testing cytochrome P450 activity in human and rat. Pharmacol. Rep. 2008, 60, 789–797. [Google Scholar] [PubMed]
- von Moltke, L.L.; Greenblatt, D.J.; Granda, B.W.; Giancarlo, G.M.; Duan, S.X.; Daily, J.P.; Harmatz, J.S.; Shader, R.I. Inhibition of human cytochrome P450 isoforms by nonnucleoside reverse transcriptase inhibitors. J. Clin. Pharmacol. 2001, 41, 85–91. [Google Scholar] [CrossRef]
- Dobrinas, M.; Cornuz, J.; Oneda, B.; Kohler Serra, M.; Puhl, M.; Eap, C.B. Impact of smoking, smoking cessation, and genetic polymorphisms on CYP1A2 activity and inducibility. Clin. Pharmacol. Ther. 2011, 90, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wu, J.H.; Wang, Q.; Lemaitre, R.N.; Mukamal, K.J.; Djousse, L.; King, I.B.; Song, X.; Biggs, M.L.; Delaney, J.A.; et al. Prospective association of fatty acids in the de novo lipogenesis pathway with risk of type 2 diabetes: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2015, 101, 153–163. [Google Scholar] [CrossRef]
- Wu, J.H.; Lemaitre, R.N.; Imamura, F.; King, I.B.; Song, X.; Spiegelman, D.; Siscovick, D.S.; Mozaffarian, D. Fatty acids in the de novo lipogenesis pathway and risk of coronary heart disease: The Cardiovascular Health Study. Am. J. Clin. Nutr. 2011, 94, 431–438. [Google Scholar] [CrossRef]
- van Leth, F.; Phanuphak, P.; Stroes, E.; Gazzard, B.; Cahn, P.; Raffi, F.; Wood, R.; Bloch, M.; Katlama, C.; Kastelein, J.J.; et al. Nevirapine and efavirenz elicit different changes in lipid profiles in antiretroviral-therapy-naive patients infected with HIV-1. PLoS Med. 2004, 1, e19. [Google Scholar] [CrossRef] [PubMed]
- Maggi, P.; Bellacosa, C.; Carito, V.; Perilli, F.; Lillo, A.; Volpe, A.; Trillo, G.; Angiletta, D.; Regina, G.; Angarano, G. Cardiovascular risk factors in patients on long-term treatment with nevirapine- or efavirenz-based regimens. J. Antimicrob. Chemother. 2011, 66, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Rokx, C.; Verbon, A.; Rijnders, B.J. Short communication: Lipids and cardiovascular risk after switching HIV-1 patients on nevirapine and emtricitabine/tenofovir-DF to rilpivirine/emtricitabine/tenofovir-DF. AIDS Res. Hum. Retroviruses 2015, 31, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Yang, S.; Lu, J.; Wu, M. Small, Dense Low-Density Lipoprotein-Cholesterol and Atherosclerosis: Relationship and Therapeutic Strategies. Front. Cardiovasc. Med. 2021, 8, 804214. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Discovery Cohort | Validation Cohort | |||||
|---|---|---|---|---|---|---|
| INSTI | NNRTI | p-val | INSTI | NNRTI | p-val | |
| n = 730 | n = 617 | n = 209 | n = 90 | |||
| Age in years (IQR) | 51.0 (41.0–58.0) | 53.0 (46.0–60.0) | 0.0001 | 52.0 (45.0–61.0) | 54.0 (48.0–60.0) | 0.27 |
| Sex at birth (male) | 619 (84.8%) | 533 (86.4%) | 0.44 | 178 (85.2%) | 74 (82.2%) | 0.60 |
| BMI in kg/m2 Median (IQR) | 25.0 (22.7–27.7) | 24.7 (22.2–27.4) | 0.061 | 25.8 (23.0–28.4) | 25.1 (22.6–27.4) | 0.25 |
| Ethnicity (white) | 543 (74.4%) | 454 (73.6%) | 0.71 | 180 (86.1%) | 78 (86.7%) | 1.0 |
| Non-white | 185 (25.3%) | 163 (26.4%) | 29 (13.9%) | 12 (13.3%) | ||
| Missing | 2 (0.3%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | ||
| HIV duration in years Median (IQR) | 10.8 (5.7–16.5) | 14.6 (9.9–20.4) | <0.0001 | 7.0 (3.5–12.7) | 14.1 (10.3–19.4) | <0.0001 |
| cART Duration in years Median (IQR) | 8.3 (4.7–13.4) | 11.5 (8.3–17.9) | <0.0001 | 6.0 (3.3–10.6) | 11.2 (7.8–17.9) | <0.0001 |
| Latest CD4 count (×106 cells/L) Median (IQR) | 740.0 (550.5–940.0) | 698.5 (550.2–917.5) | 0.096 | 660.0 (480.0–810.0) | 685.0 (512.5–867.5) | 0.16 |
| CD4 Nadir (×106 cells/L) Median (IQR) | 280.0 (160.0–422.5) | 240.0 (150.0–350.0) | <0.0001 | 290.0 (170.0–452.5) | 280.0 (145.0–365.0) | 0.072 |
| Viral Load Zenith (copies/mL) Median (IQR) | 97,000.0 (36,650.5–248,154.0) | 100,000.0 (40,000.0–262,000.0) | 0.36 | 156,748.0 (39,240.8–346,862.5) | 200,000.0 (70,264.0–387,309.0) | 0.20 |
| Currently smoking | 238 (32.6%) | 167 (27.1%) | 0.013 | 59 (28.2%) | 31 (34.4%) | 0.40 |
| Missing | 56 (7.7%) | 35 (5.7%) | 23 (11.0%) | 7 (7.8%) | ||
| Packyears Median (IQR) | 6.0 (0.0–22.0) | 4.5 (0.0–21.5) | 0.73 | 6.0 (0.0–28.5) | 10.3 (0.0–34.0) | 0.33 |
| Had Non-AIDS malignancy | 28 (3.8%) | 30 (4.9%) | 0.42 | 12 (5.7%) | 3 (3.3%) | 0.57 |
| Had previous cardiovascular disease | 214 (29.3%) | 209 (33.9%) | 0.077 | 66 (31.6%) | 29 (32.2%) | 1.0 |
| On lipid lowering medication | 141 (19.3%) | 112 (18.2%) | 0.62 | 40 (19.1%) | 22 (24.4%) | 0.35 |
| INSTI in use: | ||||||
| Dolutegravir | 435 (60%) | 77 (37%) | ||||
| Bictegravir | 164 (22%) | 39 (19%) | ||||
| Elvitegravir | 112 (15% | 90 (43%) | ||||
| Raltegravir | 19 (3%) | 3 (1%) | ||||
| NNRTI in use: | ||||||
| Nevirapine | 236 (38%) | 38 (42%) | ||||
| Doravirine | 164 (27%) | 5 (6%) | ||||
| Rilpivirine | 133 (22%) | 28 (31%) | ||||
| Efavirenz | 84 (14%) | 19 (21%) | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vos, W.A.J.W.; Vadaq, N.; Matzaraki, V.; Otten, T.; Groenendijk, A.L.; Blaauw, M.J.T.; van Eekeren, L.E.; Brinkman, K.; de Mast, Q.; Riksen, N.P.; et al. Cardiometabolic Differences in People Living with HIV Receiving Integrase Strand Transfer Inhibitors Compared to Non-nucleoside Reverse Transcriptase Inhibitors: Implications for Current ART Strategies. Viruses 2024, 16, 582. https://doi.org/10.3390/v16040582
Vos WAJW, Vadaq N, Matzaraki V, Otten T, Groenendijk AL, Blaauw MJT, van Eekeren LE, Brinkman K, de Mast Q, Riksen NP, et al. Cardiometabolic Differences in People Living with HIV Receiving Integrase Strand Transfer Inhibitors Compared to Non-nucleoside Reverse Transcriptase Inhibitors: Implications for Current ART Strategies. Viruses. 2024; 16(4):582. https://doi.org/10.3390/v16040582
Chicago/Turabian StyleVos, Wilhelm A. J. W., Nadira Vadaq, Vasiliki Matzaraki, Twan Otten, Albert L. Groenendijk, Marc J. T. Blaauw, Louise E. van Eekeren, Kees Brinkman, Quirijn de Mast, Niels P. Riksen, and et al. 2024. "Cardiometabolic Differences in People Living with HIV Receiving Integrase Strand Transfer Inhibitors Compared to Non-nucleoside Reverse Transcriptase Inhibitors: Implications for Current ART Strategies" Viruses 16, no. 4: 582. https://doi.org/10.3390/v16040582