
Semi-Covariance Coefficient Analysis of Spike Proteins from SARS-CoV-2 and Its Variants Omicron, BA.5, EG.5, and JN.1 for Viral Infectivity, Virulence and Immune Escape

Abstract
:1. Introduction
2. Literature Review
2.1. Research on Spike Proteins of SARS-CoV-2
2.2. Research on Electrostatic Feature of SARS-CoV-2 Spike Protein
3. Methodologies
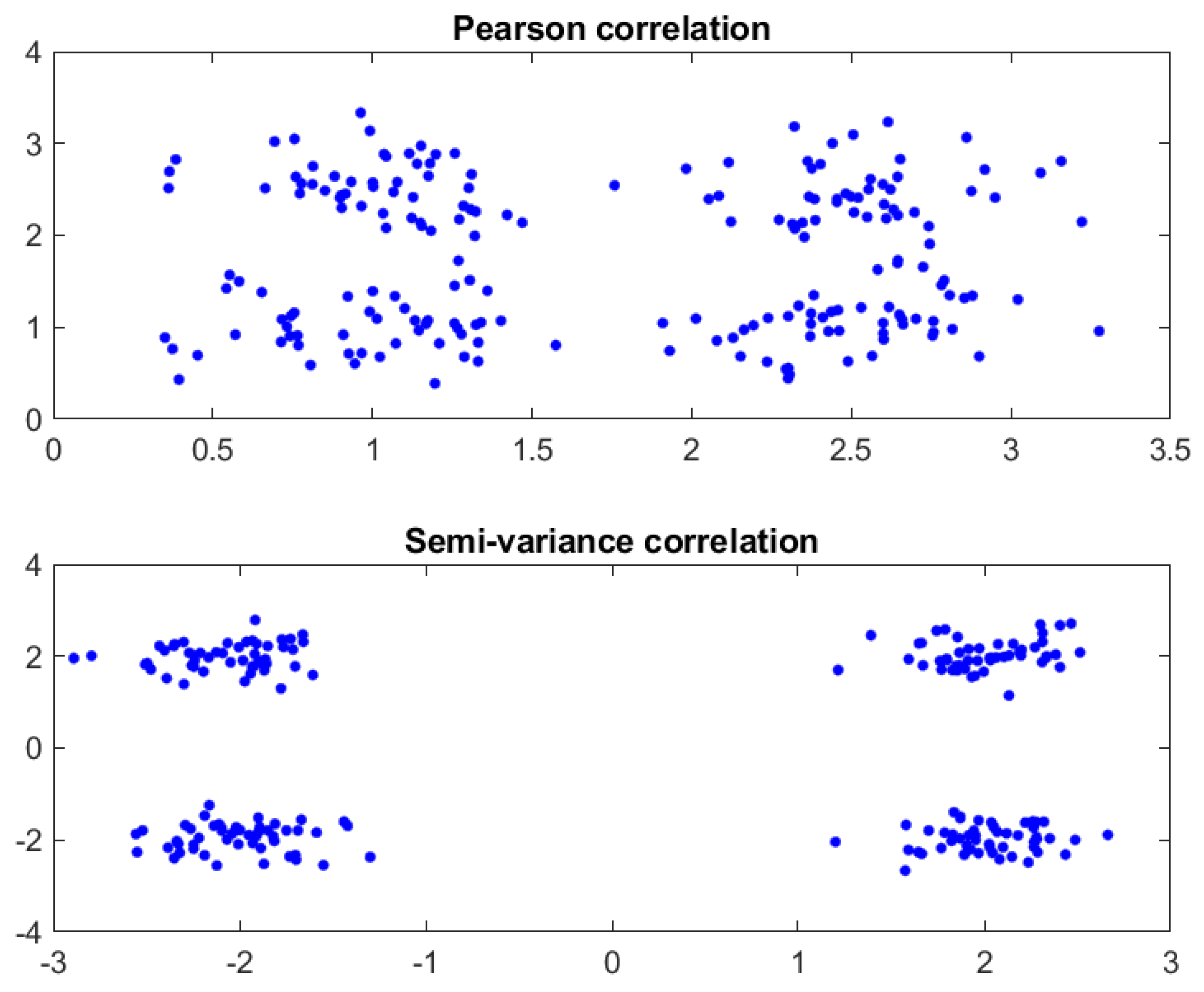
3.1. Pearson Correlation Coefficient
3.2. Semi-Variance Correlation
3.2.1. Semi-Variance Correlation Coefficient
3.2.2. Quantity of Charge
3.2.3. Gravity
3.2.4. Charge Span
3.2.5. Reproduction Rate
3.2.6. Maximal Position
3.2.7. Charge Density
3.2.8. Virulence
4. Result and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Peters, E.E. Fractal DNA Analysis: Applying Chaos Theory to Investment and Economics; Wiley: Hoboken, NJ, USA, 1994. [Google Scholar]
- Peng, C.K.; Buldyrev, S.V.; Goldberger, A.L.; Havlin, S.; Sciortino, F.; Simons, M.; Stanley, H.E. Long-range correlations in nucleotide sequences. Nature 1992, 356, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Voss, R.F. Evolution of long-range fractal correlations and 1/f noise in DNA base sequences. Phys. Rev. Lett. 1992, 68, 3805–3808. [Google Scholar] [CrossRef] [PubMed]
- Buldyrev, S.V.; Goldberger, A.L.; Havlin, S.; Mantegna, R.N.; Matsa, M.E.; Peng, C.K.; Simons, M.; Stanley, H.E. Long-range correlation properties of coding and noncoding DNA sequences: GenBank analysis. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 1995, 51, 5084–5091. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Bai, Y. Semiparametric statistical inferences for longitudinal data with nonparametric covariance modelling. Statistics 2017, 51, 1280–1303. [Google Scholar] [CrossRef]
- Fernandes, T.; Oliveira Filho, J.S.; Silva Lopes, I.M.S. Fractal signature of coronaviruses related to severe acute respiratory syndrome. Res. Biomed. Eng. 2022, 38, 293–297. [Google Scholar] [CrossRef]
- Kumar, S.; Karuppanan, K.; Subramaniam, G. Omicron (BA.1) and sub-variants (BA.1.1, BA.2, and BA.3) of SARS-CoV-2 spike infectivity and pathogenicity: A comparative sequence and structural-based computational assessment. J. Med. Virol. 2022, 94, 4780–4791. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.S.; Huang, J.M.; Zhang, W. Semi-covariance coefficient analysis of spike proteins from SARS-CoV-2 and other coronaviruses for viral evolution and characteristics associated with fatality. Entropy 2021, 23, 512. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.K.; Buldyrev, S.V.; Havlin, S.; Simons, M.; Stanley, H.E.; Goldberger, A.L. Mosaic organization of DNA nucleotides. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 1994, 49, 1685–1689. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhou, S.; Huang, J.S.; Zhang, W. Normalized semi-covariance co-efficiency analysis of spike proteins from SARS-CoV-2 variant Omicron and other coronaviruses for their infectivity and virulence. Arch. Clin. Biomed. Res. 2023, 7, 387–399. [Google Scholar] [CrossRef]
- Hathout, R.M.; Kassem, D.H. Positively charged electroceutical spun chitosan nanofibers can protect health care providers from COVID-19 infection: An opinion. Front. Bioeng. Biotechnol. 2020, 8, 885. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.; Tiu, C.; Hu, Z.; Chen, V.C.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2-spike protein-protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, J.; Nguyen, T.; Adkins, J.L.; Schank, M.; Khanal, S.; Nguyen, L.; Dang, X.; Cao, D.; Thakuri, C.; et al. Blockade of SARS-CoV-2 spike protein-mediated cell-cell fusion using COVID-19 convalescent plasma. Sci. Rep. 2021, 11, 5558. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, J.H.; Lee, K.; Gim, Y. The prediction of COVID-19 using LSTM algorithms. Int. J. Netw. Distrib. Comput. 2021, 9, 19–24. [Google Scholar] [CrossRef]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 579, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Desingu, P.A.; Nagarajan, K.; Dhama, K. SARS-CoV-2 gained a novel spike protein S1-N-Terminal Domain (S1-NTD). Environ. Res. 2022, 211, 113047. [Google Scholar] [CrossRef]
- Mittal, A.; Khattri, A.; Verma, V. Structural and antigenic variations in the spike protein of emerging SARS-CoV-2 variants. PLoS Pathog. 2022, 18, e1010260. [Google Scholar] [CrossRef] [PubMed]
- Tao, K.; Tzou, P.L.; Nouhin, J.; Gupta, R.K.; Oliveira, T.; Kosakovsky Pond, S.L.; Fera, D.; Shafer, R.W. The biological and clinical significance of emerging SARS-CoV-2 variants. Nat. Rev. Genet. 2021, 22, 757–773. [Google Scholar] [CrossRef]
- Banerjee, S.; Wang, X.; Du, S.; Zhu, C.; Jia, Y.; Wang, Y.; Cai, Q. Comprehensive role of SARS-CoV-2 spike glycoprotein in regulating host signaling pathway. J. Med. Virol. 2022, 94, 4071–4087. [Google Scholar] [CrossRef] [PubMed]
- Kumavath, R.; Barh, D.; Andrade, B.S.; Imchen, M.; Aburjaile, F.F.; Ch, A.; Rodrigues, D.L.N.; Tiwari, S.; Alzahrani, K.J.; Góes-Neto, A.; et al. The spike of SARS-CoV-2: Uniqueness and applications. Front. Immunol. 2021, 12, 663912. [Google Scholar] [CrossRef] [PubMed]
- Cocherie, T.; Zafilaza, K.; Leducq, V.; Marot, S.; Calvez, V.; Marcelin, A.G.; Todesco, E. Epidemiology and characteristics of SARS-CoV-2 variants of concern: The impacts of the spike mutations. Microorganisms 2023, 11, 30. [Google Scholar] [CrossRef]
- Kuzmina, A.; Wattad, S.; Engel, S.; Rosenberg, E.; Taube, R. Functional analysis of spike from SARS-CoV-2 variants reveals the role of distinct mutations in neutralization potential and viral infectivity. Viruses 2022, 14, 803. [Google Scholar] [CrossRef] [PubMed]
- Cordsmeier, A.; Jungnickl, D.; Herrmann, A.; Korn, K.; Ensser, A. Analysis of SARS-CoV-2 spike protein variants with recombinant reporter viruses created from a bacmid system. Int. J. Mol. Sci. 2023, 24, 8156. [Google Scholar] [CrossRef] [PubMed]
- Emmelot, M.E.; Vos, M.; Boer, M.C.; Rots, N.Y.; van Els, C.A.C.M.; Kaaijk, P. SARS-CoV-2 Omicron BA.4/BA.5 mutations in spike leading to T cell escape in recently vaccinated individuals. Viruses 2023, 15, 101. [Google Scholar] [CrossRef] [PubMed]
- Bains, A.; Guan, W.; LiWang, P.J. The effect of select SARS-CoV-2 n-linked glycan and variant of concern spike protein mutations on c-type lectin-receptor-mediated infection. Viruses 2023, 15, 1901. [Google Scholar] [CrossRef] [PubMed]
- Escalera, A.; Gonzalez-Reiche, A.S.; Aslam, S.; Mena, I.; Laporte, M.; Pearl, R.L.; Fossati, A.; Rathnasinghe, R.; Alshammary, H.; van de Guchte, A.; et al. Mutations in SARS-CoV-2 variants of concern link to increased spike cleavage and virus transmission. Cell Host Microbe 2022, 30, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Pastorio, C.; Zech, F.; Noettger, S.; Jung, C.; Jacob, T.; Sanderson, T.; Sparrer, K.M.J.; Kirchhoff, F. Determinants of spike infectivity, processing, and neutralization in SARS-CoV-2 Omicron subvariants BA.1 and BA.2. Cell Host Microbe 2022, 30, 1255–1268. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, J.; Wang, J. Surface charge changes in spike RBD mutations of SARS-CoV-2 and its variant strains alter the virus evasiveness via HSPGs: A review and mechanistic hypothesis. Front. Public Health 2022, 10, 952916. [Google Scholar] [CrossRef]
- Pawlowski, P.H.; Zielenkiewicz, P. The role of electric charge in SARS-CoV-2 and other viral infections. AIMS Biophys. 2024, 11, 166–189. [Google Scholar] [CrossRef]
- Magazine, N.; Zhang, T.; Wu, Y.; McGee, M.C.; Veggiani, G.; Huang, W. Mutations and evolution of the SARS-CoV-2 spike protein. Viruses 2022, 14, 640. [Google Scholar] [CrossRef]
- Pawłowski, P.H. Additional Positive Electric Residues in the Crucial Spike Glycoprotein S Regions of the New SARS-CoV-2 Variants. Infect. Drug Resist. 2021, 14, 5099–5105. [Google Scholar] [CrossRef]
- Cotton, M.; Phan, M.V.T. Evolution of increased positive charge on the SARS-CoV-2 spike protein may be adaptation to human transmission. iScience 2023, 26, 106230. [Google Scholar] [CrossRef] [PubMed]
- Jawad, B.; Adhikari, P.; Podgornik, R.; Ching, W.Y. Binding Interactions between Receptor-Binding Domain of Spike Protein and Human Angiotensin Converting Enzyme-2 in Omicron Variant. J. Phys. Chem. Lett. 2022, 13, 3915–3921. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, P.; Jawad, B.; Podgornik, R.; Ching, W.Y. Mutations of Omicron Variant at the Interface of the Receptor Domain Motif and Human Angiotensin-Converting Enzyme-2. Int. J. Mol. Sci. 2022, 23, 2870. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.L.; Lan, P.D.; Thai, N.Q.; Nissley, D.A.; O’Brien, E.P.; Li, M.S. Does SARS-CoV-2 bind to human ACE2 more strongly than does SARS-CoV. J. Phys. Chem. 2020, 124, 7336–7347. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Kearns, F.L.; Rosenfeld, M.A.; Votapka, L.; Casalino, L.; Papanikolas, M.; Rommie, E.; Amaro, R.E.; Freeman, R. Positively bound: Remapping of increased positive charge drives SARS-CoV-2 spike evolution to optimize its binding to cell surface receptors. ChemRxiv 2022. [Google Scholar] [CrossRef]
- Božič, A.; Podgornik, R. Changes in total charge on spike protein of SARS-CoV-2 in emerging lineages. Bioinform. Adv. 2024, 4, vbae053. [Google Scholar] [CrossRef] [PubMed]
- Pascarella, S.; Ciccozzi, M.; Zella, D.; Bianchi, M.; Benedetti, F.; Benvenuto, D.; Broccolo, F.; Cauda, R.; Caruso, A.; Angeletti, S.; et al. SARS-CoV-2 B.1.617 Indian variants: Are electrostatic potential changes responsible for a higher transmission rate? J. Med. Virol. 2021, 93, 6551–6556. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhao, T.; Lu, M.; Zhang, Y.; Yao, X.; Wu, G.; Dai, F.; Zhang, F.; Zhang, G. The analyses of high infectivity mechanism of SARS-CoV-2 and its variants. COVID 2021, 1, 666–673. [Google Scholar] [CrossRef]
- Barroso da Silva, F.L.; Giron, C.C.; Laaksonen, A. Electrostatic features for the receptor binding domain of SARS-COV-2 wildtype and its variants. Compass to the severity of the future variants with the charge-rule. J. Phys. Chem. B 2022, 126, 6835–6852. [Google Scholar] [CrossRef] [PubMed]
- Saccenti, E.; Hendriks, B.; Smilde, A.K. Corruption of the pearson correlation coefficient by measurement error and its estimation, bias, and correction under different error models. Sci. Rep. 2020, 10, 438. [Google Scholar] [CrossRef]
- Chen, Y.; Dai, X.; Liu, M.; Chen, D.; Yuan, L.; Liu, Z. Dynamic relu. In Proceedings of the European Conference on Computer Vision, Glasgow, UK, 23–28 August 2020; pp. 351–367. [Google Scholar]
- Carabelli, A.M.; Peacock, T.P.; Thorne, L.G.; Harvey, W.T.; Hughes, J. SARS-CoV-2 variant biology: Immune escape, transmission and fitness. Nat. Rev. Microbiol. 2023, 21, 162–177. [Google Scholar] [CrossRef] [PubMed]
- Castonguay, N.; Zhang, W.; Langlois, M.A. Meta-analysis and structural dynamics of the emergence of genetic variants of SARS-CoV-2. Front. Microbiol. 2021, 12, 676314. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| UK (B.1.1.7) | Delta (B.1.617) | Omicron | BA.5 | EG.5 | JN.1 | |
|---|---|---|---|---|---|---|
| 99.58% | 99.44% | 98.09% | 96.04% | 87.63% | 63.9% | |
| 0.32% | 0.17% | 1% | 0.4% | 1.95% | 7.75% | |
| 0.9926 | 0.9927 | 0.9709 | 0.9564 | 0.8559 | 0.5615 | |
| 17 | 15 | 18 | 18 | 17 | 19 | |
| 626 | 629 | 631 | 665 | 718 | 868 | |
| 655 | 557 | 361 | 568 | 167 | 303 | |
| * | 29 | 72 | 270 | 97 | 551 | 565 |
| 0.84 | 2.08 | 7.79 | 2.8 | 15.89 | 16.3 | |
| 516 | 518 | 518 | 1264 | 1262 | 1262 | |
| 614 | 616 | 216 | 683 | 97 | 97 | |
| * | 1 | 0.4 | 0.31 | 0.66 | 0.82 | 0.8 |
| 5.33% | 2.15% | 1.64% | 3.51% | 4.35% | 4.24% | |
| Actual rate of death (sourced from the Internet) | 1.3–5.3% | 0.3–3.4% | 0.06–0.3% | 0.06–0.3% | / | 1.81% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, B.; Lin, H.; Huang, J.S.; Zhang, W. Semi-Covariance Coefficient Analysis of Spike Proteins from SARS-CoV-2 and Its Variants Omicron, BA.5, EG.5, and JN.1 for Viral Infectivity, Virulence and Immune Escape. Viruses 2024, 16, 1192. https://doi.org/10.3390/v16081192
Zhu B, Lin H, Huang JS, Zhang W. Semi-Covariance Coefficient Analysis of Spike Proteins from SARS-CoV-2 and Its Variants Omicron, BA.5, EG.5, and JN.1 for Viral Infectivity, Virulence and Immune Escape. Viruses. 2024; 16(8):1192. https://doi.org/10.3390/v16081192
Chicago/Turabian StyleZhu, Botao, Huancheng Lin, Jun Steed Huang, and Wandong Zhang. 2024. "Semi-Covariance Coefficient Analysis of Spike Proteins from SARS-CoV-2 and Its Variants Omicron, BA.5, EG.5, and JN.1 for Viral Infectivity, Virulence and Immune Escape" Viruses 16, no. 8: 1192. https://doi.org/10.3390/v16081192