


First Molecular Detection and Epidemiological Analysis of Equine Influenza Virus in Two Regions of Colombia, 2020–2023

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Epidemiological Variables: Survey
2.2. Sample Collection
2.3. Molecular Diagnosis and H3N8 Confirmation
2.4. Data Analysis
2.5. Ethical Statement
3. Results
3.1. Molecular Diagnosis and Phylogenetic Analysis
3.2. Zoographic Characteristics
3.3. Bivariate Analysis of Clinical Variables and Treatment
3.4. Characteristics of the Horse Herds
3.5. Shared Space with Other Species
3.6. Vaccination and Sanitary Measures
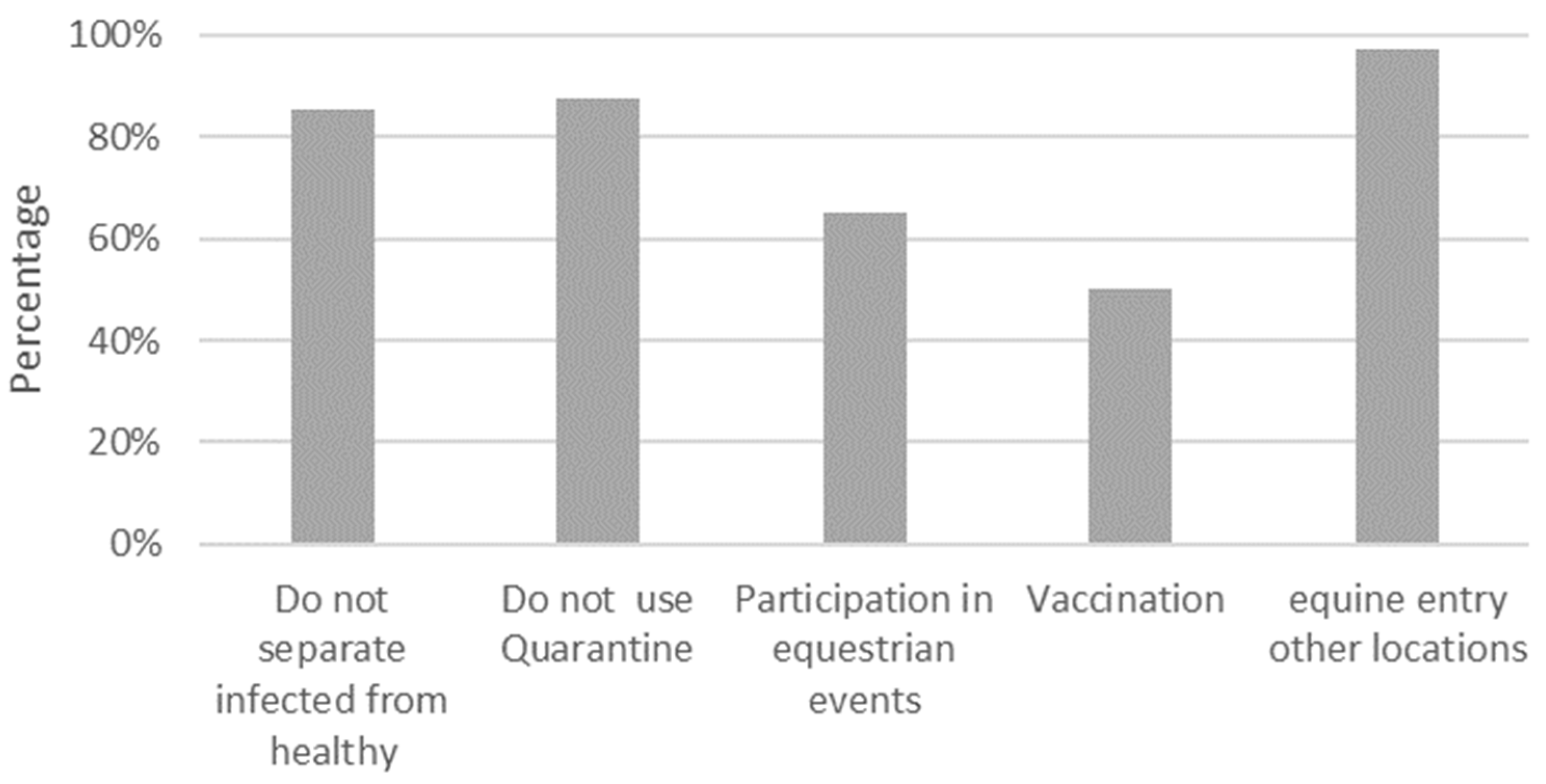
3.7. Other Sanitary Measures Evaluated
3.8. Herd Variables and Disinfection and Health Management
3.9. Veterinary Care and Health Management
3.10. Sanitary Management Analysis at the Herd Level
3.11. Multivariate Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, H.; Damdinjav, B.; Gonzalez, G.; Patrono, L.V.; Ramirez-Mendoza, H.; Amat, J.A.R.; Crispell, J.; Parr, Y.A.; Hammond, T.-a.; Shiilegdamba, E.; et al. Absence of adaptive evolution is the main barrier against influenza emergence in horses in Asia despite frequent virus interspecies transmission from wild birds. PLoS Pathog. 2019, 15, e1007531. [Google Scholar] [CrossRef]
- Singh, R.K.; Dhama, K.; Karthik, K.; Khandia, R.; Munjal, A.; Khurana, S.K.; Chakraborty, S.; Malik, Y.S.; Virmani, N.; Singh, R.; et al. A Comprehensive Review on Equine Influenza Virus: Etiology, Epidemiology, Pathobiology, Advances in Developing Diagnostics, Vaccines, and Control Strategies. Front. Microbiol. 2018, 9, 1941. [Google Scholar] [CrossRef]
- WOAH (Ed.) Chapter 3.6.7. Equine influenza (infection with equine influenza virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; World Organisation for Animal Health: Paris, France, 2019. [Google Scholar]
- Siddell, S.G.; Smith, D.B.; Adriaenssens, E.; Alfenas-Zerbini, P.; Dutilh, B.E.; Garcia, M.L.; Junglen, S.; Krupovic, M.; Kuhn, J.H.; Lambert, A.J.; et al. Virus taxonomy and the role of the International Committee on Taxonomy of Viruses (ICTV). J. Gen. Virol. 2023, 104, 001840. [Google Scholar] [CrossRef]
- Cullinane, A.; Newton, J.R. Equine influenza—A global perspective. Vet. Microbiol. 2013, 167, 205–214. [Google Scholar] [CrossRef]
- Hemida, M.; Perera, R.; Chu, D.; Alnaeem, A.; Peiris, M. Evidence of equine influenza A (H3N8) activity in horses from Eastern and Central Saudi Arabia: 2013–2015. Equine Vet. J. 2019, 51, 218–221. [Google Scholar] [CrossRef]
- Lim, S.-I.; Kim, M.J.; Kim, M.-J.; Lee, S.-K.; Yang, H.-S.; Kwon, M.; Lim, E.H.; Ouh, I.-O.; Kim, E.-J.; Hyun, B.-H. Assessment of Equine Influenza Virus Status in the Republic of Korea from 2020 to 2022. Viruses 2023, 15, 2135. [Google Scholar] [CrossRef]
- Chambers, T.M. Equine influenza. Cold Spring Harb. Perspect. Med. 2022, 12, a038331. [Google Scholar] [CrossRef]
- Ogata, M.; Koizumi, A.; Otsubo, T.; Ikeda, K.; Sakamoto, M.; Aita, R.; Kato, T.; Park, E.Y.; Yamanaka, T.; Hidari, K.I.P.J. Chemoenzymatic synthesis and characterization of N-glycolylneuraminic acid-carrying sialoglycopolypeptides as effective inhibitors against equine influenza virus hemagglutination. Biosci. Biotechnol. Biochem. 2017, 81, 1520–1528. [Google Scholar] [CrossRef]
- Murcia, P.R.; Wood, J.L.N.; Holmes, E.C. Genome-Scale Evolution and Phylodynamics of Equine H3N8 Influenza A Virus. J. Virol. 2011, 85, 5312–5322. [Google Scholar] [CrossRef]
- Mucha, V.; Hollý, J.; Varečková, E.; Kostolanský, F. Avian influenza A virus adaptation to the equine host and identification of host-specific markers. Acta Virol. 2018, 62, 266–276. [Google Scholar] [CrossRef]
- Waghmare, S.; Mode, S.; Kolte, A.; Babhulkar, N.; Vyavahare, S.; Patel, A. Equine influenza: An overview. Vet. World 2010, 3, 194–197. [Google Scholar]
- Collins, P.J.; Vachieri, S.G.; Haire, L.F.; Ogrodowicz, R.W.; Martin, S.R.; Walker, P.A.; Xiong, X.; Gamblin, S.J.; Skehel, J.J. Recent evolution of equine influenza and the origin of canine influenza. Proc. Natl. Acad. Sci. USA 2014, 111, 11175–11180. [Google Scholar] [CrossRef]
- Tu, J.; Zhou, H.; Jiang, T.; Li, C.; Zhang, A.; Guo, X.; Zou, W.; Chen, H.; Jin, M. Isolation and molecular characterization of equine H3N8 influenza viruses from pigs in China. Arch. Virol. 2009, 154, 887–890. [Google Scholar] [CrossRef]
- Crawford, P.C.; Dubovi, E.J.; Castleman, W.L.; Stephenson, I.; Gibbs, E.P.J.; Chen, L.; Smith, C.; Hill, R.C.; Ferro, P.; Pompey, J.; et al. Transmission of Equine Influenza Virus to Dogs. Science 2005, 310, 482–485. [Google Scholar] [CrossRef]
- Su, S.; Wang, L.; Fu, X.; He, S.; Hong, M.; Zhou, P.; Lai, A.; Gray, G.; Li, S. Equine influenza A (H3N8) virus infection in cats. Emerg. Infect. Dis. 2014, 20, 2096–2099. [Google Scholar] [CrossRef]
- Frymus, T.; Belák, S.; Egberink, H.; Hofmann-Lehmann, R.; Marsilio, F.; Addie, D.D.; Boucraut-Baralon, C.; Hartmann, K.; Lloret, A.; Lutz, H.; et al. Influenza Virus Infections in Cats. Viruses 2021, 13, 1435. [Google Scholar] [CrossRef]
- Gonzalez-Obando, J.; Forero, J.E.; Zuluaga-Cabrera, A.M.; Ruiz-Saenz, J. Equine Influenza Virus: An Old Known Enemy in the Americas. Vaccines 2022, 10, 1718. [Google Scholar] [CrossRef]
- Moreira, R.; García, A.; Ahumada, C.; Badía, C.; Suárez, P.; Yangari, B.; Aguayo, C.; Herrera, J.; Espejo, G.; Pinto, E. Report of 2018 equine influenza outbreak in Chile. Austral J. Vet. Sci. 2019, 51, 27–31. [Google Scholar] [CrossRef]
- Oladunni, F.S.; Oseni, S.O.; Martinez-Sobrido, L.; Chambers, T.M. Equine Influenza Virus and Vaccines. Viruses 2021, 13, 1657. [Google Scholar] [CrossRef]
- Olguin Perglione, C.; Golemba, M.D.; Torres, C.; Barrandeguy, M. Molecular Epidemiology and Spatio-Temporal Dynamics of the H3N8 Equine Influenza Virus in South America. Pathogens 2016, 5, 61. [Google Scholar] [CrossRef]
- Olguin-Perglione, C.; Vissani, M.A.; Alamos, F.; Tordoya, M.S.; Barrandeguy, M. Multifocal outbreak of equine influenza in vaccinated horses in Argentina in 2018: Epidemiological aspects and molecular characterisation of the involved virus strains. Equine Vet. J. 2020, 52, 420–427. [Google Scholar] [CrossRef]
- Olguin-Perglione, C.; Barrandeguy, M.E. An Overview of equine influenza in South America. Viruses 2021, 13, 888. [Google Scholar] [CrossRef]
- Colombian Agricultural Institute. RESOLUCIÓN No. 1022 de 1999. Por la Cual se Establecen Medidas Sanitarias Para la Prevención y Control de la Influenza Equina y de la Anemia Infecciosa Equina; Colombian Agricultural Institute: Bogota, Colombia, 1999; p. 2.
- Colombian Agricultural Institute. Livestock Census. Available online: https://www.ica.gov.co/areas/pecuaria/servicios/epidemiologia-veterinaria/censos-2016/censo-2018 (accessed on 20 February 2024).
- Naranjo, E.D. Sector Equino, Asnal y Mular; Colombian Agricultural Institute: Bogota, Colombia, 2018; p. 15.
- Heine, H.; Trinidad, L.; Selleck, P.; Lowther, S. Rapid detection of highly pathogenic avian influenza H5N1 virus by TaqMan reverse transcriptase–polymerase chain reaction. Avian Dis. 2007, 51, 370–372. [Google Scholar] [CrossRef]
- Jurado-Tarifa, E.; Daly, J.M.; Pérez-Écija, A.; Barba-Recreo, M.; Mendoza, F.J.; Al-Shuwaikh, A.M.; García-Bocanegra, I. Epidemiological survey of equine influenza in Andalusia, Spain. Prev. Vet. Med. 2018, 151, 52–56. [Google Scholar] [CrossRef]
- Mena, I.; Nelson, M.I.; Quezada-Monroy, F.; Dutta, J.; Cortes-Fernández, R.; Lara-Puente, J.H.; Castro-Peralta, F.; Cunha, L.F.; Trovão, N.S.; Lozano-Dubernard, B.; et al. Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. eLife 2016, 5, e16777. [Google Scholar] [CrossRef]
- King, J.; Harder, T.; Beer, M.; Pohlmann, A. Rapid multiplex MinION nanopore sequencing workflow for Influenza A viruses. BMC Infect. Dis. 2020, 20, 648. [Google Scholar] [CrossRef]
- Wang, J.; Moore, N.E.; Deng, Y.M.; Eccles, D.A.; Hall, R.J. MinION nanopore sequencing of an influenza genome. Front. Microbiol. 2015, 6, 766. [Google Scholar] [CrossRef]
- Läll, K.; Korhonen, P.; Fischer, K. Generalized Estimating Equations: An Overview and Application in IndiMed Study. Master’s Thesis, University of Tartu, Tartu, Estonia, 2016. [Google Scholar]
- Eberly College of Science. Introduction to Generalized Estimating Equations. Available online: https://online.stat.psu.edu/stat504/lesson/12/12.1 (accessed on 23 January 2024).
- University of Virginia. Getting Started with Generalized Estimating Equations. Available online: https://library.virginia.edu/data/articles/getting-started-with-generalized-estimating-equations#:~:text=Generalized%20estimating%20equations%2C%20or%20GEE,(i.e.%2C%20model%20coefficients) (accessed on 20 February 2024).
- Wang, Y.-W.; Yang, H.-C.; Chen, Y.-H.; Guo, C.-Y. Generalized Estimating Equations Boosting (GEEB) machine for correlated data. J. Big Data 2024, 11, 20. [Google Scholar] [CrossRef]
- Khan, A.; Mushtaq, M.H.; Ahmad, M.U.D.; Nazir, J.; Fatima, Z.; Khan, A.; Farooqi, S.H. Molecular Epidemiology of the Two Internal Genes of Equine Influenza H3N8 Virus Isolated in Pakistan 2015-16. Pak. Vet. J. 2018, 38, 96–100. [Google Scholar]
- Barrington, G.M. Biosecurity. In Llama and Alpaca Care; Cebra, C., Anderson, D.E., Tibary, A., Van Saun, R.J., Johnson, L.W., Eds.; W.B. Saunders: St. Louis, MO, USA, 2014. [Google Scholar] [CrossRef]
- Republica, L. Up to 22 Days per Month Was the Average Rainfall between January and October of Last Year. Available online: https://www.larepublica.co/especiales/lluvias-mil/hasta-22-dias-mensuales-fue-el-promedio-de-lluvias-entre-enero-y-octubre-de-2022-3527844 (accessed on 20 February 2024).
- Pusterla, N.; Kass, P.H.; Mapes, S.; Wademan, C.; Akana, N.; Barnett, C.; MacKenzie, C.; Vaala, W. Voluntary surveillance program for equine influenza virus in the United States from 2010 to 2013. J. Vet. Intern. Med. 2015, 29, 417–422. [Google Scholar] [CrossRef]
- Laval, K.; Poelaert, K.C.K.; Van Cleemput, J.; Zhao, J.; Vandekerckhove, A.P.; Gryspeerdt, A.C.; Garré, B.; van der Meulen, K.; Baghi, H.B.; Dubale, H.N.; et al. The Pathogenesis and Immune Evasive Mechanisms of Equine Herpesvirus Type 1. Front. Microbiol. 2021, 12, 662686. [Google Scholar] [CrossRef] [PubMed]
- Myatt, T.A.; Kaufman, M.H.; Allen, J.G.; MacIntosh, D.L.; Fabian, M.P.; McDevitt, J.J. Modeling the airborne survival of influenza virus in a residential setting: The impacts of home humidification. Environ. Health 2010, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Sack, A.; Daramragchaa, U.; Chuluunbaatar, M.; Gonchigoo, B.; Bazartseren, B.; Tsogbadrakh, N.; Gray, G.C. Low prevalence of enzootic equine influenza virus among horses in Mongolia. Pathogens 2017, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Gildea, S.; Sanchez Higgins, M.J.; Johnson, G.; Walsh, C.; Cullinane, A. Concurrent vaccination against equine influenza and equine herpesvirus–a practical approach. Influenza Other Respir. Viruses 2016, 10, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Boyle, A.G.; Timoney, J.F.; Newton, J.R.; Hines, M.T.; Waller, A.S.; Buchanan, B.R. Streptococcus equi Infections in Horses: Guidelines for Treatment, Control, and Prevention of Strangles—Revised Consensus Statement. J. Vet. Intern. Med. 2018, 32, 633–647. [Google Scholar] [CrossRef] [PubMed]
- BLM. A Virus Has Been Identified as the Principal Cause of the Outbreak and Mortality at Blm’s Wild Horse and Burro Facility; U.S. Bureau of Land Management, Ed.; US Department of Interior: Washington, DC, USA, 2022.
- Chappell, D.E.; Barnett, D.C.; James, K.; Craig, B.; Bain, F.; Gaughan, E.; Schneider, C.; Vaala, W.; Barnum, S.M.; Pusterla, N. Voluntary Surveillance Program for Equine Influenza Virus in the United States during 2008–2021. Pathogens 2023, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Castro, E.; Perez, R.; Rodriguez, S.; Bassetti, L.; Negro, R.; Vidal, R. Epidemiological and virological findings during an outbreak of equine influenza in Uruguay in 2018. Rev. Sci. Tech. (Int. Off. Epizoot.) 2019, 38, 737–749. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Mendez, A.; Viel, L.; Hewson, J.; Doig, P.; Carman, S.; Chambers, T.; Tiwari, A.; Dewey, C. Surveillance of equine respiratory viruses in Ontario. Can. J. Vet. Res. 2010, 74, 271–278. [Google Scholar] [PubMed]
- Ahmed, B.M.; Bayoumi, M.M.; Farrag, M.A.; Elgamal, M.A.; Daly, J.M.; Amer, H.M. Emergence of equine influenza virus H3Nx Florida clade 2 in Arabian racehorses in Egypt. Virol. J. 2022, 19, 185. [Google Scholar] [CrossRef]
- Barbic, L.; Madic, J.; Turk, N.; Daly, J. Vaccine failure caused an outbreak of equine influenza in Croatia. Vet. Microbiol. 2009, 133, 164–171. [Google Scholar] [CrossRef]
- Martella, V.; Elia, G.; Decaro, N.; Di Trani, L.; Lorusso, E.; Campolo, M.; Desario, C.; Parisi, A.; Cavaliere, N.; Buonavoglia, C. An outbreak of equine influenza virus in vaccinated horses in Italy is due to an H3N8 strain closely related to recent North American representatives of the Florida sub-lineage. Vet. Microbiol. 2007, 121, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, M.; Reedy, S.E.; Yano, T.; Suzuki, K.; Fukuda, S.; Garvey, M.; Kambayashi, Y.; Bannai, H.; Tsujimura, K.; Yamanaka, T.; et al. Antigenic comparison of H3N8 equine influenza viruses belonging to Florida sublineage clade 1 between vaccine strains and North American strains isolated in 2021–2022. Arch. Virol. 2023, 168, 94. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, F.; Murcia, P.R.; Newton, J.R. A Review on Equine Influenza from a Human Influenza Perspective. Viruses 2022, 14, 1312. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Mushtaq, M.H.; Ahmad, M.U.D.; Nazir, J.; Farooqi, S.H.; Khan, A. Molecular Epidemiology of a novel re-assorted epidemic strain of equine influenza virus in Pakistan in 2015–16. Virus Res. 2017, 240, 56–63. [Google Scholar] [CrossRef]
- Bravo-Vasquez, N.; Yao, J.; Jimenez-Bluhm, P.; Meliopoulos, V.; Freiden, P.; Sharp, B.; Estrada, L.; Davis, A.; Cherry, S.; Livingston, B. Equine-like H3 avian influenza viruses in wild birds, Chile. Emerg. Infect. Dis. 2020, 26, 2887. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Year | Antioquia | Number of Herds of Antioquia | Positive Herds | Cundinamarca | Number of Herds of Cundinamarca | Positive Herds |
|---|---|---|---|---|---|---|
| 2020 | 0/21 (0%) | 8 | 0 | 0 | 0 | 0 |
| 2021 | 0/38 (0%) | 9 | 0 | 0 | 0 | 0 |
| 2022 | 43/59 (72%) | 14 | 14 | 20/67 (29%) | 8 | 7 |
| 2023 | 0/3 (0%) | 1 | 0 | 0 | 0 | 0 |
| 121 | 32 | 14 | 67 | 8 | 7 |
| Age (Years) | n | Positive | % | p-Value |
|---|---|---|---|---|
| <2 | 67 | 29 | 43.28 | 0.68 |
| 2–5 | 56 | 15 | 26.79 | |
| >5–10 | 38 | 11 | 28.95 | |
| >10 | 27 | 8 | 29.6 | |
| Total | 188 | 63 | ||
| Sex | 0.20 | |||
| Male | 53 | 14 | 26.4 | |
| Female | 135 | 49 | 36.3 | |
| Type of activity | ||||
| Competition | 30 | 16 | 53.3 | 0.12 |
| No competition | 158 | 47 | 29.7 |
| Variables | n | Positive | % | Negative | % | p-Value |
|---|---|---|---|---|---|---|
| Bilateral Nasal Discharge | ||||||
| Yes | 181 | 60 | 95.2 | 121 | 96. 8 | 0.59 |
| No | 7 | 3 | 4.8 | 4 | 3.2 | |
| Dry Cough | ||||||
| Yes | 155 | 59 | 93.7 | 96 | 76.8 | 0.04 * |
| No | 33 | 4 | 6.3 | 29 | 23.2 | |
| Productive cough | ||||||
| Yes | 13 | 5 | 7.9 | 8 | 6.4 | 0.69 |
| No | 175 | 58 | 92.1 | 117 | 93.6 | |
| Fever | ||||||
| Yes | 48 | 26 | 41.3 | 22 | 17.6 | 0.01 * |
| No | 140 | 37 | 58.7 | 103 | 82.4 | |
| Weight loss | ||||||
| Yes | 33 | 14 | 22.2 | 19 | 15.2 | 0.23 |
| No | 155 | 49 | 77.8 | 106 | 84.8 | |
| Respiratory distress | ||||||
| Yes | 16 | 5 | 7.9 | 11 | 8.8 | 0.84 |
| No | 172 | 58 | 92.1 | 114 | 91.2 | |
| Decrease in food consumption | ||||||
| Yes | 56 | 24 | 38.1 | 32 | 25.6 | 0.09 |
| No | 132 | 39 | 61.9 | 93 | 74.4 | |
| Increase in the size of retropharyngeal nodes | ||||||
| Yes | 57 | 33 | 52.4 | 24 | 19.2 | 0.00 * |
| No | 131 | 30 | 47.6 | 101 | 80.8 | |
| Eye discharge | ||||||
| Yes | 10 | 2 | 3.2 | 8 | 6.4 | 0.35 |
| No | 178 | 61 | 96.8 | 117 | 93.6 | |
| Decreased performance | ||||||
| Yes | 51 | 22 | 34.9 | 29 | 23.2 | 0.09 |
| No | 137 | 41 | 65.1 | 96 | 76.8 |
| Disinfection | % |
|---|---|
| Sodium hypochlorite | 27.1% (51/188) |
| Creolin | 22.3% (42/188) |
| Do not disinfect | 13.8% (26/188) |
| Detergent | 12.8% (24/188) |
| Ammonium 10% | 11.7% (22/188) |
| Glutaraldehyde | 9.0% (17/188) |
| Lime | 3.2% (6/188) |
| Do not disinfect | 13.8% (26/188) |
| Detergent | 12.8% (24/188) |
| Veterinary management | % |
| On request if a problem occur | 54.8% (103/188) |
| Permanent | 45.2% (85/188) |
| Variables | CRUDE OR | 95% CI | p-Value | ORadjusted using GEE | 95% CI | p-Value | ||
|---|---|---|---|---|---|---|---|---|
| Do not isolate sick from healthy animals | 13.66 | 1.52 | 122.2 | 0.19 | 8.16 | 1.52 | 43.67 | 0.014 * |
| Share space with poultry | 2.43 | 1.26 | 4.68 | 0.007 | 2.16 | 1.09 | 4.26 | 0.027 * |
| Live on property with equines that participate in equestrian events | 2.57 | 1.25 | 5.28 | 0.01 | 1.32 | 0.612 | 2.86 | 0.47 |
| Competition equine | 2.7 | 1.22 | 5.99 | 0.12 | 1.92 | 0.75 | 4.9 | 0.17 |
| Do not quarantine | 1.90 | 1.01 | 3.54 | 0.044 | 1.88 | 0.41 | 8.52 | 0.16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Obando, J.; Zuluaga-Cabrera, A.; Moreno, I.; Úsuga, J.; Ciuderis, K.; Forero, J.E.; Diaz, A.; Rojas-Arbeláez, C.; Hernández-Ortiz, J.P.; Ruiz-Saenz, J. First Molecular Detection and Epidemiological Analysis of Equine Influenza Virus in Two Regions of Colombia, 2020–2023. Viruses 2024, 16, 839. https://doi.org/10.3390/v16060839
Gonzalez-Obando J, Zuluaga-Cabrera A, Moreno I, Úsuga J, Ciuderis K, Forero JE, Diaz A, Rojas-Arbeláez C, Hernández-Ortiz JP, Ruiz-Saenz J. First Molecular Detection and Epidemiological Analysis of Equine Influenza Virus in Two Regions of Colombia, 2020–2023. Viruses. 2024; 16(6):839. https://doi.org/10.3390/v16060839
Chicago/Turabian StyleGonzalez-Obando, Juliana, Angélica Zuluaga-Cabrera, Isabel Moreno, Jaime Úsuga, Karl Ciuderis, Jorge E. Forero, Andrés Diaz, Carlos Rojas-Arbeláez, Juan P. Hernández-Ortiz, and Julian Ruiz-Saenz. 2024. "First Molecular Detection and Epidemiological Analysis of Equine Influenza Virus in Two Regions of Colombia, 2020–2023" Viruses 16, no. 6: 839. https://doi.org/10.3390/v16060839
APA StyleGonzalez-Obando, J., Zuluaga-Cabrera, A., Moreno, I., Úsuga, J., Ciuderis, K., Forero, J. E., Diaz, A., Rojas-Arbeláez, C., Hernández-Ortiz, J. P., & Ruiz-Saenz, J. (2024). First Molecular Detection and Epidemiological Analysis of Equine Influenza Virus in Two Regions of Colombia, 2020–2023. Viruses, 16(6), 839. https://doi.org/10.3390/v16060839