
High-Throughput Screening of Antiviral Compounds Using a Recombinant Hepatitis B Virus and Identification of a Possible Infection Inhibitor, Skimmianine

 , , , , , and add
Show full author list
, , , , , and add
Show full author list
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plasmids
2.2. Cells
2.3. Virus Preparations and Infection
2.4. Reagents
2.5. HiBiT Assay
2.6. Quantitation of Extracellular HBV-DNA
2.7. Quantitation of HBsAg
2.8. Determination of the EC50
2.9. Cell Viability Assay
2.10. Live Cell Confocal Imaging
2.11. Attachment and Internalization Assay
2.12. Statistical Analysis
3. Results
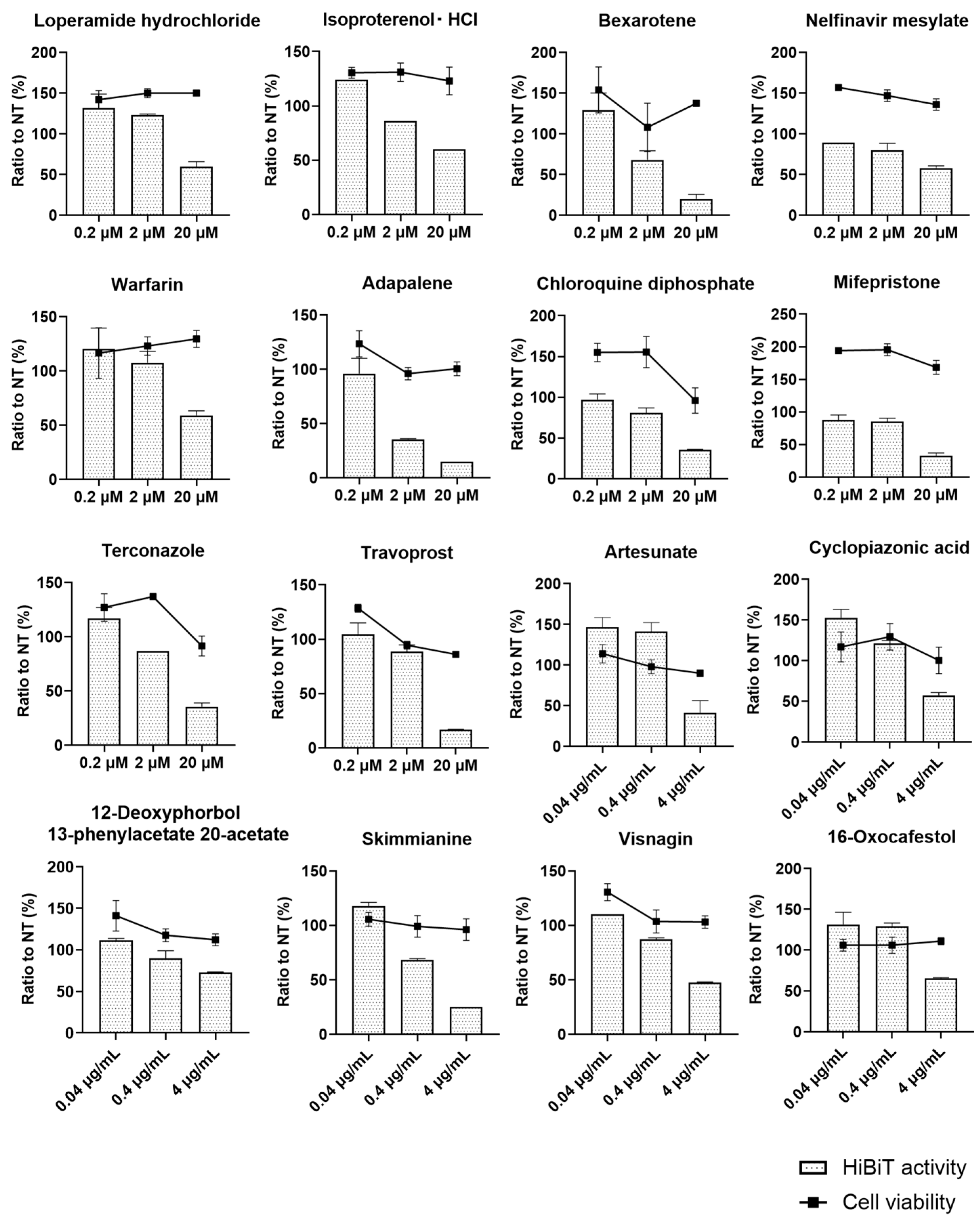
3.1. Antiviral Screening of the Compound Library with Recombinant HBV
3.2. Antiviral Effect of Skimmianine and Its Cytotoxicity
3.3. Antiviral Effect of Skimmianine at Lower Concentrations
3.4. Inhibitory Target of Skimmianine in HBV Infection
3.5. Effect of Skimmianine on HBV Replication in HepG2.2.15 Cells
3.6. Inhibition of Viral Entry by Skimmianine in Time-Lapse Fluorescence Imaging Analysis
3.7. Inhibitory Effect of Skimmianine on Viral Entry for Non-Recombinant HBV
3.8. Effect of Skimmianine on Attachment and Internalization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- The Polaris Observatory Collaborators. Global prevalence, treatment, and prevention of hepatitis B virus infection in 2016: A modelling study. Lancet Gastroenterol. Hepatol. 2018, 3, 383–403. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Cui, X.; Xie, Y.; Liu, J. Engineering Hepadnaviruses as Reporter-Expressing Vectors: Recent Progress and Future Perspectives. Viruses 2016, 8, 125. [Google Scholar] [CrossRef] [PubMed]
- Dixon, A.S.; Schwinn, M.K.; Hall, M.P.; Zimmerman, K.; Otto, P.; Lubben, T.H.; Butler, B.L.; Binkowski, B.F.; Machleidt, T.; Kirkland, T.A.; et al. NanoLuc Complementation Reporter Optimized for Accurate Measurement of Protein Interactions in Cells. ACS Chem. Biol. 2016, 11, 400–408. [Google Scholar] [CrossRef]
- Schwinn, M.K.; Machleidt, T.; Zimmerman, K.; Eggers, C.T.; Dixon, A.S.; Hurst, R.; Hall, M.P.; Encell, L.P.; Binkowski, B.F.; Wood, K.V. CRISPR-Mediated Tagging of Endogenous Proteins with a Luminescent Peptide. ACS Chem. Biol. 2018, 13, 467–474. [Google Scholar] [CrossRef]
- Sumiyadorj, A.; Murai, K.; Shimakami, T.; Kuroki, K.; Nishikawa, T.; Kakuya, M.; Yamada, A.; Wang, Y.; Ishida, A.; Shirasaki, T.; et al. A single hepatitis B virus genome with a reporter allows the entire viral life cycle to be monitored in primary human hepatocytes. Hepatol. Commun. 2022, 6, 2441–2454. [Google Scholar] [CrossRef]
- Sugiyama, M.; Tanaka, Y.; Kato, T.; Orito, E.; Ito, K.; Acharya, S.K.; Gish, R.G.; Kramvis, A.; Shimada, T.; Izumi, N.; et al. Influence of hepatitis B virus genotypes on the intra- and extracellular expression of viral DNA and antigens. Hepatology 2006, 44, 915–924. [Google Scholar] [CrossRef]
- Li, Y.Y.; Kuroki, K.; Shimakami, T.; Murai, K.; Kawaguchi, K.; Shirasaki, T.; Nio, K.; Sugimoto, S.; Nishikawa, T.; Okada, H.; et al. Hepatitis B Virus Utilizes a Retrograde Trafficking Route via the Trans-Golgi Network to Avoid Lysosomal Degradation. Cell Mol. Gastroenterol. Hepatol. 2023, 15, 533–558. [Google Scholar] [CrossRef]
- Iwamoto, M.; Watashi, K.; Tsukuda, S.; Aly, H.H.; Fukasawa, M.; Fujimoto, A.; Suzuki, R.; Aizaki, H.; Ito, T.; Koiwai, O.; et al. Evaluation and identification of hepatitis B virus entry inhibitors using HepG2 cells overexpressing a membrane transporter NTCP. Biochem. Biophys. Res. Commun. 2014, 443, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef]
- Ladner, S.K.; Otto, M.J.; Barker, C.S.; Zaifert, K.; Wang, G.H.; Guo, J.T.; Seeger, C.; King, R.W. Inducible expression of human hepatitis B virus (HBV) in stably transfected hepatoblastoma cells: A novel system for screening potential inhibitors of HBV replication. Antimicrob. Agents Chemother. 1997, 41, 1715–1720. [Google Scholar] [CrossRef]
- Ishida, Y.; Yamasaki, C.; Yanagi, A.; Yoshizane, Y.; Fujikawa, K.; Watashi, K.; Abe, H.; Wakita, T.; Hayes, C.N.; Chayama, K.; et al. Novel robust in vitro hepatitis B virus infection model using fresh human hepatocytes isolated from humanized mice. Am. J. Pathol. 2015, 185, 1275–1285. [Google Scholar] [CrossRef]
- Yamasaki, C.; Kataoka, M.; Kato, Y.; Kakuni, M.; Usuda, S.; Ohzone, Y.; Matsuda, S.; Adachi, Y.; Ninomiya, S.; Itamoto, T.; et al. In vitro evaluation of cytochrome P450 and glucuronidation activities in hepatocytes isolated from liver-humanized mice. Drug Metab. Pharmacokinet. 2010, 25, 539–550. [Google Scholar] [CrossRef]
- Iwamoto, M.; Saso, W.; Sugiyama, R.; Ishii, K.; Ohki, M.; Nagamori, S.; Suzuki, R.; Aizaki, H.; Ryo, A.; Yun, J.H.; et al. Epidermal growth factor receptor is a host-entry cofactor triggering hepatitis B virus internalization. Proc. Natl. Acad. Sci. USA 2019, 116, 8487–8492. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Ko, C.; Henning, C.; Lucko, A.; Harris, J.M.; Chen, F.; Zhuang, X.; Wettengel, J.M.; Roessler, S.; Protzer, U.; et al. Synchronised infection identifies early rate-limiting steps in the hepatitis B virus life cycle. Cell Microbiol. 2020, 22, e13250. [Google Scholar] [CrossRef] [PubMed]
- Murai, K.; Shirasaki, T.; Honda, M.; Shimizu, R.; Shimakami, T.; Nakasho, S.; Shirasaki, N.; Okada, H.; Sakai, Y.; Yamashita, T.; et al. Peretinoin, an Acyclic Retinoid, Inhibits Hepatitis B Virus Replication by Suppressing Sphingosine Metabolic Pathway In Vitro. Int. J. Mol. Sci. 2018, 19, 108. [Google Scholar] [CrossRef] [PubMed]
- Ratheesh, M.; Sindhu, G.; Helen, A. Anti-inflammatory effect of quinoline alkaloid skimmianine isolated from Ruta graveolens L. Inflamm. Res. 2013, 62, 367–376. [Google Scholar] [CrossRef]
- Kiplimo, J.J.; Islam, M.S.; Koorbanally, N.A. A novel flavonoid and furoquinoline alkaloids from Vepris glomerata and their antioxidant activity. Nat. Prod. Commun. 2011, 6, 1847–1850. [Google Scholar] [CrossRef]
- Hanawa, F.; Fokialakis, N.; Skaltsounis, A.L. Photo-activated DNA binding and antimicrobial activities of furoquinoline and pyranoquinolone alkaloids from rutaceae. Planta Med. 2004, 70, 531–535. [Google Scholar] [CrossRef]
- Huang, A.; Xu, H.; Zhan, R.; Chen, W.; Liu, J.; Chi, Y.; Chen, D.; Ji, X.; Luo, C. Metabolic Profile of Skimmianine in Rats Determined by Ultra-Performance Liquid Chromatography Coupled with Quadrupole Time-of-Flight Tandem Mass Spectrometry. Molecules 2017, 22, 489. [Google Scholar] [CrossRef]
- Yang, Z.D.; Zhang, D.B.; Ren, J.; Yang, M.J. Skimmianine, a furoquinoline alkaloid from Zanthoxylum nitidum as a potential acetylcholinesterase inhibitor. Med. Chem. Res. 2012, 21, 722–725. [Google Scholar] [CrossRef]
- Ou, G.; He, L.; Wang, L.; Song, J.; Lai, X.; Tian, X.; Wang, L.; Zhang, K.; Zhang, X.; Deng, J.; et al. The Genotype (A to H) Dependent N-terminal Sequence of HBV Large Surface Protein Affects Viral Replication, Secretion and Infectivity. Front. Microbiol. 2021, 12, 687785. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Yan, J.; Xia, C.; Lin, Y.; Jiang, X.; Liu, H.; Ren, H.; Yan, J.; Lin, J.; He, X. Visualizing hepatitis B virus with biarsenical labelling in living cells. Liver Int. 2014, 34, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.A.; Kim, S.; Park, M.; Park, H.J.; Kim, J.H.; Park, S.; Hwang, J.R.; Kim, Y.C.; Jun Kim, Y.; Cho, Y.; et al. Ciclopirox inhibits Hepatitis B Virus secretion by blocking capsid assembly. Nat. Commun. 2019, 10, 2184. [Google Scholar] [CrossRef]
- Yang, G.; Chen, D. Alkaloids from the roots of Zanthoxylum nitidum and their antiviral and antifungal effects. Chem. Biodivers. 2008, 5, 1718–1722. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoshita, M.; Funaki, M.; Shimakami, T.; Kakuya, M.; Murai, K.; Sugimoto, S.; Kawase, S.; Matsumori, K.; Kawane, T.; Nishikawa, T.; et al. High-Throughput Screening of Antiviral Compounds Using a Recombinant Hepatitis B Virus and Identification of a Possible Infection Inhibitor, Skimmianine. Viruses 2024, 16, 1346. https://doi.org/10.3390/v16081346
Yoshita M, Funaki M, Shimakami T, Kakuya M, Murai K, Sugimoto S, Kawase S, Matsumori K, Kawane T, Nishikawa T, et al. High-Throughput Screening of Antiviral Compounds Using a Recombinant Hepatitis B Virus and Identification of a Possible Infection Inhibitor, Skimmianine. Viruses. 2024; 16(8):1346. https://doi.org/10.3390/v16081346
Chicago/Turabian StyleYoshita, Mika, Masaya Funaki, Tetsuro Shimakami, Masaki Kakuya, Kazuhisa Murai, Saiho Sugimoto, Shotaro Kawase, Koji Matsumori, Taro Kawane, Tomoki Nishikawa, and et al. 2024. "High-Throughput Screening of Antiviral Compounds Using a Recombinant Hepatitis B Virus and Identification of a Possible Infection Inhibitor, Skimmianine" Viruses 16, no. 8: 1346. https://doi.org/10.3390/v16081346
APA StyleYoshita, M., Funaki, M., Shimakami, T., Kakuya, M., Murai, K., Sugimoto, S., Kawase, S., Matsumori, K., Kawane, T., Nishikawa, T., Nakamura, A., Suzuki, R., Ishida, A., Kawasaki, N., Sato, Y., Li, Y.-Y., Sumiyadorj, A., Nio, K., Takatori, H., ... Yamashita, T. (2024). High-Throughput Screening of Antiviral Compounds Using a Recombinant Hepatitis B Virus and Identification of a Possible Infection Inhibitor, Skimmianine. Viruses, 16(8), 1346. https://doi.org/10.3390/v16081346