
Bovine Rhinitis B Virus Variant as the Putative Cause of Bronchitis in Goat Kids

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Case History and Sample Collection
2.2. Routine Diagnostics
2.3. Whole-Genome Sequencing, Metagenomics, and Phylogenetic Analysis
2.4. Virus Isolation, PCR, and In Situ Hybridization
3. Results
3.1. Routine Diagnostics
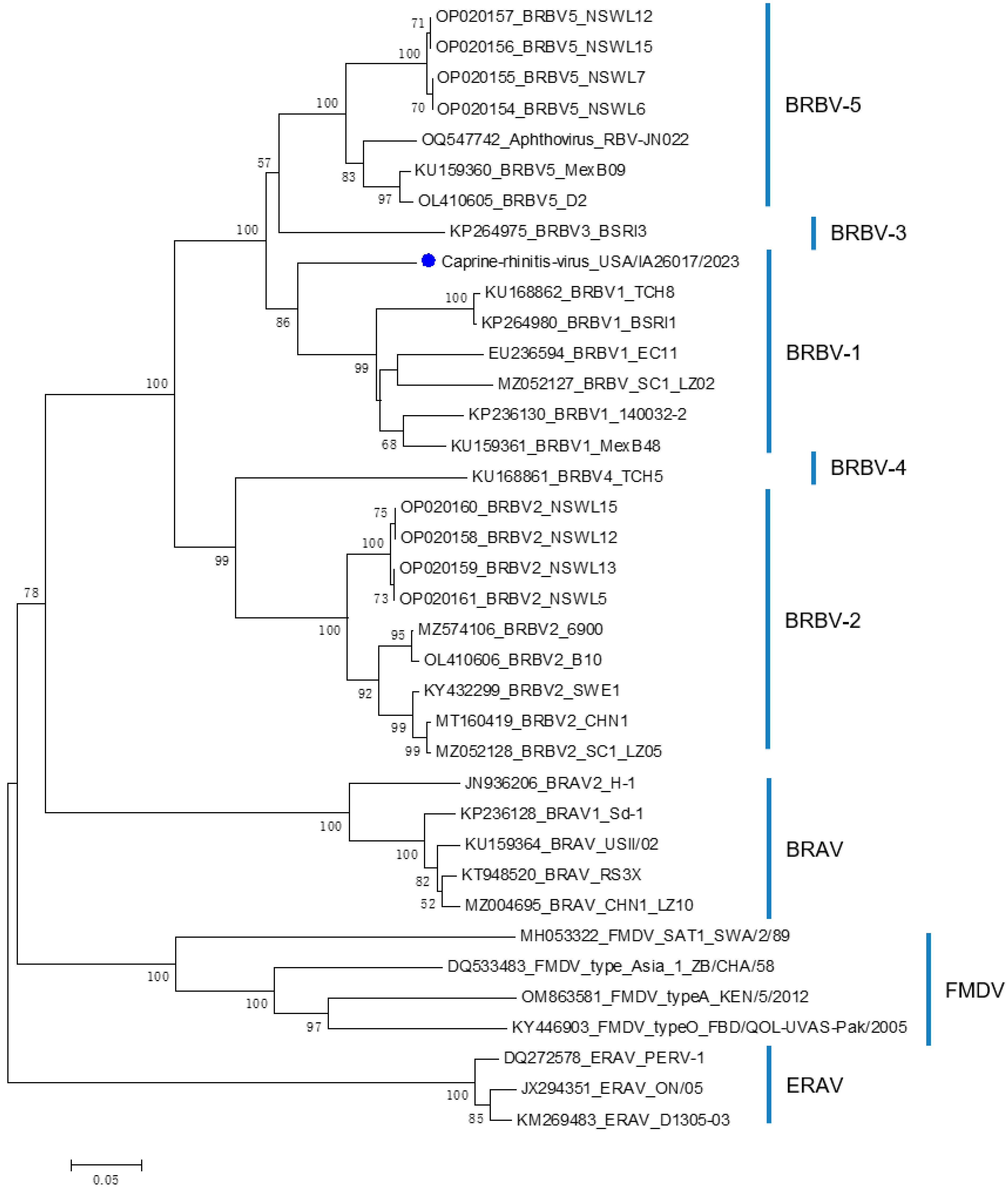
3.2. Whole-Genome Sequencing, Metagenomics, and Phylogenetic Analysis
3.3. Virus Isolation, PCR, and In Situ Hybridization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tuthill, T.J.; Groppelli, E.; Hogle, J.M.; Rowlands, D.J. Picornaviruses. Curr. Top. Microbiol. Immunol. 2010, 343, 43–89. [Google Scholar] [PubMed]
- Kriegshäuser, G.; Deutz, A.; Kuechler, E.; Skern, T.; Lussy, H.; Nowotny, N. Prevalence of neutralizing antibodies to Equine rhinitis A and B virus in horses and man. Vet. Microbiol. 2005, 106, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Plummer, G. An equine respiratory enterovirus. Some biological and physical properties. Arch. Gesamte Virusforsch. 1963, 12, 694–700. [Google Scholar] [CrossRef] [PubMed]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus taxonomy: The database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, D708–D717. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, S.; Lin, C.M.; Temeeyasen, G.; Palinski, R.; Li, F.; Kaushik, R.S.; Hause, B.M. Bovine rhinitis B virus is highly prevalent in acute bovine respiratory disease and causes upper respiratory tract infection in calves. J. Gen. Virol. 2022, 103, 001714. [Google Scholar] [CrossRef] [PubMed]
- Hollister, J.R.; Vagnozzi, A.; Knowles, N.J.; Rieder, E. Molecular and phylogenetic analyses of bovine rhinovirus type 2 shows it is closely related to foot-and-mouth disease virus. Virology 2008, 373, 411–425. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Mao, L.; Cheng, S.; Wang, Q.; Huang, J.; Deng, J.; Wang, Z.; Zhang, W.; Yang, L.; Hao, F.; et al. A novel parainfluenza virus type 3 (PIV3) identified from goat herds with respiratory diseases in eastern China. Vet. Microbiol. 2014, 174, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Horwood, P.F.; Mahony, T.J. Multiplex real-time RT-PCR detection of three viruses associated with the bovine respiratory disease complex. J. Virol. Methods 2011, 171, 360–363. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, L.; Zheng, Y.; Zhang, J.; Guo, B.; Yoon, K.J.; Gauger, P.C.; Harmon, K.M.; Main, R.G.; Li, G. Metagenomic analysis of the RNA fraction of the fecal virome indicates high diversity in pigs infected by porcine endemic diarrhea virus in the United States. Virol. J. 2018, 15, 95. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Kishimoto, M.; Tsuchiaka, S.; Rahpaya, S.S.; Hasebe, A.; Otsu, K.; Sugimura, S.; Kobayashi, S.; Komatsu, N.; Nagai, M.; Omatsu, T.; et al. Development of a one-run real-time PCR detection system for pathogens associated with bovine respiratory disease complex. J. Vet. Med. Sci. 2017, 79, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, X.; Tang, C.; Yue, H. Detection and Genomic Characterization of Bovine Rhinitis Virus in China. Animals 2023, 13, 312. [Google Scholar] [CrossRef] [PubMed]
- Rahe, M.C.; Magstadt, D.R.; Groeltz-Thrush, J.; Gauger, P.C.; Zhang, J.; Schwartz, K.J.; Siepker, C.L. Bovine coronavirus in the lower respiratory tract of cattle with respiratory disease. J. Vet. Diagn. Investig. 2022, 34, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.R.; Brogden, K.A. Response of the ruminant respiratory tract to Mannheimia (Pasteurella) haemolytica. Microbes Infect. 2000, 2, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Hill, J.E.; Fernando, C.; Alexander, T.W.; Timsit, E.; van der Meer, F.; Huang, Y. Respiratory viruses identified in western Canadian beef cattle by metagenomic sequencing and their association with bovine respiratory disease. Transbound. Emerg. Dis. 2019, 66, 1379–1386. [Google Scholar] [CrossRef] [PubMed]
- Mosier, D. Review of BRD pathogenesis: The old and the new. Anim. Health Res. Rev. 2014, 15, 166–168. [Google Scholar] [CrossRef]
- Gioia, J.; Qin, X.; Jiang, H.; Clinkenbeard, K.; Lo, R.; Liu, Y.; Fox, G.E.; Yerrapragada, S.; McLeod, M.P.; McNeill, T.Z.; et al. The genome sequence of Mannheimia haemolytica A1: Insights into virulence, natural competence, and Pasteurellaceae phylogeny. J. Bacteriol. 2006, 188, 7257–7266. [Google Scholar] [CrossRef]
- Maksimović, Z.; Rifatbegović, M.; Loria, G.R.; Nicholas, R.A.J. Mycoplasma ovipneumoniae: A Most Variable Pathogen. Pathogens 2022, 11, 1477. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.; Jones, K.; Jacobson, B.T.; Schearer, J.; Adams, N.; Thornton, I.; Mosdal, C.; Jones, S.; Jutila, M.; Rynda-Apple, A.; et al. Experimental infection of specific-pathogen-free domestic lambs with Mycoplasma ovipneumoniae causes asymptomatic colonization of the upper airways that is resistant to antibiotic treatment. Vet. Microbiol. 2022, 265, 109334. [Google Scholar] [CrossRef]
- DaMassa, A.J.; Wakenell, P.S.; Brooks, D.L. Mycoplasmas of goats and sheep. J. Vet. Diagn. Investig. 1992, 4, 101–113. [Google Scholar] [CrossRef]
- Yang, F.; Tang, C.; Wang, Y.; Zhang, H.; Yue, H. Genome sequence of Mycoplasma ovipneumoniae strain SC01. J. Bacteriol. 2011, 193, 5018. [Google Scholar] [CrossRef]
- Huaman, J.L.; Pacioni, C.; Sarker, S.; Doyle, M.; Forsyth, D.M.; Pople, A.; Carvalho, T.G.; Helbig, K.J. Novel Picornavirus Detected in Wild Deer: Identification, Genomic Characterisation, and Prevalence in Australia. Viruses 2021, 13, 2412. [Google Scholar] [CrossRef]
- Neff, S.; Sá-Carvalho, D.; Rieder, E.; Mason, P.W.; Blystone, S.D.; Brown, E.J.; Baxt, B. Foot-and-mouth disease virus virulent for cattle utilizes the integrin alpha(v)beta3 as its receptor. J. Virol. 1998, 72, 3587–3594. [Google Scholar] [CrossRef]
- Fry, E.E.; Tuthill, T.J.; Harlos, K.; Walter, T.S.; Rowlands, D.J.; Stuart, D.I. Crystal structure of Equine rhinitis A virus in complex with its sialic acid receptor. J. Gen. Virol. 2010, 91, 1971–1977. [Google Scholar] [CrossRef]
- Jackson, T.; Sheppard, D.; Denyer, M.; Blakemore, W.; King, A.M. The epithelial integrin alphavbeta6 is a receptor for foot-and-mouth disease virus. J. Virol. 2000, 74, 4949–4956. [Google Scholar] [CrossRef] [PubMed]
- Duque, H.; LaRocco, M.; Golde, W.T.; Baxt, B. Interactions of foot-and-mouth disease virus with soluble bovine alphaVbeta3 and alphaVbeta6 integrins. J. Virol. 2004, 78, 9773–9781. [Google Scholar] [CrossRef]
- Monaghan, P.; Gold, S.; Simpson, J.; Zhang, Z.; Weinreb, P.H.; Violette, S.M.; Alexandersen, S.; Jackson, T. The alpha(v)beta6 integrin receptor for Foot-and-mouth disease virus is expressed constitutively on the epithelial cells targeted in cattle. J. Gen. Virol. 2005, 86, 2769–2780. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noel, A.; Zhang, J.; Shen, H.; Saxena, A.; Groeltz-Thrush, J.; Li, G.; Rahe, M.C. Bovine Rhinitis B Virus Variant as the Putative Cause of Bronchitis in Goat Kids. Viruses 2024, 16, 1023. https://doi.org/10.3390/v16071023
Noel A, Zhang J, Shen H, Saxena A, Groeltz-Thrush J, Li G, Rahe MC. Bovine Rhinitis B Virus Variant as the Putative Cause of Bronchitis in Goat Kids. Viruses. 2024; 16(7):1023. https://doi.org/10.3390/v16071023
Chicago/Turabian StyleNoel, Andrew, Jianqiang Zhang, Huigang Shen, Anugrah Saxena, Jennifer Groeltz-Thrush, Ganwu Li, and Michael C. Rahe. 2024. "Bovine Rhinitis B Virus Variant as the Putative Cause of Bronchitis in Goat Kids" Viruses 16, no. 7: 1023. https://doi.org/10.3390/v16071023
APA StyleNoel, A., Zhang, J., Shen, H., Saxena, A., Groeltz-Thrush, J., Li, G., & Rahe, M. C. (2024). Bovine Rhinitis B Virus Variant as the Putative Cause of Bronchitis in Goat Kids. Viruses, 16(7), 1023. https://doi.org/10.3390/v16071023