
Sosuga Virus Detected in Egyptian Rousette Bats (Rousettus aegyptiacus) in Sierra Leone
 , , , , ,
, , , , ,  , , , and add
Show full author list
, , , and add
Show full author list
Abstract
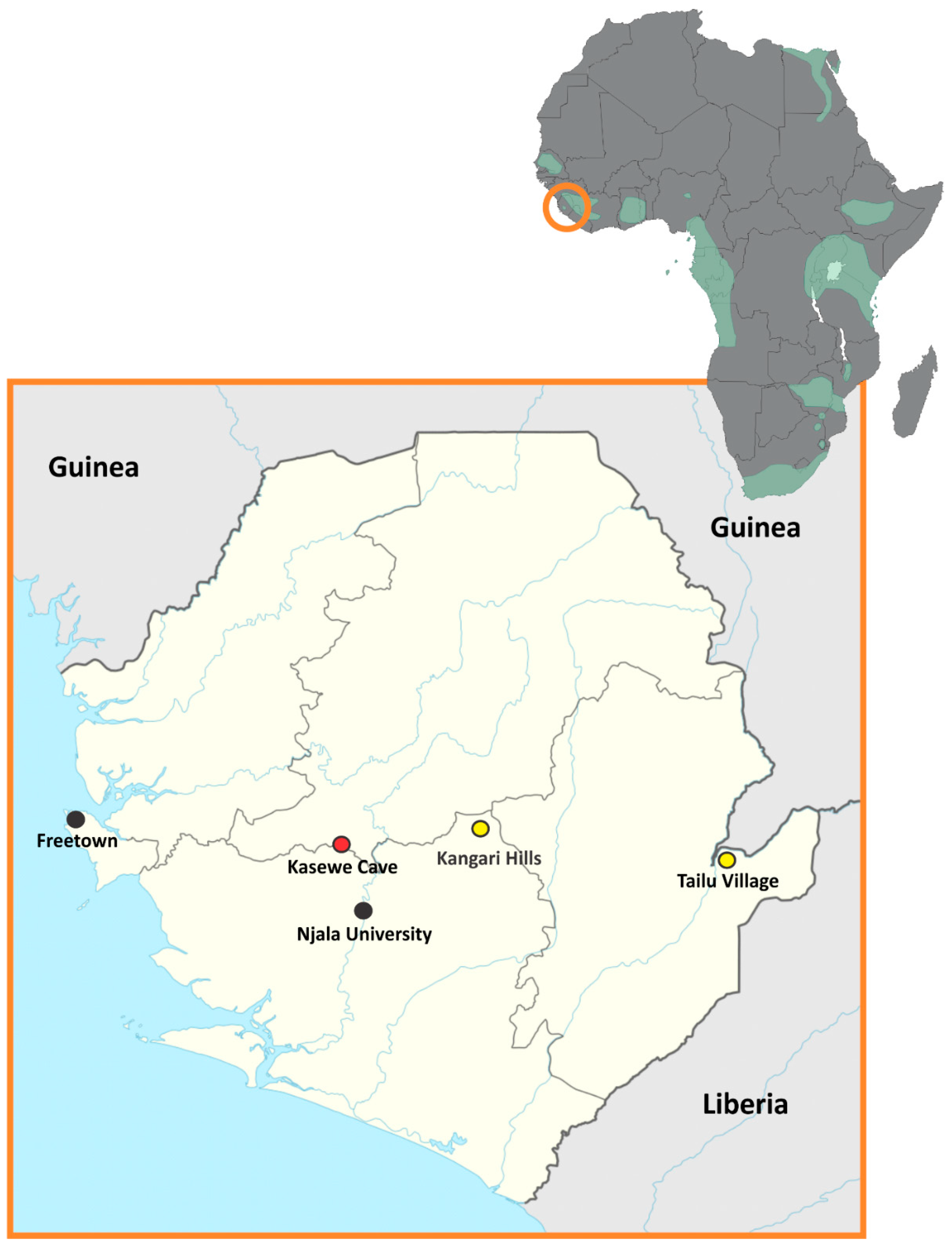
:1. Introduction
2. Materials and Methods
2.1. Bat Capture and Processing
2.2. Statistical Analyses
2.3. Sosuga Virus qRT-PCR
2.4. Serology
2.5. Virus Isolation
2.6. Sequencing
2.7. Phylogenetic Analysis
3. Results
3.1. Sosuga Virus qRT-PCR
3.2. Serology
3.3. Virus Isolation
3.4. Sequencing and Phylogenetic Analysis
4. Discussion
4.1. Sosuga Virus Circulation
4.2. Infection Bias
4.3. Public Health
4.4. Reservoir Status
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albarino, C.G.; Foltzer, M.; Towner, J.S.; Rowe, L.A.; Campbell, S.; Jaramillo, C.M.; Bird, B.H.; Reeder, D.M.; Vodzak, M.E.; Rota, P.; et al. Novel Paramyxovirus Associated with Severe Acute Febrile Disease, South Sudan and Uganda, 2012. Emerg. Infect. Dis. 2014, 20, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Amman, B.R.; Albarino, C.G.; Bird, B.H.; Nyakarahuka, L.; Sealy, T.K.; Balinandi, S.; Schuh, A.J.; Campbell, S.M.; Stroher, U.; Jones, M.E.; et al. A Recently Discovered Pathogenic Paramyxovirus, Sosuga Virus, is Present in Rousettus aegyptiacus Fruit Bats at Multiple Locations in Uganda. J. Wildl. Dis. 2015, 51, 774–779. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Corman, V.M.; Muller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Rasche, A.; Yordanov, S.; Seebens, A. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [PubMed]
- Pavri, K.M.; Singh, K.R.; Hollinger, F.B. Isolation of a new parainfluenza virus from a frugivorous bat, Rousettus leschenaulti, collected at Poona, India. Am. J. Trop. Med. Hyg. 1971, 20, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Henderson, G.W.; Laird, C.; Dermott, E.; Rima, B.K. Characterization of Mapuera virus: Structure, proteins and nucleotide sequence of the gene encoding the nucleocapsid protein. J. Gen. Virol. 1995, 76 Pt 10, 2509–2518. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.L.; Schountz, T.; Wang, L.F. Antiviral immune responses of bats: A review. Zoonoses Public Health 2013, 60, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Kurth, A.; Kohl, C.; Brinkmann, A.; Ebinger, A.; Harper, J.A.; Wang, L.F.; Muhldorfer, K.; Wibbelt, G. Novel paramyxoviruses in free-ranging European bats. PLoS ONE 2012, 7, e38688. [Google Scholar] [CrossRef] [PubMed]
- Pomeroy, L.W.; Bjornstad, O.N.; Holmes, E.C. The evolutionary and epidemiological dynamics of the paramyxoviridae. J. Mol. Evol. 2008, 66, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.B.; Chua, B.H.; Wang, C.W. Anthropogenic deforestation, El Niño and the emergence of Nipah virus in Malaysia. Malay. J. Pathol. 2002, 24, 15–21. [Google Scholar]
- Halpin, K.; Young, P.L.; Field, H.E.; Mackenzie, J.S. Isolation of Hendra virus from pteropid bats: A natural reservoir of Hendra virus. J. Gen. Virol. 2000, 81, 1927–1932. [Google Scholar] [CrossRef]
- Johara, M.Y.; Field, H.; Rashdi, A.M.; Morrissy, C.; van der Heide, B.; Rota, P.A.; bin Adzhar, A.; White, J.; Daniels, P.; Jamaluddin, A.; et al. Nipah virus infection in bats (order Chiroptera) in Peninsular Malaysia. Emerg. Infect. Dis. 2001, 7, 439–441. [Google Scholar]
- Young, P.L.; Halpin, K.; Selleck, P.W.; Field, H.; Gravel, J.L.; Kelly, M.A.; Mackenzie, J.S. Serologic evidence for the presence in Pteropus bats of a paramyxovirus related to equine morbillivirus. Emerg. Infect. Dis. 1996, 2, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Amman, B.R.; Schuh, A.J.; Sealy, T.K.; Spengler, J.R.; Welch, S.R.; Kirejczyk, S.G.; Albariño, C.G.; Nichol, S.T.; Towner, J.S. Experimental infection of Egyptian rousette bats (Rousettus aegyptiacus) with Sosuga virus demonstrates potential transmission routes for a bat-borne human pathogenic paramyxovirus. PLoS Neglected Trop. Dis. 2020, 14, e0008092. [Google Scholar] [CrossRef] [PubMed]
- Kirejczyk, S.G.; Amman, B.R.; Schuh, A.J.; Sealy, T.K.; Albariño, C.G.; Zhang, J.; Brown, C.C.; Towner, J.S. Histopathologic and Immunohistochemical Evaluation of Induced Lesions, Tissue Tropism and Host Responses following Experimental Infection of Egyptian Rousette Bats (Rousettus aegyptiacus) with the Zoonotic Paramyxovirus, Sosuga Virus. Viruses 2022, 14, 1278. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, N.H.G.; Du Plessis, E. Observations on the ecology and biology of the Cape fruit bat Rousettus aegyptiacus leachi in the Eastern Transvaal. S. Afr. J. Sci. 1976, 72, 270–273. [Google Scholar]
- Kingdon, J.; Happold, D.; Butynski, T.; Hoffmann, M.; Happold, M.; Kalina, J. Mammals of Africa; Bloomsbury Publishing: London, UK, 2013; Volume 1–6. [Google Scholar]
- Towner, J.S.; Amman, B.R.; Sealy, T.K.; Carroll, S.A.; Comer, J.A.; Kemp, A.; Swanepoel, R.; Paddock, C.D.; Balinandi, S.; Khristova, M.L.; et al. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009, 5, e1000536. [Google Scholar] [CrossRef]
- Amman, B.R.; Carroll, S.A.; Reed, Z.D.; Sealy, T.K.; Balinandi, S.; Swanepoel, R.; Kemp, A.; Erickson, B.R.; Comer, J.A.; Campbell, S.; et al. Seasonal pulses of Marburg virus circulation in juvenile Rousettus aegyptiacus bats coincide with periods of increased risk of human infection. PLoS Pathog. 2012, 8, e1002877. [Google Scholar] [CrossRef]
- Mutere, F.A. The breeding biology of the fruit bat Rousettus aegyptiacus E. Geoffroy living at o degrees 22′S. Acta Trop. 1968, 25, 97–108. [Google Scholar] [PubMed]
- Amman, B.R.; Nyakarahuka, L.; McElroy, A.K.; Dodd, K.A.; Sealy, T.K.; Schuh, A.J.; Shoemaker, T.R.; Balinandi, S.; Atimnedi, P.; Kaboyo, W.; et al. Marburgvirus resurgence in Kitaka Mine bat population after extermination attempts, Uganda. Emerg. Infect. Dis. 2014, 20, 1761–1764. [Google Scholar] [CrossRef]
- Mortlock, M.; Dietrich, M.; Weyer, J.; Paweska, J.T.; Markotter, W.J.V. Co-circulation and excretion dynamics of diverse rubula-and related viruses in Egyptian rousette bats from South Africa. Viruses 2019, 11, 37. [Google Scholar] [CrossRef]
- Amman, B.R.; Bird, B.H.; Bakarr, I.A.; Bangura, J.; Schuh, A.J.; Johnny, J.; Sealy, T.K.; Conteh, I.; Koroma, A.H.; Foday, I.; et al. Isolation of Angola-like Marburg virus from Egyptian rousette bats from West Africa. Nat. Commun. 2020, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005. [Google Scholar]
- Amman, B.R.; Schuh, A.J.; Towner, J.S. Ebola Virus Field Sample Collection. In Ebolaviruses: Methods and Protocols; Hoenen, T., Groseth, A., Walker, J.M., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2017; Volume 1628, pp. 373–393. [Google Scholar] [CrossRef]
- Balinandi, S.; Whitmer, S.; Mulei, S.; Nyakarahuka, L.; Tumusiime, A.; Kyondo, J.; Baluku, J.; Mutyaba, J.; Mugisha, L.; Malmberg, M. Clinical and molecular epidemiology of Crimean-Congo hemorrhagic fever in humans in Uganda, 2013–2019. Am. J. Trop. Med. Hyg. 2022, 106, 88. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and high-performance computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Method Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C.-L. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Tavaré, S. Some probabilistic and statistical problems on the analysis of DNA sequence. Lect. Math. Life Sci. Am. Math. Soc. 1986, 17, 57. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Larget, B.; Miller, R.E.; Ronquist, F. Potential applications and pitfalls of Bayesian inference of phylogeny. Syst. Biol. 2002, 51, 673–688. [Google Scholar] [CrossRef]
- Lučan, R.K.; Bartonička, T.; Benda, P.; Bilgin, R.; Jedlička, P.; Nicolaou, H.; Reiter, A.; Shohdi, W.M.; Šálek, M.; Řeřucha, Š. Reproductive seasonality of the Egyptian fruit bat (Rousettus aegyptiacus) at the northern limits of its distribution. J. Mammal. 2014, 95, 1036–1042. [Google Scholar] [CrossRef]
- Penzhorn, B.; Rautenbach, I.L. Reproduction of the Egyptian fruit bat Rousettus aegyptiacus in the southern tropics. S. Afr. J. Wildl. Res. 1988, 18, 88–92. [Google Scholar]
- Epstein, J.H.; Baker, M.L.; Zambrana-Torrelio, C.; Middleton, D.; Barr, J.A.; DuBovi, E.; Boyd, V.; Pope, B.; Todd, S.; Crameri, G. Duration of maternal antibodies against canine distemper virus and Hendra virus in pteropid bats. PLoS ONE 2013, 8, e67584. [Google Scholar] [CrossRef]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G. Ecological dynamics of emerging bat virus spillover. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142124. [Google Scholar] [CrossRef]
- Storm, N.; Jansen Van Vuren, P.; Markotter, W.; Paweska, J.T. Antibody responses to Marburg virus in Egyptian rousette bats and their role in protection against infection. Viruses 2018, 10, 73. [Google Scholar] [CrossRef]
- Klein, S.L. The effects of hormones on sex differences in infection: From genes to behavior. Neurosci. Biobehav. Rev. 2000, 24, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Klein, S. Host factors mediating sex differences in viral infection. Gender Med. 2005, 2, 197–207. [Google Scholar] [CrossRef]
- Kingdon, J. East African Mammals, an Atlas of Evolution in Africa; Academic Press: London, UK, 1974; Volume II. [Google Scholar]
- Amman, B.R.; Schuh, A.J.; Akurut, G.; Kamugisha, K.; Namanya, D.; Sealy, T.K.; Graziano, J.C.; Enyel, E.; Wright, E.A.; Balinandi, S.; et al. Micro-Global Positioning Systems for Identifying Nightly Opportunities for Marburg Virus Spillover to Humans by Egyptian Rousette Bats. Emerg. Infect. Dis. 2023, 29, 2238. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| n | SOSV RNA+ | (%) | n | Anti-SOSV IgG+ | (%) | ||
|---|---|---|---|---|---|---|---|
| Adult | Female | 106 | 27 | 25.47 | 68 | 34 ** | 50.0 |
| Male | 114 | 21 | 18.42 | 85 | 33 | 38.82 | |
| Total | 220 | 48 | 21.82 | 153 | 67 | 43.79 | |
| Juvenile | Female | 81 | 25 | 30.86 | 65 | 27 ** | 41.54 |
| Male | 76 | 25 | 32.90 | 63 | 12 | 19.05 | |
| Total | 157 | 50 | 31.85 | 128 | 39 | 30.47 | |
| Total | 377 | 98 | 26.00 | 281 | 106 * | 37.72 |
| SOSV | Oral Swab | Liver/Spleen | Axillary Lymph Node | Salivary Gland | Colon/Rectum |
|---|---|---|---|---|---|
| Number positive | 2 | 39 | 85 | 28 | 19 |
| CT Min | 27.31 | 32.13 | 27.32 | 31.62 | 29.11 |
| CT Max | 37.13 | 39.88 | 39.68 | 39.57 | 39.02 |
| Human | B1021 | B1271 | B1319 | B1605 | B1624 | B926 | B1302 | B1392 | B1450 | B1516 | B1541 | B1571 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Human | 98.97 | 99.04 | 99.28 | 99.52 | 99.28 | 100.00 | 100.00 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B1021 | 158 | 98.07 | 98.07 | 98.31 | 98.07 | 96.06 | 96.06 | 96.06 | 96.06 | 96.06 | 96.06 | 96.06 | |
| B1271 * | 4 | 8 | 98.8 | 99.04 | 98.8 | 99.21 | 99.21 | 100.00 | 100.00 | 99.21 | 99.21 | 99.21 | |
| B1319 * | 3 | 8 | 5 | 99.28 | 100.00 | 100.00 | 100.00 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B1605 * | 2 | 7 | 4 | 3 | 99.28 | 100.00 | 100.00 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B1624 * | 3 | 8 | 5 | 0 | 3 | 100.00 | 100.00 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B926 ** | 0 | 5 | 1 | 0 | 0 | 0 | 100.00 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B1302 ** | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 99.21 | 99.21 | 100.00 | 100.00 | 100.00 | |
| B1392 ** | 1 | 5 | 0 | 1 | 1 | 1 | 1 | 1 | 100.00 | 99.21 | 99.21 | 99.21 | |
| B1450 ** | 1 | 5 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 99.21 | 99.21 | 99.21 | |
| B1516 ** | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 100.00 | 100.00 | |
| B1541 ** | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 100.00 | |
| B1571 ** | 0 | 5 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amman, B.R.; Koroma, A.H.; Schuh, A.J.; Conteh, I.; Sealy, T.K.; Foday, I.; Johnny, J.; Bakarr, I.A.; Whitmer, S.L.M.; Wright, E.A.; et al. Sosuga Virus Detected in Egyptian Rousette Bats (Rousettus aegyptiacus) in Sierra Leone. Viruses 2024, 16, 648. https://doi.org/10.3390/v16040648
Amman BR, Koroma AH, Schuh AJ, Conteh I, Sealy TK, Foday I, Johnny J, Bakarr IA, Whitmer SLM, Wright EA, et al. Sosuga Virus Detected in Egyptian Rousette Bats (Rousettus aegyptiacus) in Sierra Leone. Viruses. 2024; 16(4):648. https://doi.org/10.3390/v16040648
Chicago/Turabian StyleAmman, Brian R., Alusine H. Koroma, Amy J. Schuh, Immah Conteh, Tara K. Sealy, Ibrahim Foday, Jonathan Johnny, Ibrahim A. Bakarr, Shannon L. M. Whitmer, Emily A. Wright, and et al. 2024. "Sosuga Virus Detected in Egyptian Rousette Bats (Rousettus aegyptiacus) in Sierra Leone" Viruses 16, no. 4: 648. https://doi.org/10.3390/v16040648